
Abstract
Cordycepin, 3’-deoxyadenosine, is an effective component isolated from the rare Chinese caterpillar fungus Cordyceps militaris. It exerts potent anti-inflammatory actions in different cell and animal models. However, its action remains unclear on the TNF-α-induced inhibition of osteogenic differentiation of adipose-derived mesenchymal stem cells (ADMSCs). In the present study, we demonstrated that cordycepin induced cell death at 20 and 40 μg/mL. Interestingly, 10 μg/mL cordycepin abrogated the cell death induced by 20 ng/mL TNF-α. Meanwhile, cordycepin exhibited a dose-dependent regulation of the osteogenesis of human ADMSCs: it promoted the differentiation at 10 μg/mL, whereas inhibited differentiation at 40 μg/mL. Furthermore, we discovered that 10 μg/mL cordycepin protected against the TNF-α (induced inhibition of osteogenic differentiation of human ADMSCs. It was also revealed that 10 μg/mL cordycepin restored Runx2 and Osx mRNA levels, which were significantly inhibited by TNF-αduring osteogenesis. At the same time, we found that 10 μg/mL cordycepin suppressed TNF-α-activated NF-κB signaling, by inhibiting IκBα phosphorylation and subsequent p65 release and translocation into the cell nucleus. Of clinical interest, the present study revealed mechanisms involved in inflammatory cytokine-inhibited osteogenesis, and it highlights the potential of cordycepin to promote the osteogenesis of human ADMSCs in cell-based therapy for inflammatory bone diseases.
Introduction
Bone homeostasis is a dynamic balance between bone resorption by osteoclasts and bone formation by osteoblasts. Inflammation disrupts the homeostasis by stimulating osteoclastogenesis and inhibiting osteoblast function.1,2 Patients with persistent inflammatory diseases have more bone loss and followed osteoporosis and bone fracture compared to the general population. 3 Different animal models of inflammation, together with clinical reports, indicate that tumor necrosis factor alpha (TNF-α) is a major etiologic factor of inflammatory bone diseases including periodontitis, spondyloarthropathies, and rheumatoid arthritis.4,5 TNF-α suppresses new bone formation via inducing osteoblast apoptosis, 6 and decreasing differentiation 7 and proliferation 8 of osteoblasts and their progenitors. During its inhibition of osteogenesis, many signaling pathways are implicated, including NF-κB, 9 MAPK, 10 Wnt/β-catenin, 11 and STAT signaling. 12
For the treatment of inflammatory diseases, cordycepin, 3’-deoxyadenosine, has attracted increasing attention over the years. Cordycepin is an effective component isolated from Cordyceps militaris which is known as a rare Chinese caterpillar fungus. 13 Cordycepin has been shown to exhibit many different pharmacological functions, such as anticancer, anti-inflammatory, and antioxidant activities. 13 Specifically, increasing studies show that it exerts potent anti-inflammatory actions in difference cell types, including macrophages,14,15 chondrocytes, 16 epithelial, and MCF-7 cells. 17 Meanwhile, such anti-inflammatory actions were also observed in different animal models, such as cerebral 18 and cardiac 19 ischemia/reperfusion injury, lipopolysaccharides (LPS)-induced acute lung injury, 17 allergic asthma. 20 Recently, cordycepin was also demonstrated to downregulate the proinflammatory cytokines in an inflammation-induced osteoporosis mouse model. 21
In the treatment of different bone diseases, mesenchymal stem cells (MSCs) have been widely used in cell-based therapy. MSCs have a potential to differentiate into different cells types, including osteoblasts, chondrocytes, muscle cells, and adipocytes. 22 MSCs can be isolated from bone marrow, synovium, periosteum, adipose tissue, and muscle.23,24 Adipose-derived MSCs (ADMSCs) are abundant in humans and they are easy to harvest. 25 Compared to the conventional bone marrow-derived MSCs, ADMSCs do not need a highly invasive procedure. Meanwhile, their proliferation rate was faster than that of bone marrow-derived MSCs.23–26 Therefore, ADMSCs have their own advantages compared to the conventional bone marrow-derived MSCs in cell-based therapy for bone diseases.
