
Abstract
Pteropods are important components of the marine zooplankton. Not only are they crucial parts of the pelagic ecosystem but are also key contributors to the global carbon budget. Their shells sink to the ocean floor contributing to the inorganic carbon supply to the ocean floor. Being aragonitic, they easily dissolve in oceans receiving fresh water. They also dissolve as they sink beyond the Aragonite Saturation Zone, that is, within the top 500 m of the water column, releasing the trapped CO2 into the surrounding waters. Their taxonomic and quantitative distribution on the modern ocean floor can help strengthen estimates of inorganic carbon supply in shallow oceans, which are also susceptible to climate-induced dissolution. The present study, for the first time, reports 18 pteropod taxa, their abundances and spatial distribution from the surface sediments of the Ayeyarwady Delta Shelf in the North Andaman Sea, characterised by a very high influx of riverine water and sediments.
Introduction
Commonly known as ‘sea-butterflies’, pteropods are planktonic gastropods, adapted to pelagic life. During their lifetime they constitute significantly to the zooplankton population and upon death their shells sink to the ocean bed. Empty aragonitic shells of pteropods constitute a major portion of shallow marine sediments, especially in the tropical–subtropical regions (Herman, 1968). The fact that their exoskeleton is made of aragonite, a less stable form of calcium carbonate, led to the belief that they have restricted spatial-temporal distribution/preservation. Berner (1977) prepared a global map for the distribution of pteropod ooze, based on sediment distribution data generated before (Chen, 1964; Herman, 1971; Murray & Chumley, 1924). In the Indian Ocean, he identified only four regions showing accumulation of pteropod ooze on the sea floor viz. (1) northern Arabian Sea near the mouth of the Persian Gulf, (2) southwest of the southern tip of India centred on 75°E, (3) along the eastern coast of Tanzania and (4) northeast of the northern tip of Madagascar (Seychelles plateau). However, Panchang et al. (2007) for the first time reported 10 species of pteropod species with varying states of preservation (very good to very poor) from two sediment cores collected from 37 to 50 m water depths at the mouths of the Ayeyarwady, on the Burmese Delta Shelf, in the North Andaman Sea. From the low abundances, the occasional occurrence of Creseis spp. and the absence of Limacina spp., they had concluded that due to freshwater discharged from the Ayeyarwady, the region was not very suitable for the preservation of pteropods. The present study is a comprehensive report of the qualitative and quantitative distribution of pteropod taxa from the surface sediments of the Ayeyarwady delta shelf, covering a sampling area of about 2 lakh sq km.
Van Straaten (1972) demonstrated that speciation in pteropods was a function of depth and especially at depths shallower than 400 m, considerable differences in species composition in the sediment occur, compared with deeper samples. The abundance of pteropods within different water masses is controlled by specific physico-chemical properties and by the adaptive potential of the individual species. In different water masses, different forms (or even subspecies) (Van der Spoel, 1971) of a certain species may develop. In regions where these water masses mix, populations may interbreed. The recognition of forms, clines and interbreeding represents one of the important developments in recent biogeographical studies, as they have proved to be valuable ecological indicators (Ambokar et al., 2022, 2023; Furnestin, 1979; Pafort-Van Iersel, 1982; Van der Spoel, 1967, 1976). These developments in biogeography make pteropods an exceptionally interesting group in paleoenvironmental reconstructions (Almogi-Labin et al., 1986, 1991; Biekart, 1989; Chen, 1969; Schalk, 1990). The present study, thus should be able to provide insights into pteropod distribution and deposition from a region not studied before and help understand their less-known ecological niches and aragonite distribution over one of the lowest salinity regimes in the tropics.
