
Abstract
Salmonellosis-induced diarrhea, is one of the commonest cause of childhood mortality in developing countries. Using of probiotics is viewed as a promising means for reducing the pathogenic loads of bacterial infection. The current study aimed to evaluate the potential antimicrobial and immunomodulatory efficacy of isolated lactobacillus strains against the enteropathogenic effect of S. Typhi. Different Lactobacillus strains were isolated from 13 dairy products. Their antimicrobial activities were tested against different bacterial strains. Six groups of CD1 mice were treated for 8 days as follows: group (1) untreated control; group (2) was challenged with single inoculation S. typhi, and groups (3) and (4) were treated with Lactobacillus plantarum (LA5) or Lactobacillus paracsi (LA7) for 7 days, respectively. Groups (5) and (6) were challenged with S. typhi, and then treated with either LA5 or LA 7 for 7 days, respectively. Isolated Lactobacillus showed antimicrobial activity against wide range of bacterial strains. Salmonellosis showed high widal titer, induced significant disturbance of TNF and IL-1β, while sever changes of the histological patterns of the intestinal villi and hepatocytes have been illustrated. LA5 or LA7 succeeded to eradicate typhoid infection, restore the values of inflammatory cytokines to typical levels of control group, and improve histological pictures of intestinal and hepatic tissues. It can be concluded that lactobacilli are promising candidate in protection and eradication against bacterial infection induced by S. Typhi due to its antimicrobial, anti-inflammatory, and immunomodulatory activities.
Introduction
Typhoid fever is the most common problem in developing countries due to poor sanitation and shortage of clean water. 1 This public health problem in humans and animals arises from infection with the gram-negative bacteria: Salmonella enterica serovar typhi (S. typhi) and S. Paratyphi A. Earlier studies reported that 217,000 deaths resulted from more than 27 million cases of typhoid fever. In addition, the annual incidence of typhoid infection has dramatically increased in Egypt; 13/100,000 people in Belbis District in the Nile Delta 2 and 61/100,000 people in Fayoum Governorate. 3 Salmonellosis induces three distinct clinical syndromes, including gastroenteritis, sepsis with or without metastatic infections, and typhoid (enteric) fever. 4 Nowadays, limited therapeutic options are available for the management of infectious diseases due to the prevalence of multidrug-resistant strains. 5 S. typhi strains showed resistance against various antimicrobial agents as β-lactams, quinolones, and aminoglycosides. Many resistance mechanisms have been described in S. typhi such as alteration of cell membrane permeability 6 and expression of multidrug efflux system. 7 Recently, there has been an increased interest to develop alternative therapeutic modalities to counteract infectious disease and overcome resistance to antibiotics. Previous literature reported that probiotics able to enhance the host immune system by multiple immune and non-immune mechanisms. 8 It exerts antimicrobial activity 9 and enhances phagocytic properties. 10 Earlier studies reported that lactobacilli play a crucial role in cytokines manufacturing 11 and IgA production. 12 Lactobacillus is widely distributed in human host, vegetable, meat, and dairy products.13,14 Mechanism of pathological changes of typhoid infection is not well understood, because of the lack of a comparable small animal model. 15 Considering the importance of lactobacillus strains isolated from Egyptian dairy products, the current study aims to evaluate the potential antimicrobial and immunomodulatory activities of isolated lactobacilli against enteropathogenic infection of S. typhi either in vitro or in vivo in CD1 mice.
Material and methods
Chemical and kits
Bismuth sulphite agar, cetrimide agar, muller hinton broth, and anutrient agar were purchased from HIMEDIA (India). Anaerobic kits (GasPAk) and API (Analytical Profile Index) 50CH were obtained from Becton, Dickinson, and BioMerieux (France), respectively. Big Dye Terminator and Ultra Sample Preparation Reagent kit were from Applied Biosystems. Febrile antigen kits of Salmonella typhi was purchased from linear chemicals (Barcelona, Spain). Interleukin-1β (IL-1β) and tumor necrosis factor-alpha (TNF) were purchased from Orgenium, Helsinki, Finland. All other chemicals and reagent were in analytical grade.