Cordycepin exerts beneficial actions on inflammatory bone diseases. However, its effect remains unclear on the osteogenic differentiation of ADMSCs in an inflammatory bone disease microenvironment during their implantation in cell-based therapy. Herein, the effect of cordycepin and possible mechanisms were investigated during the osteogenic differentiation of human ADMSCs exposed to TNF-α at different concentrations.
Materials and methods
Human ADMSC culturing
The isolation and culture of human ADMSCs were conducted according to routine protocols. In brief, after human adipose tissues were obtained by simple liposuction from abdominal subcutaneous fats from patients, who signed an informed consent form. A total of 200–300 μL of sterile saline was added to every 0.5 g of adipose tissue, and the tissue was cut into <1 mm3 size pieces. Sterile saline at a temperature of 37°C was added to the homogenized adipose tissue in a ratio of 3:1 (saline: adipose tissue) by volume, followed by the addition of stock collagenase solution to a final concentration of 0.5 U/mL. The tubes with the contents were placed on a 37°C shaker with constant agitation for 60 min. After 35–40 min of incubation, the contents were triturated with a 25 mL pipette for 2–3 min. The contents were transferred to 50 mL tubes after digestion, followed by centrifugation at 600 × g for 5 min at room temperature. The lipid layer and saline supernatant from the tube were removed. The obtained cell pellet was resuspended in 40 mL saline and centrifuged again at 600 × g for 5 min at room temperature. After being resuspended again in 5 mL saline, the cell suspension was filtered through a 100 μm filter into a 50 mL tube to which 2 mL of saline was added to rinse the remaining cells through the filter. The flow-through was pipetted to a 40 μm filter into a new 50 mL conical tube. The tubes were centrifuged for a third time at 600 × g for 5 min at room temperature. The cells were resuspended in saline. The isolated ADMSCs were further cultured in 10 mL DMEM culture medium containing 10% fetal bovine serum (FBS) for 14 days at 37°C in a humidified 5% CO2 incubator.
Induction of osteoblast differentiation
In the present study, osteogenic differentiation of human ADMSCs was induced by 10 nM dexamethasone (Dex) (Sigma, St Louis, MO, USA). To examine the osteogenesis of human ADMSCs, cells after three passages were seeded into a 96-well plate at ~80% confluent and incubated overnight. The medium was changed the next day and the cells were cultured in osteogenic medium: Dulbecco’s minimum essential medium (DMEM), 10% fetal carf serum (FBS), 10 mM glyceraldehydes 3 phosphate, 60 mM ascorbic acid, and 10 nM Dex. The osteogenic medium was changed every 2 days. Accordingly, TNF-α and/or cordycepin were also added during medium changes during treatment.
Flow cytometry analysis of human ADMSCs
Obtained human ADMSCs were analyzed with flow cytometer according to routine protocols. Briefly, the cells were seeded in a 24-well plate at 30–40% confluent following a passage, and cultured until 80–90% confluent. Then, the cells were rinsed with PBS solution containing 1% BSA, isolated by trypsinization, and resuspended in PBS solution. A total of 50 µL of cell suspension were incubated with fluorochrome-conjugated monoclonal antibody for 1 h in the dark, washed three times with PBS, and analyzed using a FACS Calibur flow cytometer (Becton Dickinson, San Jose, CA, USA). The antibodies used in the experiments were human CD29-FITC (Serotec, Oxford, UK), CD90-FITC (Serotec), CD31-FITC (Serotec), and CD45-FITC (Serotec). Mouse fluorochrome-conjugated isotype control IgG antibody (Serotec) was used as a negative control.
Alkaline phosphatase (ALP) staining and ALP activity assay
ALP, a periplasmic hydroylase, is a cell marker of osteoblast cells. ALP staining in the culture was conducted using an ALP staining kit (Sigma) according to the manufacturer’s instructions. Briefly, cells were fixed by 10% formalin for 1 h at room temperature before being stained for 5–20 min with chromogenic substrate in the kit depending on the ALP expression level.