Study Area
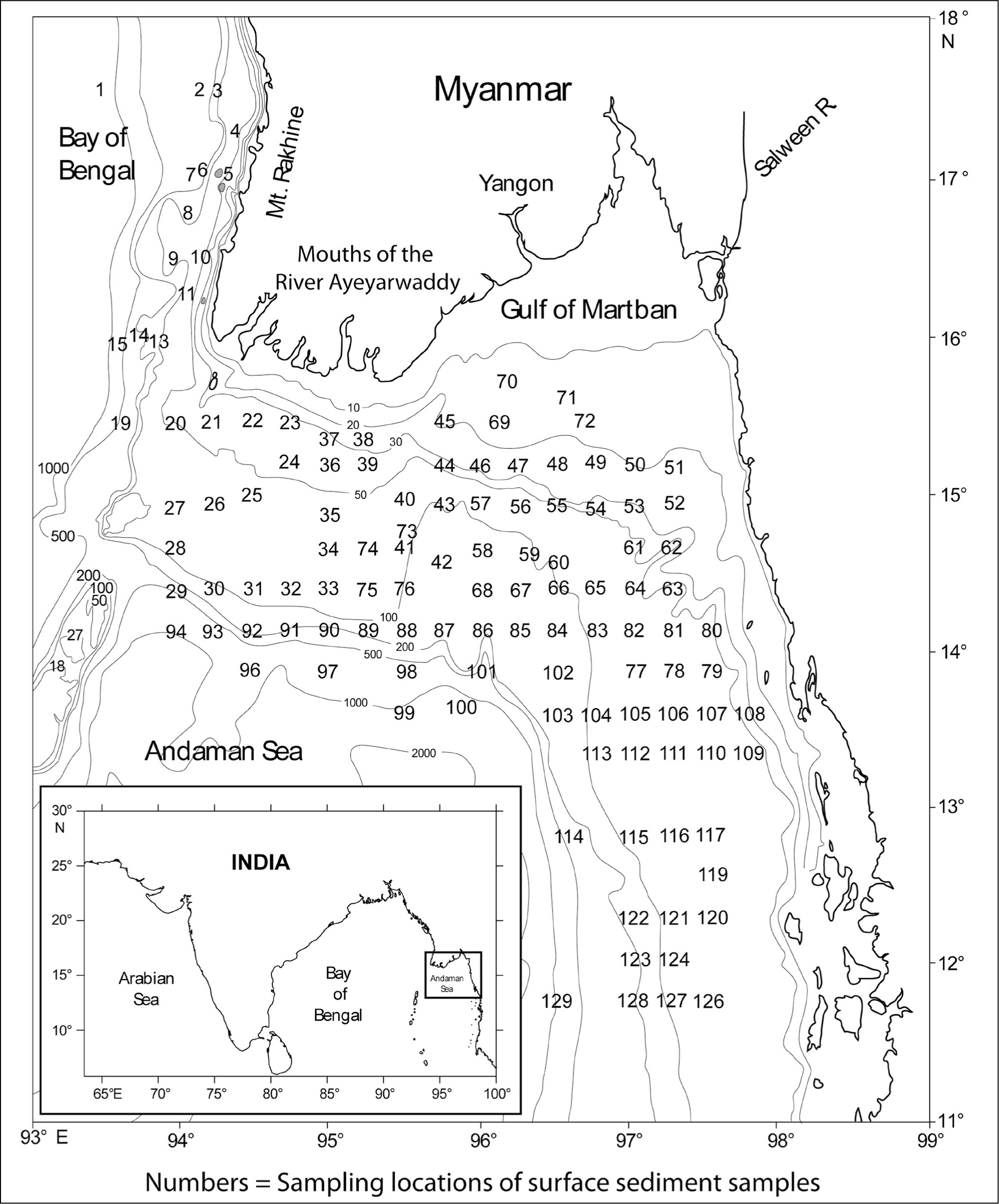
The shelf region off the Ayeyarwady Delta has a complex geological setting in the Andaman Basin (Curray et al., 1979). The shelf width is about 170 km off the Ayeyarwady River mouths and increases to more than 250 km in the centre of the Gulf of Martaban. Bathymetric data acquired during the present study shows that the shelf break is at 110 m isobath (Figure 1). Beyond the shelf break the depth increases rapidly to approximately 2,000 m, except in the bathymetric lows.
Based on grain size variations three distinct areas of sediment texture have been delineated on the Ayeyarwady shelf (i) near shore muds, (ii) outer shelf relict sands, and (iii) mixed sediments in the Martaban (Rao et al., 2005). The outer shelf has been known to be a zone of non-deposition and starved of modern fine-grained sediments. The relict sands cover an area of about 50,000 km2 suggesting probable deposition during the Holocene transgression (Rodolfo, 1969). Most of the sediments discharged by the Ayeyarwady are displaced eastwards by the prevailing westerly currents into the Gulf of Martaban. The Gulf of Martaban acts as a sediment trap. The Martaban Canyon is a conduit for terrigenous sediments reaching the deep Andaman Sea. The Gulf is characterised by modern mud and mixed sediments. At the centre of the Gulf, the mud belt is as wide as 250 km and ranks amongst the largest modern mud belts of the world oceans (Rao et al., 2005).
