
Abstract
Adult T-cell leukemia/lymphoma (ATL) is an aggressive malignancy with poor survival rates. The efficacy of radiotherapy in ATL needs enhancement with radiosensitizing agents. This study investigated whether umbelliprenin (UMB) could improve the therapeutic effects of ionizing radiation (IR) in ATL cells. UMB, a naturally occurring prenylated coumarin, exhibits anticancer properties and has shown synergistic effects when combined with chemotherapeutic drugs. Despite this promising profile, there is a notable lack of research on its potential combinatorial effects with IR, particularly for ATL treatment. UMB was extracted from Ferula persica using thin layer chromatography. MT-2 cells were treated with UMB alone and in combination with various doses of IR, and cell proliferation was assessed via alamarBlue assay. Flow cytometry with annexin V and PI staining was conducted, and candidate gene expression was analyzed by qPCR. In silico analysis involved identifying pathogenic targets of ATL, constructing protein-protein interaction (PPI) networks, and evaluating CDK6 expression in MT-2 cells. Molecular docking was used to determine the interaction between UMB and CDK6. The alamarBlue assay and flow cytometry showed that pretreating ATL cells with UMB significantly (p < .0001) enhanced anti-proliferative effects of IR. The combination index indicated a synergistic effect between UMB and IR. qPCR revealed significant (p < .0001) downregulation of CD44, CDK6, c-MYC, and cFLIPL, and overexpression of cFLIPS. Computational analysis identified CDK6 as a hub gene in the PPI network, and CDK6 overexpression was confirmed in MT-2 cells. Molecular docking revealed a favorable binding interaction between UMB and the ATP-binding site of CDK6, with a JAMDA score of −2.131, surpassing the control selonsertib. The current study provides evidence that UMB enhances the anti-proliferative effects of IR on ATL cells, and highlights the significance of targeting CDK6 in combinatorial approaches.
Introduction
Recent reports on the global burden of cancer indicate that new cancer cases and related deaths in 2020 were 19.3 million and 10 million, respectively. 1 Adult T-cell leukemia/lymphoma (ATL) is a severe lymphoid neoplasm induced by human T-cell leukemia virus type 1 (HTLV-1). This blood malignancy is endemic to South-Western Japan, Iran, the Caribbean basin, Central Africa, South America and Melanesia. It is estimated that around 20 million individuals worldwide are afflicted by HTLV-1. ATL is classified into four main subtypes-acute, chronic, smoldering, and lymphomatous-each associated with different prognoses and median survival rates. Although various therapeutic modalities are available, the recurrence rate of ATL is significant, and the survival rate of this malignancy is low.2,3 Radiotherapy has been utilized against ATL, however, its effect in a combinatorial approach still needs to be explored and improved by incorporating chemical agents with radiosensitizing effects.4,5
HTLV-1-infected cells express viral Tax protein, which plays a crucial role in the development of ATL. Tax is a potent oncoprotein that drives T-cell immortalization, regulates the expression of key cell cycle mediators, and facilitates the infiltration of infected T-lymphocytes into various tissues.6–8 Tax oncoprotein induces the expression of several key proteins, including c-MYC, c-FLIP, CD44 and CDK6, which collectively promote cell survival and proliferation, inhibit cell death, and help the cells evade immunosurveillance.9–14 Increased level of c-MYC is associated with increased growth of leukemic cells, and is regarded as a critical prognostic factor in ATL patients.12,15,16 As an anti-apoptotic protein, c-FLIPL has elevated expression in hematologic malignancies, and mononuclear cells from ATL patients have shown up regulation of this protein.17,18 The overexpression of CD44 is associated with clinicopathological features and poor prognosis in ATL, acute myeloid leukemia, Hodgkin and non-Hodgkin lymphoma and acute and chronic lymphocytic leukemia.19,20 Dysregulation of BMI-1 contributes to the pathogenesis of hematologic malignancies, and is associated to unfavorable prognosis in T-cell lymphoma, acute and chronic myelocytic leukemia, multiple myeloma and acute and chronic lymphoblastic leukemia. 21 Aberrant expression of the CDK6 has been frequently reported in hematological malignancies, including T-cell lymphoma, acute and chronic lymphoblastic leukemia and non-Hodgkin lymphoma.22–24 Regarding ATL, loss of the endogenous CDK4/6 inhibitor CDKN2A is a common feature among patients, suggesting that cell cycle deregulation plays a significant role in the pathobiology of ATL.25,26 In addition, genome-wide CRISPR screening identified CDK6 as a therapeutic target in ATL. 27
Umbelliprenin (UMB) is a prenylated coumarin found in edible plant species, including celery, coriander, lemon and ferula. This simple coumarin has valuable pharmaceutical activities such as antibacterial, antileishmanial, anti-oxidant and anti-inflammatory effects.28,29 In addition, UMB exerts cancer chemopreventive and anticancer activities, and has the ability to improve the effects of chemotherapeutic drugs, cisplatin, doxorubicin and arsenic trioxide.30–33 Despite the promising potential of UMB, there is a significant gap in research regarding its combinatorial effects with ionizing radiation (IR), particularly in the context of ATL treatment. Currently, radiotherapy alone has limited efficacy in treating ATL, necessitating the exploration of adjunct therapies to enhance its effectiveness. In response to this need, our study pioneers the investigation into whether UMB can function as a radiosensitizer, thereby potentially improving the therapeutic outcomes of IR in ATL cells. To that purpose, UMB was first extracted, and the proliferation of MT-2 cells was determined by alamarBlue assay upon single and combinatorial use of UMB and IR. Then, cell death was detected by flow cytometry using fluorescein isothiocyanate (FITC) annexin V and propidium iodide (PI), and the expression of CD44, CDK6, BMI-1, cFLIPL, cFLIPs and c-MYC was analyzed by quantitative polymerase chain reaction (qPCR). For in silico analysis, protein-protein interaction (PPI) networks were constructed based on the identification of pathogenic targets associated with ATL, and the expression of CDK6 was assessed in MT-2 cells using a Gene Expression Omnibus (GEO) dataset. Finally, the interaction between UMB and CDK6 was determined by molecular docking.