For ALP activity assay, human ADMSCs were seeded in a 96-well plate at 30–40% confluent and cultured in osteogenic medium. Osteogenic medium was changed every 2 days. At D7, the cells were washed with TB buffer twice (20 mM Tris, pH7.5, 150 mM NaCl) before being lysed with 100 μL lysis buffer (TB buffer, plus 0.1% Triton). After centrifugation at 12,000 rpm for 20 min at 4°C, 45 μL of supernatant were incubated with 100 μL of ALP substrate p-nitrophenyl phosphatate (pNPP) liquid substrate system (Promega, Madison, WI, USA) at 37°C for 10–30 min depending on the ALP activity in the extracts. The absorbance at 405 nm was measured on a 96-well plate reader with the substrate system according to the manufacturer’s instructions. And the ALP activity was normalized against the total protein.
Alizarin red-sulfate (ARS) staining and calcium content assay
Calcium accumulation in the culture was demonstrated with Alizarin red-sulfate staining. At indicated time pointes, the medium was removed and cells were washed with PBS three times before being fixed with 70% methanol at room temperature for 30 min. After three washes with PBS, the cells were stained with Alizarin red-sulfate solution for 20 min. The cells were then rinsed with PBS to remove non-specific staining, and allowed to dry completely.
Calcium content in the human ADMSC cultures was assayed with a Calcium Assay kit (Genzyme Diagnostics, Charlottetown, PE, Canada) following the manufacturer’s protocol. Briefly, cell samples were added with 1 M acetic acid and placed on a vortex overnight at 4°C to extract calcium from mineralized matrix. In a 96-well plate, 15 μL of the cell extract were mixed with 150 μL of Calcium Assay reagent before incubation for 30 s at room temperature. The absorbance at 650 nm was measured with a 96-well plate reader and the samples were measured in triplicate. The calcium content was compared to a calcium calibration curve and normalized against the cell number.
Quantitative real-time reverse transcription PCR (RT-PCR)
Quantitative RT-PCR experiment was conducted according to routine protocols. Total RNA from cells was extracted using TRIzol reagent (Invitrogen, USA) and cDNA was synthesized using a cDNA synthesis kit (Clontech, USA). RT-PCR was performed with an ABI 7300 RT-PCR system (Applied Biosystems, USA). The relative expression level of the genes compared with the control group was calculated using the ΔΔCt method. Each value represented the average of three independent experiments.
The following primers were used for the amplification of target genes: Runx2 forward primer 5′-TCTTAGAACAAATTCTGCCCTTT-3’ and reverse primer 5′-TGCTTTGGTCTTGAAATCACA-3’; Osx forward primer 5′-CCTCCTCAGCTCACCTTCTC and reverse primer 5′-GTTGGGAGCCCAAATAGAAA-3’; and β-actin forward primer 5’-CAATGTGGCCGAGGACTTTG-3’ and reverse primer 5’-CATTCTCCTTAGAGAGAAGTGG.
Western blotting
To explore protein levels in NF-κB signaling during osteogenesis, human ADMSCs were cultured in a 6-well plate for further TNF-α and cordycepin treatment. Human ADMSC cultures were washed with PBS three times at indicated time points before being collected with fractionation buffer (10 mM pH 7.9 HEPES, 1.5 mM MgCl2, 10 mM KCl, 0.5 mM dithiothreitol, 0.5% NP40, and protease inhibitors) for 20 min on ice. The cells were fractionated by centrifugation (1000 rpm for 5 min) at 4°C to pellet nuclei. And the supernatant was collected as the cytoplasmic fraction. The two fractions were analyzed with western blot according to routine protocols. Primary antibodies used in the present study: p65 (Cell Signaling Boston, MA, USA); IкBα (Cell Signaling); p-IкBα (Cell Signaling); GAPDH (Sigma); HDAC1 (Cell Signaling). For each value, at least three independent experiments were carried out.
Statistical analysis
All data were shown as mean ± SEM. All experiments were independently repeated at least three times. Mean values were compared with the control by Student’s t-test and P <0.05 was considered significant.