The temperature is about 10°C at 400 m depth and drops to about 5°C at 1,000 m (Levitus & Boyer, 1994). The Ayeyarwady (formerly known as Irrawaddy) is the third largest river in the world in terms of suspended sediment discharge and together with the Salween (also known as Thanlwin) contributes annually more than 600 million tons of sediment (Robinson et al., 2007). The large freshwater discharge from the rivers Ayeyarwady and Salween causes significant salinity changes in the Andaman Sea. The sea surface salinity varies between 31.4 psu west of the shelf and 30.2 psu in the Gulf of Martaban. There is a gradual increase in the salinity gradient southwards creating a strong salinity gradient across the delta shelf under study (Panchang & Nigam, 2014).
Methodology
In April 2002, 124 grab samples (surface sediments) were collected on the continental shelf and slope off Myanmar from depths ranging between 10 and 1,130 m (Figure 1) using a modified Peterson Grab, during the 175th cruise of the ORV Sagar Kanya in the exclusive economic zone (EEZ) of Myanmar. Approximately 10 g of the top 2 cm of surface sediments in the grab sampler were collected, dried in the hot air oven, processed as per standard sediment processing techniques to remove all organic matter and clay (Panchang et al., 2008) and was washed over a 63 µm sieve to obtain the sand fraction for micropaleontological studies. Being the second most abundant group of microfossils in the sediments (after benthic foraminifera), all possible pteropod shells were picked from 1 g sand fraction, and mounted on micropaleontological slides for taxonomic and quantitative studies. The taxonomy of euthecosomatous pteropods by Bé and Gilmer (1977) was followed to identify the pteropods.