Methods
Preparation of UMB

aJ values are in parenthesis and reported in Hz; chemical shifts are given in ppm.
Treatment of cells
MT-2 cells were obtained from Pasteur Institute (Tehran, Iran), and grown in Roswell Park Memorial Institute-1640 (Bioldea) supplemented with 10% fetal bovine serum (Gibco) and 1% penicillin-streptomycin (Bioldea). Cells were incubated at 37°C and 5% CO2 in air. To determine effects of UMB, cells were treated with 5, 10, 20, and 40 µg/ml of this agent for 24, 48, 72, 96, 120 and 144 h. For combinatorial approach, cells were pretreated with 20 and 40 µg/ml UMB for 24, 48, 72 and 96 h, followed by IR exposure (Elekta Compact™ linear accelerator, Crawley) at three different doses (2, 4, and 6 Gy). Finally, cells were recovered for 48 and 72 h, and assessed for viability.
Proliferation assessment and interaction evaluation
After treatment with UMB, alone or in combination with IR, cell proliferation was assessed by alamarBlue (Sigma). To do so, alamarBlue (0.1 mg/mL) was added and cells were incubated at 37°C for 2 h. Subsequently, the optical density (OD) of each well was measured at 600 nm and cell viability (%) was calculated as follows: 100 ‒ (TOD ‒ UOD)/(BOD ‒ UOD) × 100, in which TOD, UOD, and BOD were OD of treated cells, untreated cells and blank control, respectively.
Based on the original method from Chou and Talalay, combination index (CI) was calculated to determine the synergistic and/or additive interaction between UMB and IR using CompuSyn 1.0 software. In addition, fold change reduction of each treatment in combinatorial use was determined by calculating the dose reduction index (DRI).
Detection of cell death
To elucidate the effects of UMB + IR on cell death, MT-2 cells were treated with 40 µg/ml UMB for 48 h, then subjected to IR (6 Gy dose) and recovered for 72 h. Afterwards, cells were collected, washed and resuspended in staining solution containing FITC-annexin V and PI (BioLegend). Finally, apoptosis was analyzed by flow cytometry (BD FACS) using FL2-H and FL1-H filters.