Results
Phenotypic characterization of human ADMSCs
Human ADMSCs can differentiate into different cell types, including osteoblast cells for new bone formation in cell-based therapy of different bone diseases. In the present study, isolated cells were further cultured in vitro and they exhibited a typical spindle-shape of MSCs (Figure 1a). Usually, the cells grew exponentially from day 2 to day 3 and reached almost 100% confluence at day 7 (Figure 1a), which was consistent with previous reports on the growth and proliferation of ADMSCs.23,26 Meanwhile, cells were analyzed with flow cytometry. It was clearly shown that the cells used in the present study were positive in CD29 and CD90, and negative in CD31 and CD45 (Figure 1b), which are characteristic phenotypes of MSCs. Also, the ADMSCs showed normal capacity of differentiation to myogenic and adipogenic lineages (Supplementary Figure S1). It can be concluded that the cells used in the present study are human ADMSCs.

Phenotypic characterization of human ADMSCs. (a) Representative images for cultured ADMSC cells after initial seeding for 1, 3, and 7 days. Scale bar indicates 200 µm. (b) Flow cytometry analysis on ADMSCs shows the majority of cells are CD29+, CD90+, CD31–, CD45–, which are characteristic phenotypes of MSCs. The experiments were repeated three times.
Cell viability of human ADMSCs under different concentrations of TNF-α and cordycepin treatment
TNF-α is a major etiologic factor of inflammatory bone diseases. Four different concentrations of TNF-α were selected for further experiments, e.g. 1, 5, 10, and 20 μg/mL, after a series of concentrations were used for the treatment of human ADMSCs in a preliminary experiment (data not shown), consistent with previously published studies. Cell viability of human ADMSCs were measured with MTT assay, and it showed that TNF-α did not change the cell viability at 1 and 5 ng/mL compared to the control, but significantly induced cell death at 10 and 20 ng/mL from D3 (P <0.05, Figure 2a). Such effect was further confirmed by the significantly increased LDH release in the culture medium (P <0.05, Figure 2b). Thus, 20 ng/mL TNF-α was used in further experiments as it induced more cell death compared to 10 ng/mL TNF-α.

Cell viability of human ADMSCs under different concentrations of TNF-α (1, 5, 10, and 20 ng/mL) and cordycepin (1, 10, 20, and 40 μg/mL) treatment. Cell viability was measured by MTT assay and cell death was indicated by LDH release assay. Cell viability (a) and LDH release (b) of ADMSCs normalized to control under TNF-α treatment for 1, 3, and 5 days, respectively. Cell viability (c) and LDH release of ADMSCs normalized to control under cordycepin treatment for 1, 3, and 5 days, respectively. Cell viability (e) and LDH release (f) of ADMSCs normalized to control under co-treatment of 20 ng/mL TNF-α and 10 μg/mL cordycepin. The experiments were repeated triplicate. Data were presented as mean ± SEM. *P <0.05 and #P <0.01 versus control.
Meanwhile, the effect of cordycepin on the cell viability of human ADMSCs was examined at four different concentrations based on a preliminary experiment (data not shown). Both MTT assay and LDH analysis demonstrated that cordycepin did not change the cell viability significantly at 1 and 10 μg/mL, but induced cell death at 20 and 40 μg/mL (Figure 2b and c). It can be reasoned that cordycepin has little effect on the cell viability at 10 μg/mL but can exert a most protective effect against TNF-α-induced injury at the same time. Therefore, 10 μg/mL of cordycepin was used in further experiments to examine its anti-inflammatory effect on the osteogenic differentiation of human ADMSCs.
Interestingly, when the cells were co-treated with 20 ng/mL of TNF-α and 10 μg/mL of cordycepin, the cell viability did not change at days 1, 3, and 5, as evidenced by the unaffected MTT incorporation (Figure 2e) and LDH release in the culture medium (Figure 2f). A total of 20 ng/mL of TNF–α markedly inhibited the cell viability (Figure 2a and b) while 10 μg/mL of cordycepin did not change the cell viability significantly (Figure 2c and d). Therefore, it can be reasoned that 10 μg/mL of cordycepin inhibited the cell death of 20 ng/mL of TNF-α induced directly or indirectly.