Isolation and characterization of Lactobacillus strains
The lactobacillus (LAB) strains were isolated from 13 Egyptian dairy products and processed according to El Soda et al. 16 Characterization of antimicrobial activity of crude bacteriocin was performed to assess the effect of different PH and proteolytic enzyme (Papain) on its antimicrobial activity.
Identification of lactobacillus strains
The isolated strains were identified by standard bacteriological methods. Isolates were recognized using API 50CH according to manufacturer’s instructions. In addition, molecular characterization of lactobacillus isolates was performed by PCR amplification of 16S rDNA region and sequencing. PCR reaction was carried out using PAF and 536 R primers according to the method previously described by Yeung et al. 17
In-vitro study
Antimicrobial activity of bacteriocin
Antimicrobial activity of lactobacilli were investigated against a wide range of pathogenic bacterial strains. 18
In-vivo study
Experimental animals
Five-week-old male CD-1 mice, each weighing in the range of 22–30 g, were purchased and housed from Animal House Colony, Pharmacology & Chemistry Research Centre, 6th October, Egypt. Animals were maintained on standard lab diet (protein, 160.4; fat, 36.3; fiber, 41 g/kg of metabolisable energy = 12.08 MJ) and were housed in filter-top polycarbonate cages in a room free from any source of chemical contamination, artificially illuminated (12-h dark/light cycle), and thermally controlled (25 ± 1O°C). After an acclimatization period of 1 week, the animals were divided into six groups (10 mice/group), and received human care in compliance with the guidelines of the Animal Care and Use Committee of the Pharmacology & Chemistry Research Centre.
Experimental design
Animals within different treatment groups were treated daily for 8 days as follows: Group (1), untreated control; Group (2), animals challenged with single inoculation S. typhi (200 µL aliquot of 1X 108/ P.O); Group (3), animals treated orally with Lactobacillus plantarum (LA5) (200 µL aliquot of 1X 108/ P.O) for 7 days; Group (4), animals treated orally with Lactobacillus paracsi (LA7) (200 µL aliquot of 1X 108/ P.O) for 7 days; Group (5), animals challenged with single inoculation S. typhi, then treated with LA5 for next 7 days; and Group (6), animals challenged with single inoculation S. typhi and treated with LA 7 for next 7 days.
At the end of the treatment period, all animals were fasted for 12 h and blood samples were collected from the tail vein from each animal under ketamine anesthesia according to the method of Cocchetto and Bjornsson. 19 Blood samples were left to clot and the sera were separated using cooling centrifugation at 3000 rpm for 15 min and stored at −20°C until analysis. The sera were used for performing widal test, determination of TNF, and IL-1β according to the analytical kit instructions.
After the collection of blood samples, all animals were killed by an overdose of ketamine and samples of liver and small intestine were removed for histopathological and histochemical examinations.
Widal test
Widal test has commonly used as an aiding tool in the diagnosis of typhoid fever. According to the manufacturing instructions, the titer of antibodies revealed 1:80 and more for anti-TO antibodies were positive that indicated by recent infection of typhoid fever.
Statistical analysis
All data were statistically analyzed with the General Linear Model Procedure of the Statistical Analysis System. 20 The significance of the differences among treatment groups was determined with the Waller–Duncan k-ratio. 21 All statements of significance were based on a probability of P ⩽0.05.