Gene expression analysis
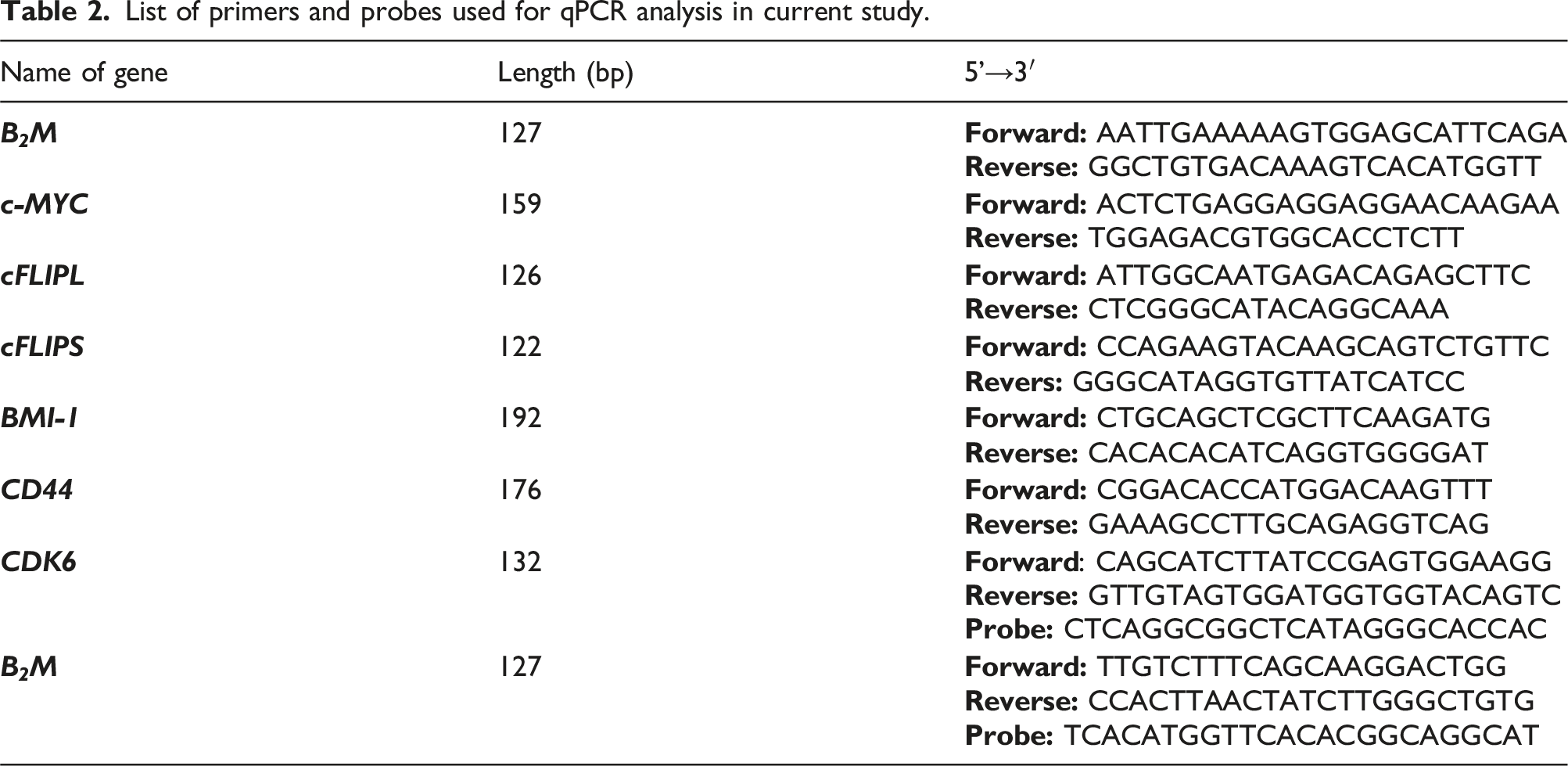
List of primers and probes used for qPCR analysis in current study.
Protein-protein interaction analysis
To define the interaction of proteins involved in ATL pathogenesis, target molecules contributing to the disease were initially screened using GeneCards (https://www.genecards.org/), a comprehensive database that provides detailed information on all annotated and predicted human genes. Subsequently, STRING database (https://string-db.org/) was utilized for interactome mapping, incorporating both direct (physical) and indirect (functional) associations. Local network clustering and Reactome pathways were analyzed, yielding a p value. The interactome data were also visualized by Cytoscape software (version 3.10.1), and key genes within the network were pinpoint via CytoHubba 0.1 plugin, which ranks genes based on their scores in the density of the maximum neighborhood component.
Data collection form Gene Expression Omnibus
Utilizing the GEO (https://www.ncbi.nlm.nih.gov/geo), a publicly available repository rich in high-throughput functional genomic data, we selected a gene expression dataset for MT-2 cells. Our search parameters included keywords ‘MT-2’ and ‘Normal lymphocyte’, and study type ‘Expression profiling by array’. Adhering to these criteria, we selected the dataset GSE6034, with the microarray data obtained from the GPL570 platform.
Molecular docking
To predict the potential interaction between UMB and the ATP-binding site of CDK6, molecular docking was performed. Crystal structure of CDK6 (ID: 4TTH) was obtained from Protein Data Bank (https://www.rcsb.org/) with resolution of 2.9 Å. Three-dimensional structure of UMB (CID: 1781413) with SDF format was retrieved from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/). In addition, selonsertib (CID: 71245288) was used as a reference point for understanding the binding characteristics of coumarin derivatives when exploring their potential as CDK6 inhibitors. 35 Docking was performed using ProteinsPlus web server (https://proteins.plus), which offers extensive molecular modeling tools on protein-ligand interactions. 36 Following pocket creation, JAMDA was utilized as a novel protein-ligand docking tool. 37 To note, protonation states and hydrogen coordinates were optimized using Protoss and docking precision was set to high. Finally, PoseEdit was employed for visualization, and 2D and 3D diagrams were generated.