Effects of different concentrations of TNF-α and cordycepin treatment on the osteogenic differentiation of human ADMSCs
Further, we examined the effects of TNF-α and cordycepin on the osteogenesis of human ADMSCs at different concentrations. Three independent experiments showed that the ALP activity in the cultures at day 7 was significantly decreased by the treatment of TNF-α at 5 ng/mL or higher from (Figure 3b, left). Such inhibitory effect of TNF-α at 5 ng/mL or higher on the osteogenesis was further confirmed by the calcium content assay at day 16 (Figure 3c, left). Interestingly, it was shown that cordycepin did not change the ALP activity at 1 and 20 μg/mL, while it promoted the activity at 10 μg/mL and inhibited at 40 μg/mL (Figure 3b, right). The same pattern was also observed in the calcium content at day 14 (Figure 3c, right). Representative images of ALP activity assay at day 7 and alizarin red staining at day 14 during osteogenesis after TNF-α or cordycepin treatment were shown in Figure 3a.
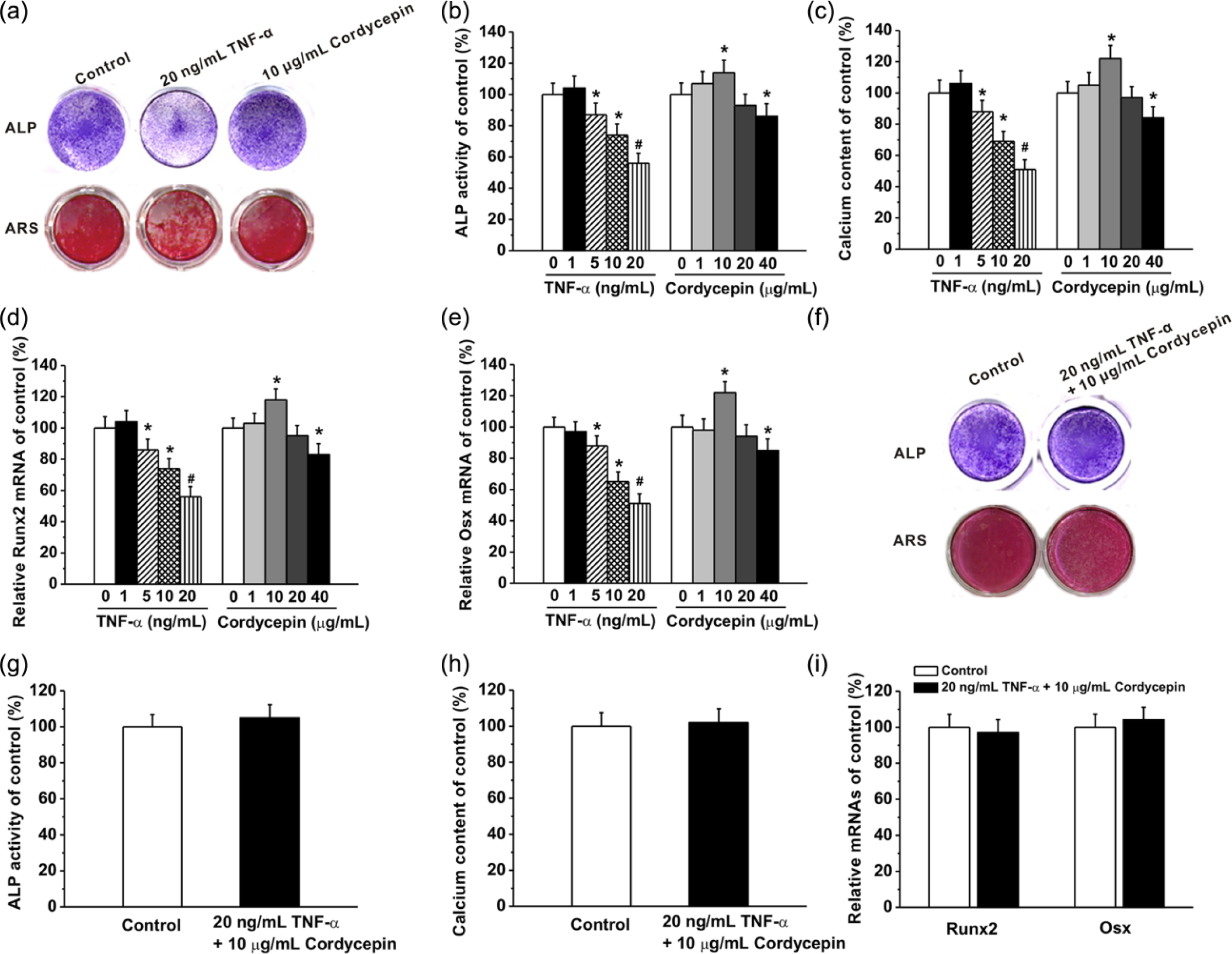
Effects of different concentrations of TNF-α (1, 5, 10, and 20 ng/mL) and cordycepin (1, 10, 20, and 40 μg/mL) treatment on the osteogenic differentiation of human ADMSCs. (a) Representative images for ALP staining in the cultures at day 7 and Alizarin red staining in the cultures at day 14. Relative ALP activity (b) and relative calcium content (c) in the different experimental groups. (d, e) The relative expression of Runx2 and Osx mRNAs in the experimental groups was quantified by qPCR analysis. (f) Representative images for ALP staining in the cultures at day 8 and Alizarin red staining in the cultures at day 16 after co-treatment of 20 ng/mL TNF-α and 10 μg/mL cordycepin. Relative ALP activity (g), relative calcium content (h), and relative expression of Runx2 and Osx mRNAs (i) in the cultures after co-treatment of 20 ng/mL TNF-α and 10 μg/mL cordycepin. The experiments were repeated three times. Data were presented as mean ± SEM. *P <0.05 and #P <0.01 versus control.
Meanwhile, two important osteogenic marker genes Runx2 and Osx were quantified by qPCR analysis at D10. Runx2 (Figure 3d) and Osx (Figure 3e) mRNA levels exhibited a same pattern as ALP activity and calcium content at different TNF-α and cordycepin concentrations. It confirmed the effects of TNF-α and cordycepin on the osteogenesis at the molecular levels.