Results
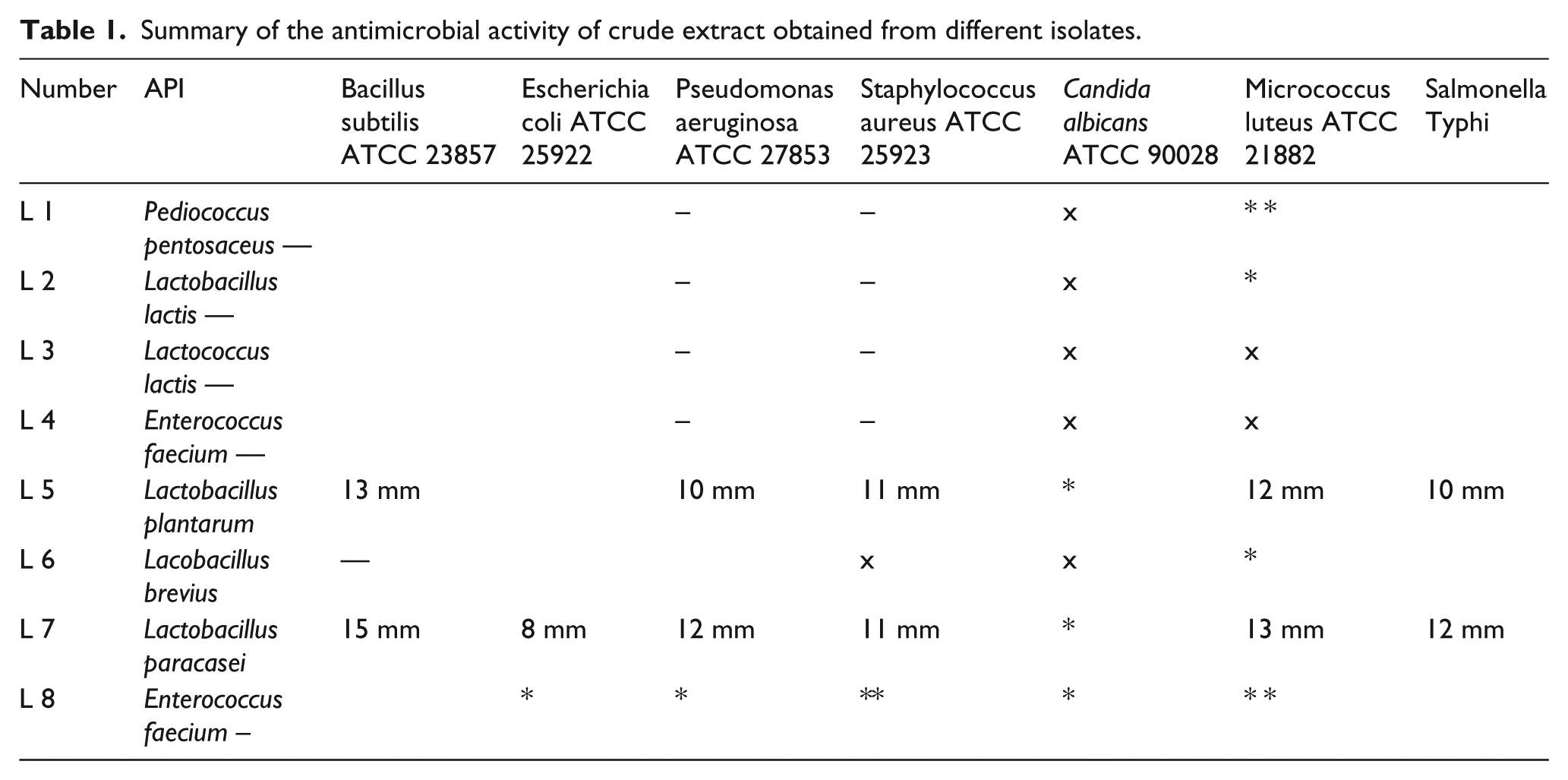
Five lactobacillus strains were isolated from 13 dairy products. Two of them were identified as lactococcus lactis, in addition to Lactobacillus brevius, Lactobacillus plantarum, and Lactobacillus paracsi. The results showed the presence of the different species of isolated lactobacilli obtained from dairy products through using of API 50CH system. PCR amplification of the 16S rRNA and sequencing revealed further confirmation of the of the isolated strain identity (Figure 1). The crude extract of Lactobacillus plantarum LA5 and Lactobacillus paracsi LA7 has a significant antimicrobial activity among different bacterial strains including S. typhi (Table 1). The highest inhibition zone value of crude bacteriocin of both LA5 and LA7 was observed at PH 2 (Figure 2). The antibacterial activity of crude extract of LA5 and LA7 isolates was completely inhibited by the proteolytic enzyme (papain).