Statistical analysis
Significance level was confirmed by one-way analysis of variance, followed by Tukey multiple comparison test 38 using Graphpad Prism software. The results were expressed as mean ± standard deviation (SD). p < .05, .01, 0.001 and 0.0001 were considered to be statistically significant.
Results
UMB was extracted from F. persica by thin layer chromatography. The structure of UMB was confirmed by 1H and 13C NMR, and all spectroscopic data are presented in Table 1.
To study the single and combinatorial effects of UMB, MT-2 cells were initially treated with increasing concentrations of UMB for six consecutive days. As shown in Figure 1, no significant change was observed in the proliferation of MT-2 cells upon administration of 5, 10, 20, and 40 µg/ml UMB in comparison with 0.4% DMSO during consecutive time points. Proliferation assay of MT-2 cells following treatment with increasing concentrations of UMB during 24, 48, and 72 h (a), and 96, 120 and 144 h (b). Assessment was carried out for at least three times and results are presented as mean ± SD.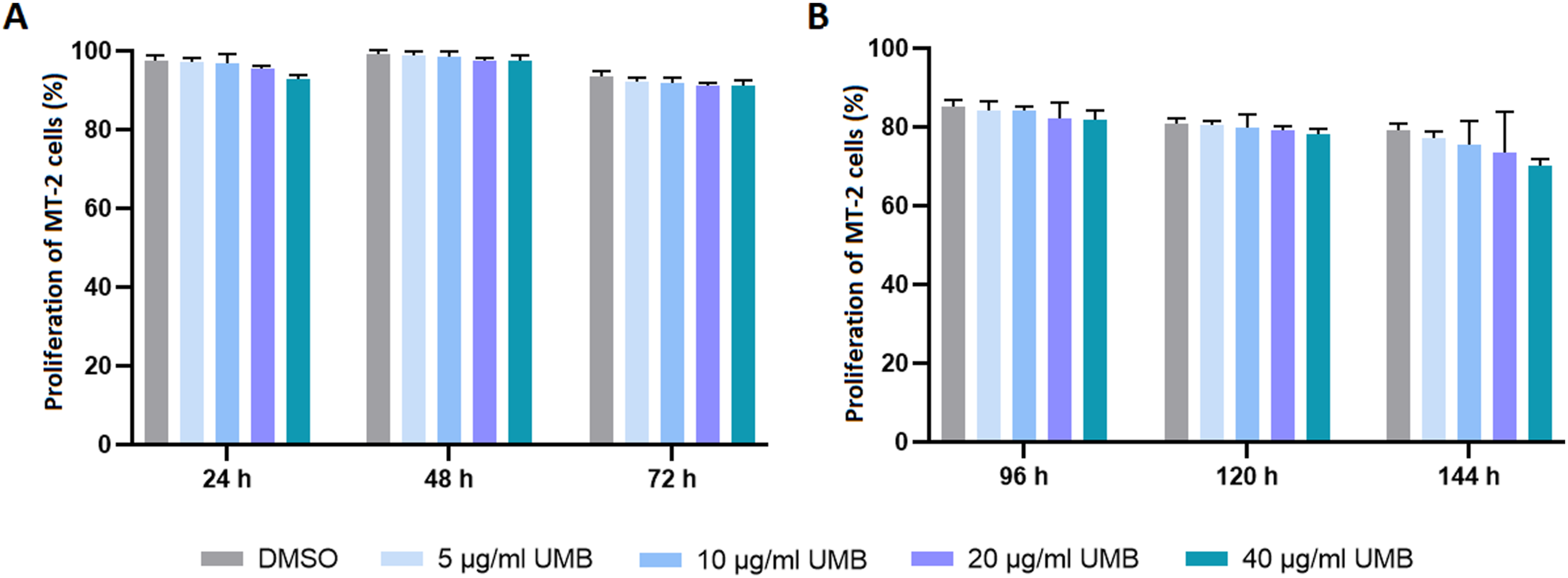
To determine the effects of UMB + IR, cells were pretreated with UMB (20 and 40 μg/mL) for 24, 48, 72 and 96 h, and then exposed to various doses of IR (2, 4, and 6 Gy). Assessment of cell proliferation after 48 and 72 h recovery revealed enhancement in the efficacy of IR compared with relevant controls (0.4% DMSO + 2, 4, or 6 Gy) almost in all treatments. As illustrated in Figure 2(a), different durations of 40 μg/mL UMB pretreatment followed by 2 Gy IR and 48 h recovery significantly (p < .05, p < .001 and p < .0001) improved effects of applied radiation. The most noticeable result was achieved upon 96 h pretreatment, as cell proliferation was significantly (p < .0001) decreased down to 64.6%. Regarding 4 Gy IR (Figure 2(b)), both 20 and 40 μg/mL UMB significantly (p < .05, p < .01, p < .001 and p < .0001) improved effects of IR upon 48 h recovery. The lowest cell proliferation was observed upon 96 h pretreatment with 40 μg/mL UMB (58.1%). As shown in Figure 2(c), all UMB pretreatments significantly (p < .05, p < .001 and p < .0001) enhanced anti-proliferative effects of 6 Gy IR. The most considerable result was achieved upon 96 h pretreatment, as cell proliferation was decreased down to 53.8%. Proliferation assay of MT-2 cells upon combinatorial treatment with UMB and IR followed by 48 h recovery. After pretreatment of cells with UMB for 24, 48, 72 and 96 h, IR was applied at 2 Gy (a), 4 Gy (b) and 6 Gy (c) and cells were recovered for 48 h. Assessment was carried out for at least three times and results are presented as mean ± SD. a p < .05, bp < .01, c p < .001 and d p < .0001 indicate significant difference with untreated cells exposed to IR alone.