However, when the cell were co-treated with TNF-α (20 ng/mL) and cordycepin (10 μg/mL), the ALP activity at day 8 (Figure 3g) and calcium deposition in the cultures at day 16 (Figure 3h) did not change noticeably (Figure 3f). Consistently, the Runx2 and Osx mRNA levels did not change significantly too after the co-treatment (Figure 3i). Together, cordycepin (10 μg/mL) protected against TNF-α (20 ng/mL) induced inhibition of osteogenic differentiation of human ADMSCs.
Relative protein expressions involved in the NF-κB signaling pathway during osteogenic differentiation of human ADMSCs under 20 ng/mL TNF-α treatment
NF-кB signaling is well reported to be implicated in TNF-α-inhibited osteogenesis. 27 NF-κB proteins, including p65, exist as dimers and are kept inactive by IκB proteins by forming a complex. After phosphorylated, IκB proteins are further ubiquitinylated and degraded. As a result, the NF-κB proteins are released from IкB/NF-κB complex and translocate from the cytoplasm into the nucleus, functioning as transcription factors regulating osteogenesis.
To investigate the NF-κB signaling during the osteogenesis of human ADMSCs after treated with TNF-α at 20 ng/mL, protein levels in NF-κB signaling were examined at different time points (days 3, 7, and 14). Western blotting indicated that the level of p65 protein, a subunit of NF-κB, decreased in the cytoplasm (Figure 4a) while it increased in the nucleus (Figure 4b). And such change was more pronounced with time (Figure 4a–d). Meanwhile, it was revealed the total IкBα protein level decreased in the cytoplasm and its phosphorylation level (p-IкBα) increased noticeably with time (Figure 4a and c). In short, NF-кB signaling was activated and upregulated by the treatment of TNF-α at 20 ng/mL, and such upregulation increased with time.

Relative protein expressions involved in the NF-κB signaling pathway during osteogenic differentiation of human ADMSCs under 20 ng/mL TNF-α treatment. The p65, IκBα, and p-IκBα protein bands (a) and the change of relative expressions (b) in the cytoplasm during the osteogenic differentiation at indicated time (days 0, 3, 7, and 14). The experiments were repeated three times. The p65 protein expression in the nucleus (c) and the change of relative expression (d) during the osteogenic differentiation at indicated time (days 0, 3, 7, and 14).