16S rRNA amplification using PCR of Lactobacillus plantarum (LA5) and Lactobacillus paracsi (LA7).
Summary of the antimicrobial activity of crude extract obtained from different isolates.
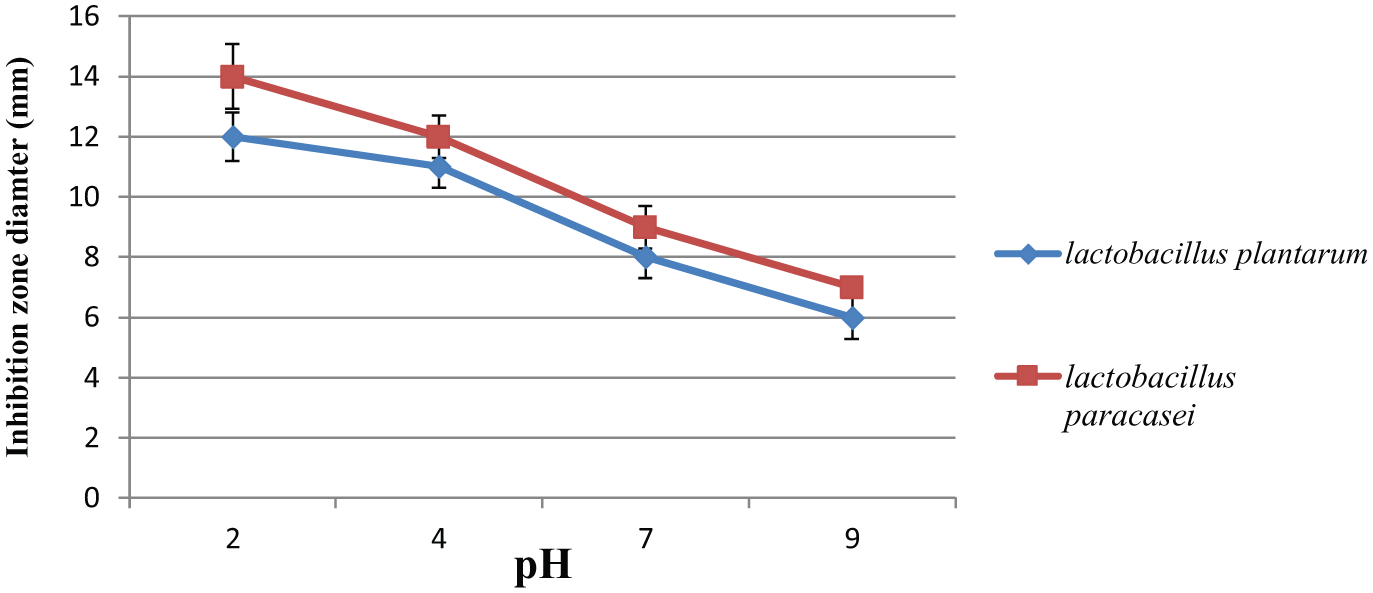
The effect of pH on the antimicrobial activity of crude extract of LA5 & LA7 ON Salmonella typhi.
The results of Widal test revealed that mice challenged with S. typhi showed highest Widal titer reached 160. The data presented in Table 2 showed that lactobacilli succeeded to reduce the risk of typhoid infection that was indicated by negative titer of Widal test less than 40 comparable to control itself. The effect of different treatments on inflammatory cytokines (Figures 3 and 4) revealed that TNF and IL-1β were significantly increased in animals infected with Salmonella typhi, while groups treated with LA5 or LA7 were comparable to the control group. However, animals infected with S. typhi and treated with LA5 or LA7 showed significant improvement in these parameters. This improvement was pronounced in the group treated with Lactobacillus paracsi (LA7).
Effect of different treatment on average of the antibody titer in infected animals with S. Typhi.


Effects of different treatment on TNF level in rats challenged with S. Typhi.

Effects of different treatment on IL-1β level in rats challenged with S. Typhi.