Figure 3 demonstrates cell proliferation upon UMB pretreatment and IR exposure followed by 72 h recovery. Interestingly, 72 h pretreatment of MT2 cells with 40 μg/mL UMB followed by 2 Gy IR significantly (p < .0001) reduced cell proliferation down to 61.2% (Figure 3(a)). Regarding 4 Gy IR (Figure 3(b)), 40 μg/mL UMB significantly (p < .0001) improved effects of radiation upon 72 h recovery, and reduced cell proliferation down to 56.4%. As illustrated in Figure 3(c), all UMB pretreatments significantly (p < .0001) enhanced anti-proliferative effects of 6 Gy IR. The most considerable results were achieved upon 72 h pretreatment with 20 and 40 μg/mL UMB, as cell proliferation was decreased down to 49.4% and 46.4%, respectively. Proliferation assay of MT-2 cells upon combinatorial treatment with UMB and IR followed by 72 h recovery. After pretreatment of cells with UMB for 24, 48 and 72 h, IR was applied at 2 Gy (a), 4 Gy (b) and 6 Gy (c) and cells were recovered for 72 h. Assessment was carried out for at least three times and results are presented as mean ± SD. d p < .0001 indicate significant difference with untreated cells exposed to IR alone.
CI valus for combinatorial treatment of MT-2 cells upon 24, 48, 72 and 96 h pretreatment with UMB followed by X-ray exposure and 48 h recovery. *Fraction of affected cells.

CI valus for combinatorial treatment of MT-2 cells upon 24, 48 and 72 h pretreatment with UMB followed by X-ray exposure and 72 h recovery. *Fraction of affected cells.

DRI valus for combinatorial treatment of MT-2 cells upon 24, 48, 72 and 96 h pretreatment with UMB followed by X-ray exposure and 48 h recovery. The Fa* (Fraction of affected cells) valeus are obtained from Table 3.

DRI valus for combinatorial treatment of MT-2 cells upon 24, 48 and 72 h pretreatment with UMB followed by X-ray exposure and 72 h recovery. The Fa* (Fraction of affected cells) valeus are obtained from Table 4.

Based on the results of proliferation assay and interaction determination, flow cytometry analysis was performed on MT-2 cells, which were pretreated with 40 µg/ml UMB for 48 h, subjected to 6 Gy IR and recovered for 72 h (Figure 4(a)). Our findings indicted that 99.8% of untreated cells and cells treated with 40 μg/mL UMB were alive. Upon 48 h treatment with DMSO and exposure to 6 Gy IR followed by 72 h recovery, 94.4%, 2.5% and 3.1% of cells were detected as alive, early and late apoptotic, respectively. Moreover, after 48 h pretreatment with 40 μg/mL UMB, 6 Gy IR and 72 h recovery, 91.4%, 4% and 4.6% of cells were sorted as alive, early apoptotic and late apoptotic, respectively. Flow cytometry analysis with FITC-annexin V and PI staining. Alive, early apoptotic and late apoptotic cells were detected upon single and combinatorial treatments of 40 μg/mL UMB and 6 Gy IR (a). Analysis of gene expression by qPCR. After administration of UMB (40 μg/mL for 48 h) followed by IR (6 Gy), cells were recovered for 72 h and the relative expression of BMI-1, CDK6, (b), c-MYC, CD44 (c), cFLIPs and cFLIPl (d) was evaluated. Normalized values were plotted over untreated cells. a p < .05, c p < .001 and d p < .0001 indicate significant difference with DMSO.