10 μg/mL cordycepin effectively inhibited NF-κB signaling activated by 20 ng/mL TNF-α
Further, the NF-кB signaling pathway was examined when the cells were co-treated with 10 μg/mL of cordycepin. It was revealed that 20 ng/mL TNF-α differentially regulated p65 (cytoplasmic and nuclear), IкBα, and p-IкBα (cytoplasmic) protein levels (Figure 5a and c), and three independent experiments confirmed that such changes were significant (Figure 5b and d). However, such differential changes were inhibited by 10 μg/mL cordycepin (Figure 5a and c), and such inhibitory effects of cordycepin were significant (P <0.05, Figure 5b and d).

A total of 10 μg/mL cordycepin effectively inhibited NF-κB signaling pathway activated by 20 ng/mL TNF-α. (a, b) The p65, IκBα and p-IκBα protein expressions in the cytoplasm in the cultures at day 14 during osteogenic differentiation in the experimental groups. (c, d) The p65 expression in the nucleus at day 14 during osteogenic differentiation in the experimental groups. The experiments were repeated triplicate. Data were presented as mean ± SEM. *P <0.05 versus control.
In short, 10 μg/mL of cordycepin significantly inhibited the activated NF-кB signaling pathway by 20 ng/mL of TNF-α during the osteogenesis of human ADMSCs.
Discussion
Cordyceps militaris is a precious herbal medicine used in China for many different refractory diseases including the circulatory, immune, respiratory, and glandular systems. However, its precise mechanisms are quite elusive. In the present study, we revealed that its effective component, cordycepin, has a potent anti-inflammatory effect and protected against the TNF-α-induced inhibition of osteogenic differentiation of human ADMSCs.
In the present study, we showed that cordycepin inhibited the cell proliferation of human ADMSCs at 20 μg/mL or higher and it did not change the cell proliferation significantly at 10 μg/mL or lower (Figure 2c and d). Interestingly, 10 μg/mL of cordycepin restored the cell proliferation of human ADMSCs which was significantly inhibited by the treatment of 20 ng/mL TNF-α (Figure 2e and f). One possibility is that 10 μg/mL of cordycepin interacted with TNF-α directly (20 ng/mL). Another possibility is that 10 μg/mL of cordycepin abrogated the downstream inflammatory effects of 20 ng/mL of TNF-α during cell proliferation indirectly.
Cordycepin was also found to differently regulate the osteogenic differentiation of human ADMSCs at different concentrations, i.e. it promoted the differentiation at 10 μg/mL, did not change at 20 μg/mL, and inhibited at 40 μg/mL (Figure 3). Such effect was observed in both ALP activity at the early stage and calcium deposition at the late stage during the osteogenesis (Figure 3b and c). Meanwhile, Runx2 and Osx mRNA levels at D10 confirmed this phenomenon (Figure 3d and e). The underlying mechanism is not clear for such differential regulation at different concentrations. It may be speculated that cordycepin has a beneficial effect on the osteogenic differentiation of human ADMSCs itself. However, it inhibits the cell proliferation of human ADMSCs, as shown in Figure 2c and d, and such inhibition was more pronounced at a higher concentration, e.g. 20 and 40 μg/mL. Its inhibitory effect on the cell proliferation is weak at a lower concentration, and therefore the osteogenesis of human ADMSCs was promoted, e.g. at 10 μg/mL. At 20 μg/mL, its beneficial effect on osteogenesis and the inhibitory action against cell proliferation counteracted each other, therefore the osteogenesis did not alter significantly at 20 μg/mL. However, at 40 μg/mL, the inhibitory effect of cordycepin on the cell proliferation overwhelmed its beneficial effect on osteogenesis, thus it was observed a suppressed osteogenic differentiation at this concentration.