Map of the study area showing the sampling locations and bathymetry (adopted from Panchang & Nigam, 2014).
Results and Discussions
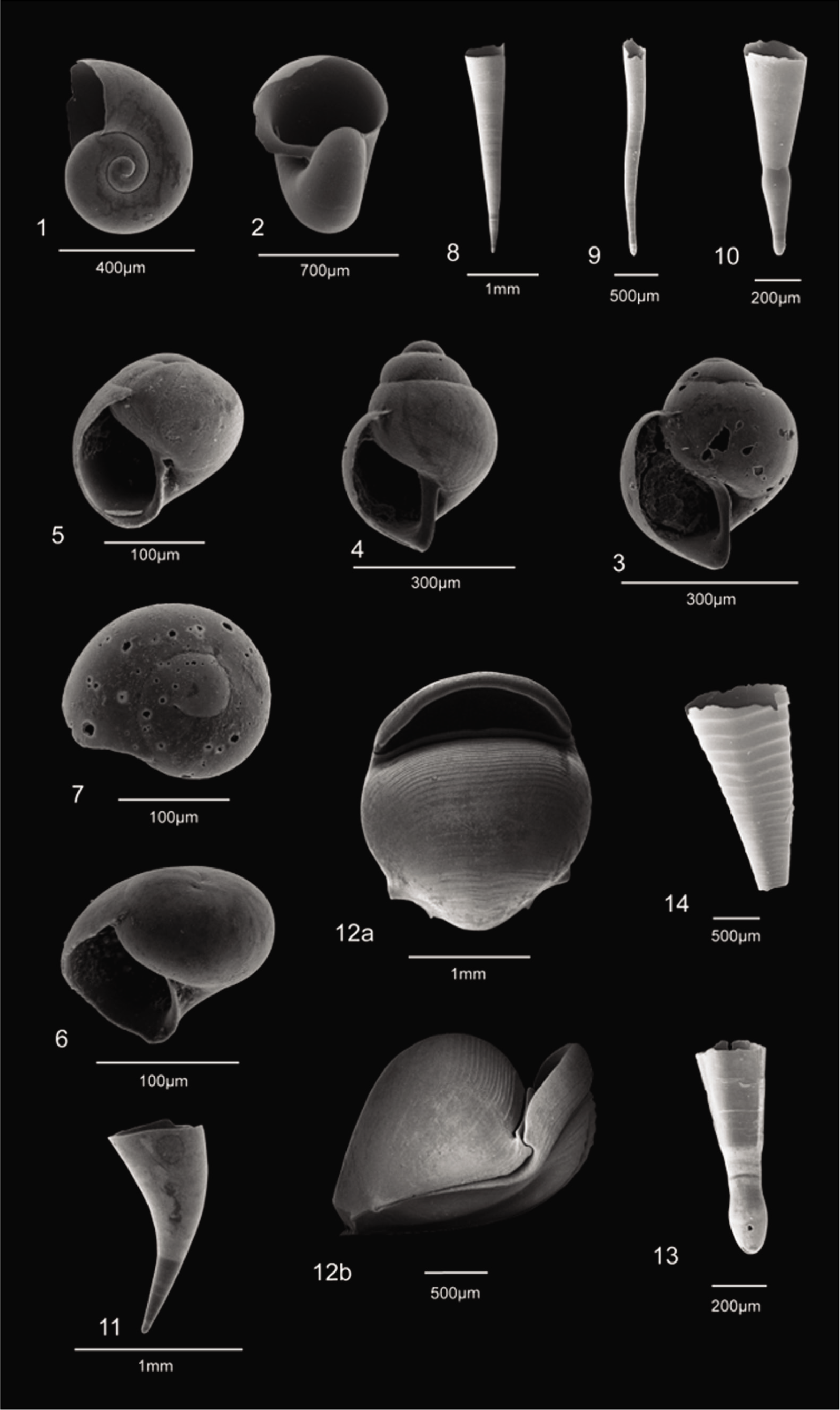
A total of 18 species were recovered from 126 surface sediments on the Ayeyarwady Delta Shelf, viz. Limacina inflata (d’Orbigny, 1836), Limacina trochiformis (d’Orbigny, 1836), Limacina bulimoides (d’Orbigny, 1836), Limacina lesueurii (d’Orbigny, 1836), Creseis conica (Eschscholtz, 1829), Creseis acicula (Rang, 1828), Creseis constricta (Chen & Bé, 1964), Creseis virgula (Rang, 1828), Diacria quadridentata (de Blainville, 1821), Diacria trispinosa (de Blainville, 1821), Hyalocylis striata (Rang, 1828), Cavolinia longirostris (de Blainville, 1821), Cavolinia gibbosa (d’Orbigny, 1836), Clio convexa (Boas, 1886), Clio polita (Pelseneer, 1888), Peraclis reticulata (d’Orbigny, 1836), Peraclis rissoides (Tesch, 1903) and an Unknown Gymnosomata (???) (Plates 1 and 2).
1, 2 Limacina inflata (d’Orbigny); 3, 4 Limacina trochiformis (d’Orbigny); 5 Limacina bulimoides (d’Orbigny); 6, 7 Limacina lesueurii (d’Orbigny); 8 Creseis conica Eschscholtz; 9 Creseis acicula Rang; 10 Creseis constricta Chen & Bé; 11 Creseis virgula (Rang); 12(a, b) Diacria quadridentata (de Blainville); 13 Diacria trispinosa (de Blainville); 14 Hyalocylis striata (Rang).
1(a, b) Cavolinia longirostris (de Blainville); 2, 3 Cavolinia gibbosa (d’Orbigny); 4, 5 Clio convexa (Boas); 6 Clio polita Pelseneer; 7 Peraclis reticulata (d’Orbigny); 8, 9 Peraclis rissoides (Tesch); 10 Unknown Gymnosomata (???) lateral view showing a well-separated and distinct protoconch.