Figure 4(b)-(d) demonstrate the expression pattern of candidate genes upon administration of 40 μg/mL UMB, alone and followed by IR. Results of qPCR indicated significant down regulation of CD44 by single use of UMB and its combination with IR (p < .001 and <0.0001, respectively). Significant (p < .0001) decrease in CDK6 expression was observed after combination of UMB and IR, in comparison with relevant control. Regarding BMI-1, significant (p < .0001) up regulation was induced by IR while combination of UMB and IR reduced BMI-1 expression. Similarly, significant (p < .0001) decrease in c-MYC expression was detected after combinatorial use of UMB and IR (p < .0001). Moreover, UMB in single and combination with IR significantly (p < .0001) induced the expression cFLIPS, although significant (p < .0001) down regulation of cFLIPL was only detected upon UMB and IR treatment. To mention, single use of 6 Gy IR up regulated the expression of CD44, CDK6, BMI-1 and cFLIPL while reduced the expression of cFLIPS.
Through screening targets from GeneCards database, a total of 349 potential molecules associated with ATL were identified, among which CDK6 (Uniprot ID: Q00534) had relevance score of 2.683, which was higher than that for CDK2 (0.410). As shown in Figure 5(a), the PPI network in STRING consisted of 92 nodes and 1512 edges with enrichment p value <1.0e-16. CDK6 was interacted with 23 nodes, including AKT1, STAT3, PTEN, TNF, JUN, MYC, CDK4, CCND1 and CCND2. The local network clustering analysis identified “drug-mediated inhibition of CDK4/CDK6 activity” as a significant enriched cluster with the false discovery rate (FDR) of 8.14e-09. Likewise, the Reactome pathway analysis identified “drug-mediated inhibition of CDK4/CDK6 activity” as a significantly enriched pathway, with the FDR of 4.90e-06. The PPI network in Cytoscape (Figure 5(b)) consisted of 93 nodes and 1516 edges. CytoHubba plugin identified top 40 hub genes in the network, including CDK6, which were connected by 640 interactions. The PPI network constructed for ATL potential targets. The STRING network consisted of 92 nodes, among which CDK6 was functionally and physically interacted with 23 targets (a). Nodes in the PPI network in Cytoscape were interacted with 1516 edges, among which 40 nodes were defined as hub genes (b). The color mapping identifies nodes from significantly high (red) to low (yellow) expression.
Given the pivotal role of CDK6 in governing cell cycle progression, we then assessed its expression in MT-2 cells. GSE6034 was selected from the GEO database, and the R software was employed to determine the expression of CDK6. As presented in Figure 6, the volcano plot revealed significant overexpression of CDK6 in MT-2 cells in comparison with normal lymphocytes. Subsequently, we conducted molecular docking to predict the spatial orientations and conformations of UMB within the ATP binding site of CDK6. As presented in Figure 7, the most favorable binding pose was characterized by a JAMDA score of −2.131, wherein two hydrogen bonds were formed with SER171 and one with ARG140. There were also four hydrophobic interactions with TYR170, PHE172, GLN173 and LEU176. Docking was also performed on selonsertib as a control CDK6 inhibitor, and the most favorable interaction had a JAMDA score of −2.092, wherein one hydrogen bond was formed with SER171, one hydrogen bond with GLN173, and three hydrophobic interactions with TYR170, PHE172 and LEU176. Volcano plot displays CDK6 expression in GSE6034 dataset (MT-2 cells). The plot uses p values and log2 (fold-change) to discriminate genes of interest. Colored dots showed genes with specific expression based on log2 FC (green dots), both p value and log2 FC (red dots), and not significant in both terms (grey dots). Molecular docking diagrams of ligands binding to CDK6. 3D and 2D images of the interactions between UMB (a) and selonsertib (b) with the ATP-binding site of CDK6. Images were generated with PoseEdit, ProteinsPlus web server.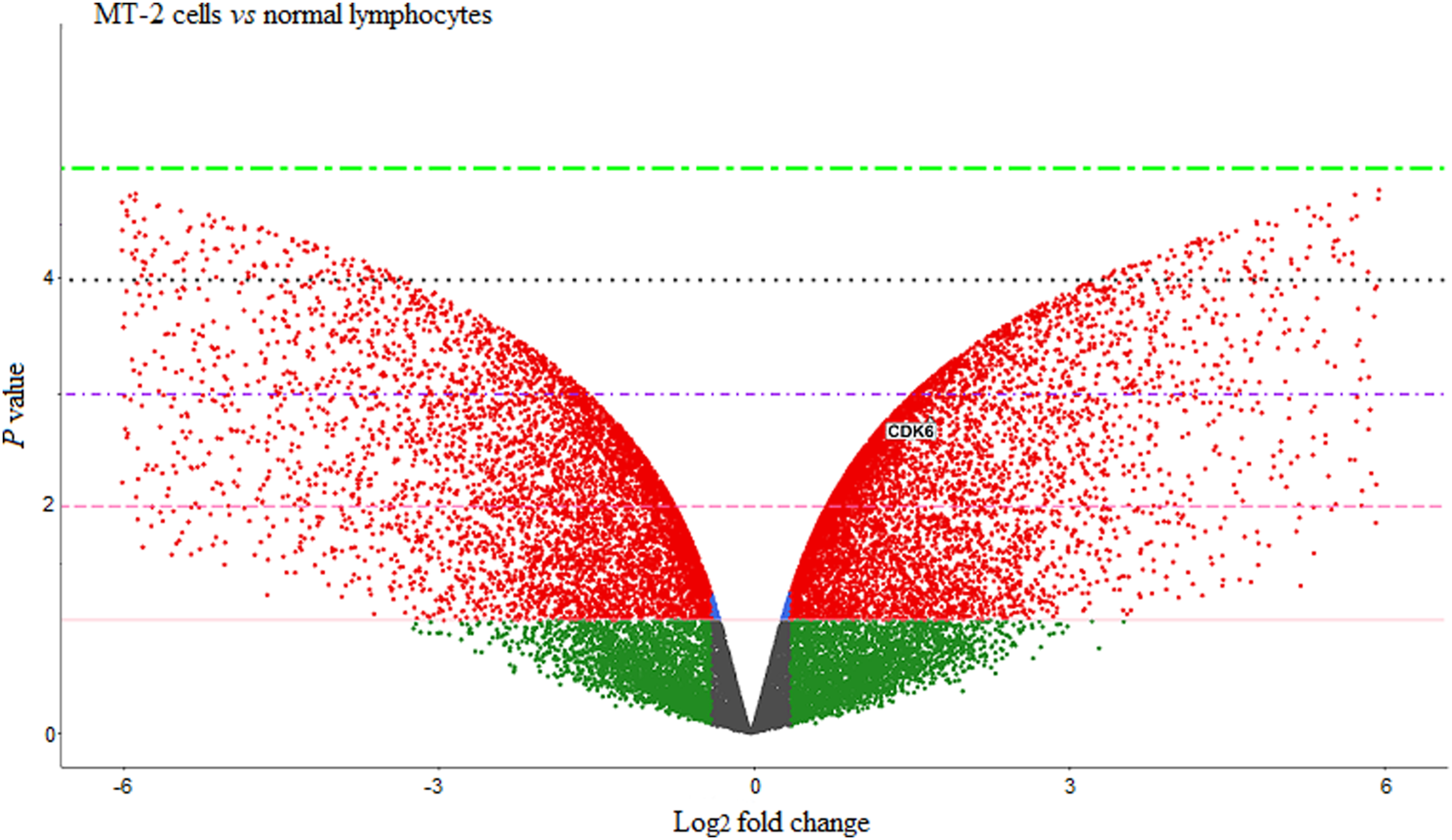