The histological examination of the liver section in the control mice showed normal distribution of central vein and blood sinusoids (Figure 5a). The liver section of animals challenged with S. typhi infection showed complete distortion of the lobules and replacement by variable aggregation of the inflammatory cells around the portal tracts, multifocal necrosis, and with fibrous tissue (Figure 5b).

A photomicrograph of liver section showing (a) control mice showing the central vein and hepatocytes, (b) mice challenged with S. Typhi showing multifocal necrosis edema, congestion, with fibrous tissues and inflammatory cells around the portal tracts, (c) mice treated with LA5 showing normal central vein and hepatocytes, (d) liver section of mice treated with LA7 showing normal hepatocytes architecture, (e) mice treated with LA5 and infected with S. Typhi showing marked decreased in inflammation and restore the portal tract size, (f) challenged mice and treated with LA7 showing obvious improvement in hepatocytes architecture marked decrease of fibrous tissues.
The mice treated with Lactobacillus plantarum showed normal central vein and hepatocytes (Figure 5c). Histological examination of hepatic cell in mice treated with Lactobacillus paracsi displayed normal distribution of hepatocytes (Figure 5d). Photomicrograph of liver section of mice infected with S. typhi, then treated with Lactobacillus plantarum showed marked decreased in inflammatory cells and restore the portal tract size. Few small nodules of inflammatory cells (Figure 5e). Liver section of group treated with Lactobacillus paracsi that challenged S. typhi infection showed marked improvement in hepatocytes and the central vein (Figure 5f). The microscopic examination of liver section stained with Periodic-Sheif reagent Stain (PAS) for glycogen demonstration revealed that control liver showed normal distribution of carbohydrates in hepatocytes and connective tissue (Figure 6a). The animal infected with S. typhi showed marked decrease in PAS reaction in hepatocytes due to the decrease in carbohydrates molecules (Figure 6b). A photomicrograph of liver section of mice treated with Lactobacillus plantarum showed normal distribution of carbohydrates in hepatocytes (Figure 6c). Liver section of mice treated with Lactobacillus paracsi showed significant reaction of PAS stain (Figure 6d). However, the microscopic examination of liver of infected animals with S. typhi and treated with Lactobacillus plantarum showed marked improvement in PAS reaction (Figure 6e). Lactobacillus paracsi treatment in infected mice with S. typhi showed normal distribution of carbohydrate that indicated with patchy dimension of PAS reactions (Figure 6f). The histological studies of the intestinal mucosa of the control group showed normal tall and finger-like intestine villi with crypts in between villi (Figure 7a). On the other hand, intestinal mucosa of animals infected with S. typhi revealed intestinal erosion and sloughing of surface epithelium of the tip of the villi. Numerous goblet cells, congestion, and inflammatory cells in the villi core are determined (Figure 7b). Intestinal mucosa of mice treated with Lactobacillus plantarum displayed normal tall finger-like intestine villi with pyknotic form of some enterocytes nuclei (Figure 7c). Intestinal section of mice treated with Lactobacillus paracsi showed normal tall-finger like intestine villi, but some crypts were damaged with congested blood capillaries (Figure 7d). Animals infected with S. typhi and treated with Lactobacillus plantarum showed intact surface columnar cells lining epithelium villi (Figure 7e). Lactobacillus paracsi treatment showed marked improvement of intestinal mucosa of infected animals and intact microvilli of the alimentary tract beside regular brush border (Figure 7f). The microscopic examination of intestinal section stained with Periodic-Sheif reagent Stain (PAS) for glycogen demonstration revealed that the control group showed positive reactions indicated with magenta red staining of intestinal villi in the mucous globules of goblet cells and of brush border and connective tissues (Figure 8a). On the other hand, intestinal mucosa of infected animals showed significant reduction of PAS reaction indicated with few goblet cells and interrupted staining of brush border (Figure 8b). Animals treated with Lactobacillus plantarum only showed positive reaction of PAS stain of brush border and discoloration with magenta red in mucous globules of goblet cells (Figure 8c). Intestinal section of animals treated with Lactobacillus paracsi revealed normal distribution of mucin in microvilli of goblet cells and brush border (Figure 8d). However, Lactobacillus plantarum treatment in infected mice showed positive reaction of PAS stain in many goblet cells at the villus tips with variable size (Figure 8e). Microscopic examination of intestinal mucosa of infected animals with S. typhi and treated with Lactobacillus paracsi showed prominent distribution of positive PAS reaction in goblet cells and of striated brush border (Figure 8f).