In the present study, when the human ADMSCs were treated with 20 ng/mL of TNF-α, the ALP activity at day 7 was 57% of that in the control without treatment (P <0.05, Figure 3b). Meanwhile, when treated with 10 μg/mL cordycepin, the ALP activity at day 7 was 114% of that in the control (P <0.05, Figure 3b). Interestingly, when cells were co-treated with 20 ng/mL of TNF-α and 10 μg/mL of cordycepin, the ALP activity was not significantly changed (Figure 3g). It was evident that the beneficial effect of 10 μg/mL of cordycepin could not totally compensate the inhibitory effect of 20 ng/mL of TNF-α. Similar phenomenon was also observed in calcium deposition (Figure 3h) and Runx2 and Osx mRNAs (Figure 3i). Therefore, it can be reasoned that direct or indirect interaction between cordycepin (10 μg/mL) and TNF-α (20 ng/mL) may be occurring in addition to the beneficial effect of cordycepin on the osteogenesis itself. Further mechanistic study showed that 10 μg/mL cordycepin significantly inhibited the 20 ng/mL TNF-α activated NF-κB signaling, a critical signaling pathway involved in the downregulation of osteogenesis by TNF-α.
Generally, TNF-α may suppress the osteogenesis of human ADMSCs in different ways, e.g. inhibiting proliferation and differentiation of MSCs into osteoblasts, inducing apoptosis of osteoblast linage cells, and suppressing osteoblast activity during osteogenesis. In the present study, it was clearly demonstrated that TNF-α inhibited the cell proliferation and induced cell death (Figure 2a and b). Also, it downregulated Runx2 and Osx expression. Runx2 drives the differentiation of mesenchymal cells into osteoblasts, and it is the master regulator of osteoblast differentiation since it is required. 28 Osx is a critical regulator of the early stages of osteoblast differentiation. 9 Moreover, TNF-α activated the NF-κB signaling pathway and the signaling was upregulated with time during osteogenesis (Figure 4). NF-κB signaling plays a critical role in osteogenesis and it suppresses osteoblast differentiation and activity.27,29
In the present study, cordycepin (10 μg/mL) restored the TNF-α-inhibited osteogenesis (20 ng/mL) by different mechanisms, for example through inhibition of the TNF-α induced cell death (Figure 2e and f), restoration of TNF-α-inhibited expression of Runx2 and Osx (Figure 3i). Moreover, cordycepin (10 μg/mL) downregulated the TNF-α-activated NF-κB signaling by decreasing the phosphorylation of IκBα and followed degradation, which was upregulated by 20 ng/mL of TNF-α. IκBα is an inhibitor of NF-κB signaling by forming a complex with NF-κB proteins, preventing them to translocate into the cell nucleus taking action.
In summary, we demonstrated that cordycepin inhibited cell proliferation at a higher concentration (20 μg/mL or higher), and it exhibited a dose-dependent manner in the regulation of the osteogenic differentiation of human ADMSCs. Meanwhile, it was revealed that 10 μg/mL of cordycepin effectively abrogated the inhibition of osteogenesis induced by 20 ng/mL of TNF-α. During its protection against the TNF-α-induced inhibition of osteogenic differentiation of human ADMSCs, cordycepin suppressed the TNF-α activated NF-κB signaling by direct decreasing the IκBα phosphorylation and followed degradation. The present study highlights the role of cordycepin to promote the osteogenesis of human ADMSCs used in cell-based therapy for different bone disease in addition to be used as a TNF-α inhibitor to reduce inflammation for the treatment of inflammatory bone diseases.
Conclusion
In the present study, it was shown that cordycepin promoted the osteogenic differentiation of human ADMSCs at 10 μg/mL, while inhibited the osteogenesis at 40 μg/mL, which may be due to its adverse effects against the cell proliferation of human ADMSC at 20 μg/mL or higher. Interestingly, 10 μg/mL of cordycepin restored the cell proliferation and osteogenic differentiation which were significantly inhibited by 20 mg/mL of TNF-α by different ways, including restoring the cell proliferation, Runx2 and Osx mRNA expressions, and NF-κB signaling via inhibition of IκBα phosphorylation. The present study revealed that cordycepin protected against the TNF-α-induced inhibition of osteogenic differentiation of human ADMSCs in addition to its role as an anti-TNF-α therapeutic agent for the therapy of inflammatory bone diseases.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported by the Department of Orthopaedics, Changzhou Traditional Chinese Medical Hospital, Affiliated to Nanjing University of Traditional Chinese Medicine, and we thank them for their assistance with the study.