Discussion
ATL is an aggressive type of mature T-cell neoplasm, and it is estimated that approximately 20 million people worldwide are affected by this lymphoid malignancy. 3 Despite advances in therapeutic interventions, prolongation of survival rate might be difficult by intensive chemotherapy, since leukemia/lymphoma cells show chemotherapy resistance. 2 Radiotherapy is another utilized modality that its single use is not always associated with satisfactory outcomes, mainly because high dose radiations are correlated with late development of complications for ATL patients. 39 Hence, our study was concentrated on introducing a multimodal approach in which a natural agent improves effects of radiotherapy in ATL cells.
UMB is a prenylated coumarin with a wide range of pharmaceutical properties including anticancer effects. Inhibition of the cell cycle, regulation of extrinsic and intrinsic apoptotic pathways and attenuating cell migration and invasion by modulating WNT, NF-ĸB, TGFβ and FOX3 signaling pathways have been introduced as mechanisms for UMB anticancer action. 30 In addition, negative regulation of BMI-1, CD44 and NF-κB (REL-A) has been reported upon treatment of ATL cells with UMB. 33 Interestingly, previous studies reported the non-toxic nature of this coumarin on human peripheral blood mononuclear cells, and thus proved its biosafety.40,41 In the current study, we explored for the first time whether UMB could enhance the efficacy of IR on MT-2 cells. According to the obtained findings, our novel combinatorial approach led to significant enhancement in the anti-proliferative effects compared to radiation treatment alone. qPCR results indicated down regulation of CD44, CDK6, c-MYC and cFLIPL and up regulation of cFLIPS. CD44, which is linked to unfavorable clinicopathological characteristics and prognosis in ATL, 19 is involved in resistance to IR in several cancers including glioblastoma and bladder and breast carcinomas.42,43 In consistence with our findings, significant increase in CD44 expression after IR exposure has been reported previously, 44 and intriguingly, 40 µg/ml UMB, in single use and in combination with IR, induced considerable negative regulatory effects on CD44 expression. The c-MYC proto-oncogene stimulates the proliferation of leukemic cells, and is regarded as a crucial prognostic indicator in ATL patients.12,15,16 Aberrant expression of CDK6 has been detected in various hematological malignancies, and loss of the CDK4/6 inhibitor is a common feature in ATL.22–26 It has been shown that down regulation of c-MYC and CDK6 induced radiosensitizing effects on osteosarcoma and esophageal carcinoma cells.45,46 Hence, reduced expression of c-MYC and CDK6 by UMB explains, to some extent, enhanced effects of IR on ATL cells in our study. BMI-1, a polycomb ring finger oncogene that contributes to the pathogenesis of hematologic malignancies, 21 induces radioprotective effects on cancer cells. 47 It has been reported previously that IR induces BMI-1 expression,48,49 similar to our results that indicated up regulation of this gene after 6 Gy IR. Moreover, suppression of BMI-1 has been linked to increased radiosensitivity of head and neck, nasopharyngal, cervical and breast carcinoma cells.48,50–54 Therefore, down regulation of BMI-1 by UMB might be another mechanism involved in observed effects of our combinatorial approach. cFLIPL, an anti-apoptotic regulator with elevated expression in hematologic malignancies, 18 reduces radiosensitivity of lung cancer cells, and shRNA targeting cFLIP improves radioresponse of cervical cancer cells.55,56 Present results that demonstrated down regulation of cFLIPL by combination of UMB and IR indicate the significant potential of this novel approach.