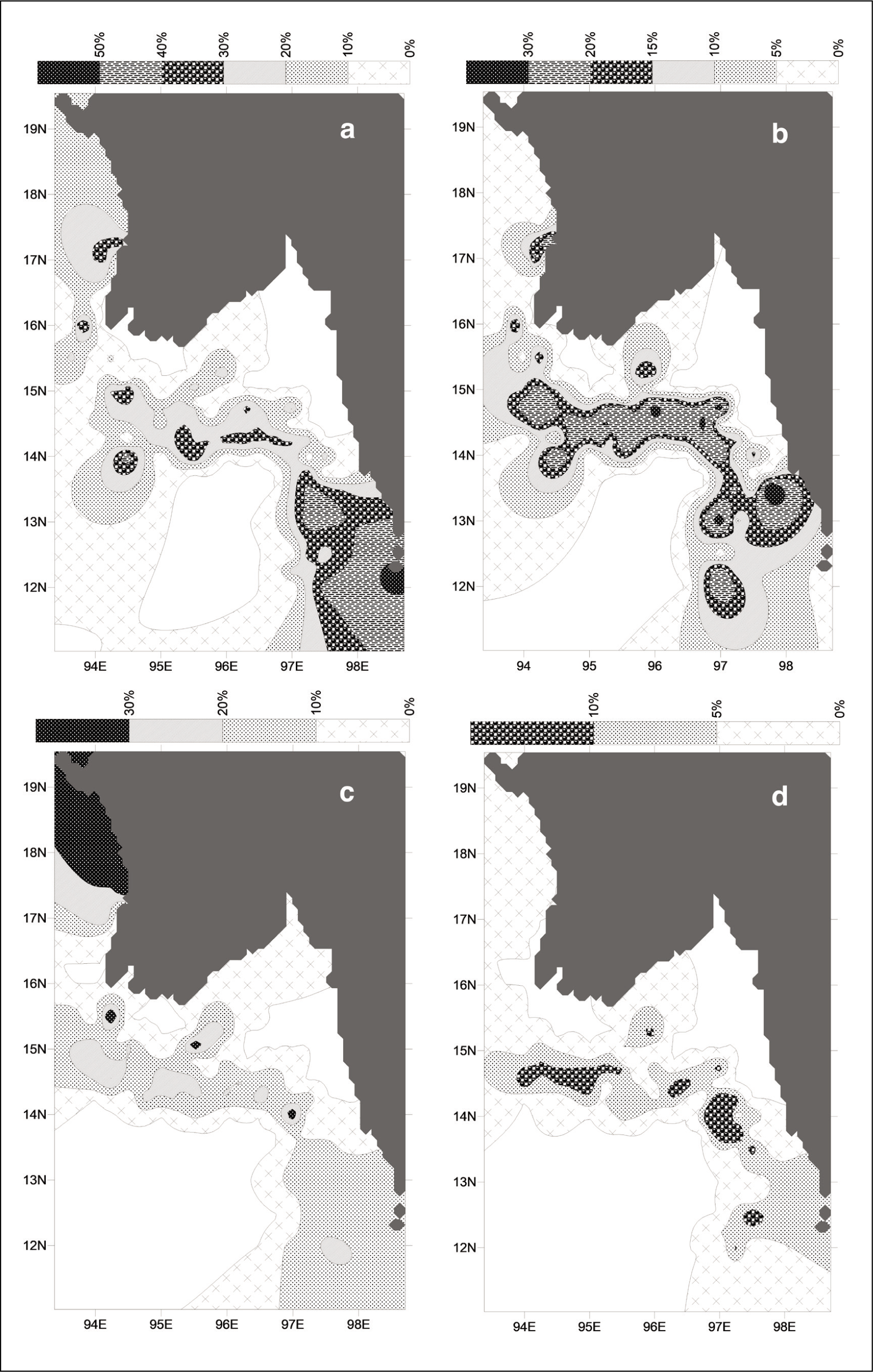
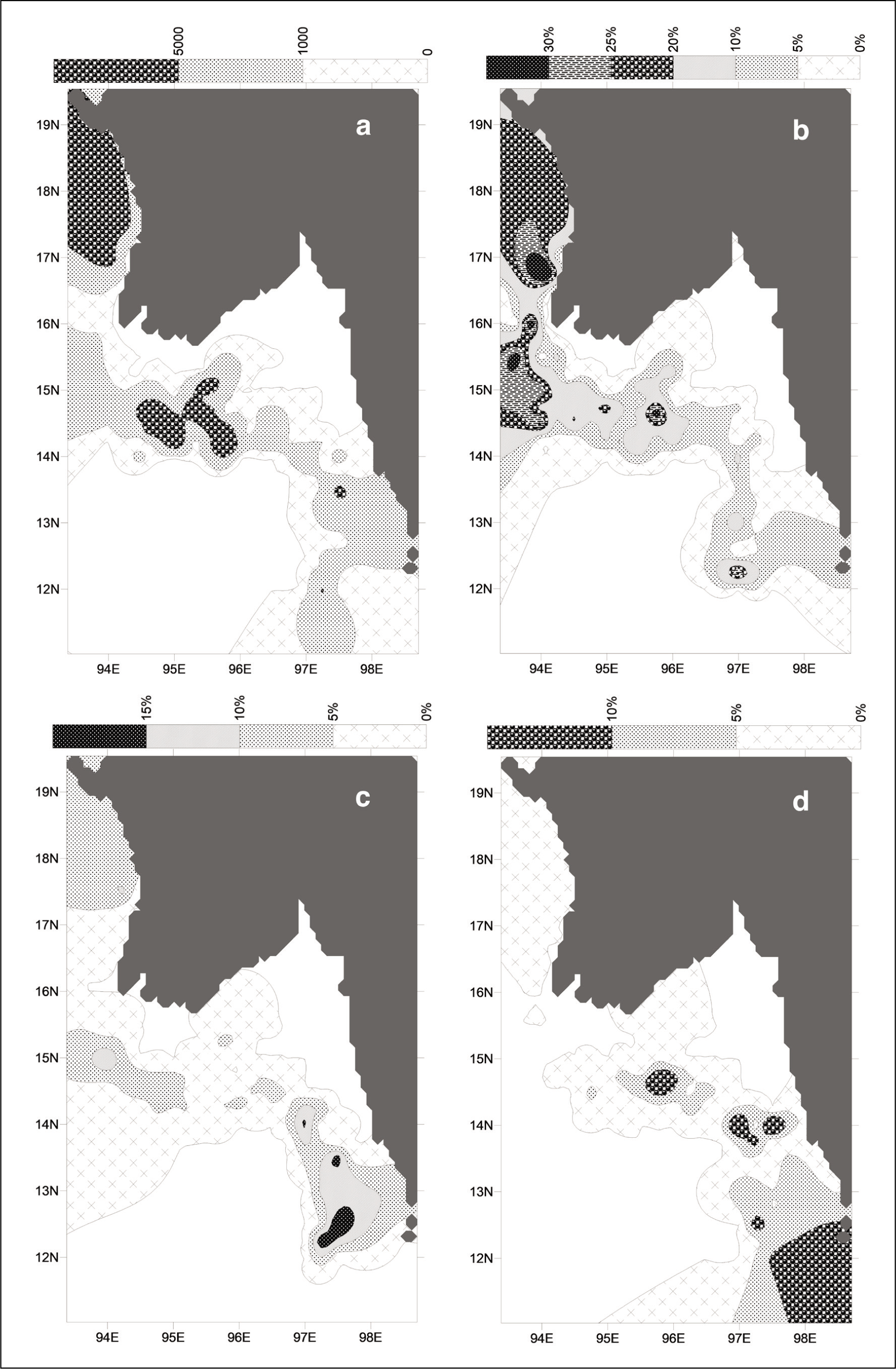
Amongst the 18 species, the 7 species that occur as major fauna (5% or more at a minimum of 5 stations) are L. inflata, L. bulimoides, Limacina leseurii, C. conica, C. acicula, C. constricta and Peraclis Sp. Their spatial distributions are illustrated in contour maps (Figures 2 and 3).
Spatial distribution of (a) total pteropods abundance; (b) L. inflata; (c) L. bulimoides and (d) L. leseurii.
The total abundance of pteropods ranges between a maximum of 22,923/g sand at Station 32 and a minimum of 121/g sand at Station 128. However, there are only 5 stations that show abundances greater than 10,000 (Figure 2a). From the maximum abundances occurring off the Rakhine coast and in the outer shelf regions off the Ayeyarwady delta, it is evident that the pteropods do not occur in regions close to the river mouths and the Gulf of Martaban.
Maximum L. inflata are seen along the Rakhine coast known to be an upwelling region (Li et al., 2020) and its complete absence in regions around the mouth of the Ayeyarwady is suggestive of its aversion to fresh water and turbidity (Figure 2b). Similarly, L. bulimoides, also known as upwelling indicators, shows abundances above 10% at very few locations far down southeast of the study area (Figure 2c) also reported to be chlorophyll rich zones (Li et al., 2020)
L. leseurii shows maximum abundances in the outer shelf away from the Gulf of Martaban, closer to the Mergui shelf and southeast of the study area (Figure 2d), indicating an aversion for low salinity and turbid waters. C. conica seems to prefer deeper depths with maximum abundance in the outer shelf and over the entire breadth of the Mergui shelf, southeast of the study area (Figure 3a). The maximum abundances of both, on the Mergui shelf away from the Martaban, suggests that they seem to avoid low saline regions but prefer high nutrients brought down by longshore currents from the Sittang, Salween and other rivers joining the Mergui coast (Reid, 2013).
C. acicula abundances suggest a preference for shallower depths but occur across the entire outer shelf; maximum abundances are concentrated along the shelf break (Figure 3b). Its low occurrences at some stations surrounding the mouths of the Ayeyarwady indicate its sensitivity to low salinities, though it seems to be more tolerant than C. conica. Panchang and Nigam (2014) report excellent preservation of C. acicula from a core at 37 m water depth off the mouths of the Ayeyarwady, attributing it to low riverine influx approximately 350 yr BP.
Inner shelf and slope regions are characterised by lower than 5% abundances of C. constricta (Figure 3c). Its highest abundances are seen off the Rakhine coast and the outer shelf show abundances between 10% and 20%. The presence of this species in regions close to the mouths of the Ayeyarwady indicates its tolerance to lower salinities.
Figure 3d shows the combined abundances of both P. reticulata and P. rissoides. Their maximum abundances are seen on the outer shelf. Their complete absence from the mouths of the Ayeyarwady indicates intolerance to low salinities. Ambokar et al. (2022) have reported that the abundances of both these species peak during cooler periods, indicating a preference for cooler temperatures and higher salinities.
Spatial distribution of (a) C. conica; (b) C. acicula; (c) C. constricta and (d) Peraclis sp.
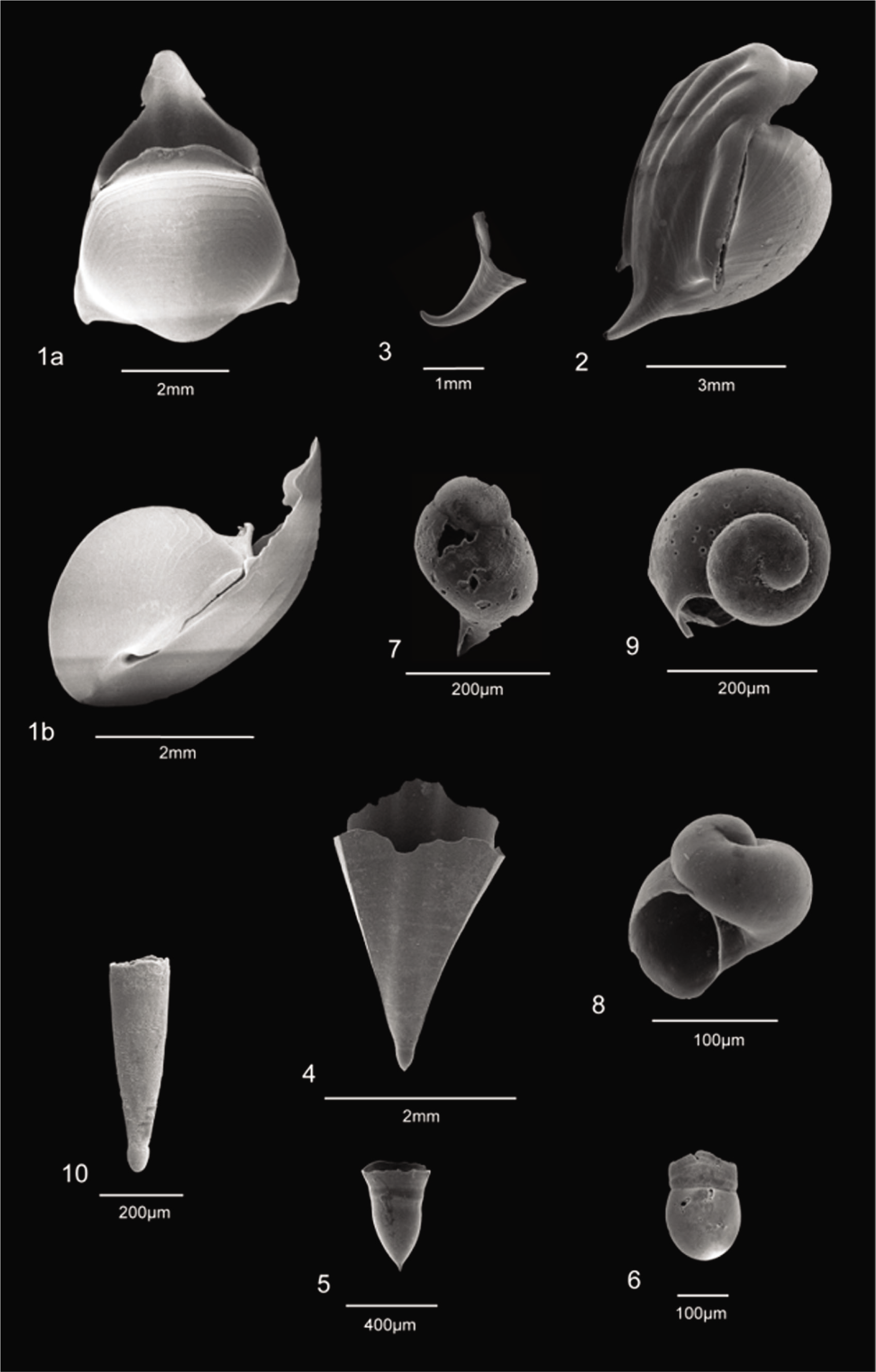