Following in vitro studies, computational analyses were performed to provide a deeper understanding of UMB effects. CDK6 was among the top 40 hub genes in the PPI network, which was constructed based on molecular targets associated with ATL. CDK6 had 23 interactions in the STRING network, including those with AKT1, STAT3, JUN, MYC, CDK4, CCND1, and CCND2, all of which are associated with cell growth and proliferation. The low FDR values of “drug-mediated inhibition of CDK4/CDK6 activity” in local network clustering and Reactome pathway analyses underscore the importance of these cell cycle mediators as therapeutic targets, and highlight the relevance of developing drugs that specifically inhibit their kinase activity. In consistence with our findings, it has been shown that pharmacological inhibition of CDK6 suppressed the proliferation and survival of ATL cells. 27
Since the assessment of CKD6 expression in MT-2 cells revealed its overexpression, we performed molecular docking to predict the interaction of UMB within the ATP binding site of CDK6. The structure of CDK6 exhibits a bilobal fold characteristic of kinases, comprising an N-terminal lobe (residues 1-100) with five antiparallel β-sheets and an αC-helix, and a C-terminal lobe (residues 101-326) that is predominantly α-helical. The C-terminal lobe portion of the ATP-binding site consists of the activation loop (residues 163-189), encompassing the phosphorylation site THR177. 57 Molecular docking is a valuable tool that predicts the binding interactions between enzymes and small molecules by simulating their spatial orientations within the enzyme’s active site, thereby helping to identify potential inhibitors.58,59 In the present study, molecular docking analysis indicated a favorable interaction between UMB and the ATP-binding site of CDK6, in which two hydrogen bonds were formed with SER171 and hydrophobic interactions involved TYR170, PHE172, GLN173 and LEU176. In addition to our primary focus on UMB, we conducted docking on selonsertib as a reference compound to benchmark the binding characteristics of potential CDK6 inhibitors. Notably, results revealed that the interaction between UMB and the ATP-binding site of CDK6 was more favorable compared to selonsertib. This finding underscores the substantial potential of UMB as a potent CDK6 inhibitor, positioning it as a promising candidate for further therapeutic development.
The present study has certain limitations that warrant further investigation. To fully validate the findings from our in vitro and in silico analyses and to elucidate the precise molecular mechanisms underlying the synergistic action of UMB, additional experiments on other ATL cell lines will contribute to a more robust assessment. Furthermore, flow cytometry analysis of the MT-2 cell cycle would be beneficial in gaining a deeper comprehension of the anti-proliferative effects of UMB in combination with radiotherapy.
Conclusion
The current findings provide, for the first time, evidence that UMB enhances the anti-proliferative effects of IR on ATL cells. Furthermore, our results indicate that UMB interacts with CDK6, a cell cycle regulator that is overexpressed in ATL cells. Therefore, UMB has the potential to be developed as a powerful agent in future combinatorial therapies targeting ATL.
Footnotes
Author contributions
K. Ebrahimi and R. Bagheri performed experiments, H. Gholamhosseinian, H. Rafatpanah, and M. Iranshahi provided the research materials, M.R. Keramati supervised the project and F.B. Rassouli was contributed to the conceptualization and supervisions of the project and revision of the manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by Mashhad University of Medical Sciences (Grant NO.: 970297) and Ferdowsi University of Mashhad, Mashhad, Iran.
Ethical statement
Data availability statement
The data that support the findings of this study are available on request from the corresponding authors.