L. trochiformis (max. 14%), D. quadridentata (max. 42%), D. trispinosa (max. ~4%), C. longirostris (max. 20%) and C. convexa (max. 20%) are the other species whose occurrences remain nil to below 5% across the entire shelf. High abundances as stated in parenthesis occur only at a few deeper stations in the outer shelf. C. virgula occurs only at Station 4 with an abundance of ~4% off the Rakhine coast. It is very rare to find intact specimens of H. striata, though its typically concentrically banded shell fragments are found intermittently at various locations. Its maximum abundance is 1.11% at Station 91. The other rare species are C. gibbosa, C. polita and P. reticulata.
Creseis is the only genus that shows very high abundances and is found at all depths covering the middle to outer shelf. The Limacina species are restricted to the middle shelf. Only L. inflata has a ubiquitous presence, though in low abundances and rare at the deeper stations.
All the pteropod species and the total pteropod number show maximum abundances along the outer shelf/shelf break. Interestingly, these trends closely coincide with high salinity (>34 p.s.u.) and low temperature (<24oC) benthic environmental zones on the delta shelf as mapped by Panchang (2008) and Panchang and Nigam (2014). Glacial periods have been known to preserve more abundant and better-quality of pteropods (Klöcker et al., 2006.). This gives an insight into the supply, deposition and preservation of aragonite on the ocean floor on the Ayeyarwady Delta Shelf. However, as physico-chemical conditions vary across the Delta Shelf, the taxa too vary, highlighting their ecological preferences.
Conclusions
The present study reports the quantitative distribution of 18 pteropod species from the entire expanse of the Ayeyarwady Delta Shelf in the Northern Andaman Sea, introducing the region on the map of aragonite distribution on the world ocean floor. It is evident from the spatial distributions that, except for C. acicula and C. constricta, all other species are intolerant towards extremely low salinities. Maximum occurrences of L. inflata, L. bulimoides and C. constricta, along the Rakhine coast, suggest that these species prefer low turbidity, high nutrients, high salinity and low temperatures. The highest abundances of all species occur in the outer shelf, identifying it as a zone of better preservation of inorganic carbon in the Northern Andaman Sea.
Footnotes
Acknowledgements
We thank Dr P.S. Rao and Dr V. Ramaswamy, former Scientists at the CSIR National Institute of Oceanography, Goa, India, for providing samples. R.P. also thanks German Academic Exchange (DAAD) for short term Fellowship during which part of this work was done.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
This work is financially supported by the Ministry of Earth Sciences, Government of India. R. P. thanks the Council of Scientific and Industrial Research for the CSIR Senior Research Fellowship. M. A. is thankful to the Ministry of Earth Sciences (MoES), Government of India for the research funding (Grant no. MoES/CCR/ Paleo – 1/ 2019) for the Senior Research Fellowship.