A photomicrograph of liver section stained with PAS for carbohydrates detection shows: (a) control mice showing normal distribution of carbohydrates in hepatocytes and in connective tissues, (b) mice challenged with S. Typhi showing marked decrease in PAS reaction in hepatocytes, (c) mice treated with LA5 showing normal amount of carbohydrates distribution in hepatocytes, (d) liver section of mice treated with LA7 showing normal distribution of PAS reaction in hepatocytes, (e) mice treated with LA5 and infected with S. Typhi showing marked improvement in PAS reaction in hepatic cells, (f) challenged mice with S. Typhi and treated with LA7 showing normal distribution of carbohydrates reaction beside patches of PAS reaction in portal areas (PAS X 400).

A photomicrograph of section of intestine shows: (a) control mice showing normal mucosa is thrown into finger-like villi, (b) mice challenged with S. Typhi showing shortening of the villi with destruction and exfoliation in the epithelial lining of villi which is replaced by pyknotic nuclei, with edema, (c) mice treated with LA5 showing normal tall finger-like intestine villi with crypts in between some enterocytes nuclei are pyknotic, (d) intestinal section of mice treated with LA7 showing normal tall finger-like intestine villi with some damaged crypts and congested blood capillaries, (e) mice treated with LA5 and infected with S. Typhi showing surface columnar cells and some villi contain vesicular nuclei and regular brush border, (f) challenged mice and treated with LA7 showing normal finger-like projection of the villi and crypts of Leiberkuhn between the bases of the villi nearly similar to the control.

A photomicrograph of section of intestine showing: (a) control mice showing normal intestine villi stained magenta red color in the mucous globules of goblet cells, brush border, and in connective tissues, (b) mice challenged with S. Typhi showing few goblet cells and interrupted stained brush border, (c) mice treated with LA5 showing normal magenta red stained mucous globules of goblet cells and PAS positive reaction of brush border, (d) intestinal section of mice treated with LA7 showing normal distribution of mucin in villi goblet cells and brush border, (e) mice treated with LA5 and infected with S. Typhi showing many goblet cells with variable size and various degree of PAS reaction in localized area, (f) challenged mice treated with LA7 showing the noticeable distribution of PAS positive reaction in goblet cells and striated brush border (PAS X 400).
Discussion
Recently, there has been a great interest in products with natural origin. In the same line, wide range of lactobacillus strains were isolated from dairy products and re-evaluated as a valuable source of novel therapeutic tools. The results of the current study revealed that eight lactic acid were isolated from dairy products including Lactobacillus plantarum (LA5) and Lactobacillus paracsi (LA7). The results of the present study were in agreement with previous studies that mentioned dairy products are rich in probiotics from different species.22,23 Isolated Lactobacillus, LA5 and LA7, exhibited significant antimicrobial activity against different pathogens. Antimicrobial activity of lactobacilli may be due to production of organic acids, hydrogen peroxide, bacteriocins, and bacteriocin-like substances. 24 However, the data of the present work revealed that lactobacilli isolates showed bactericidal activity in a wide range of acidic pH and enhanced by the reduction of PH with greatest bactericidal activity in PH 2. The results of the current study imply that LA5 and LA7 have a broad-spectrum antimicrobial activity and confer of pathogenic bacteria in various conditions. 25 In the present study, serum of infected mice with S. typhi showed positive Widal agglutination test. In the present study, administration of LA5 or LA7 counteracts the pathogenic effect resulted from Salmonella infection. These results may be due to antagonistic activity of lactobacillus strains against typhoid infection. 26 The lactobacilli succeeded to get rid of salmonellosis based on its phagocytic and immunostimulant activity against typhoid antigen. 27 The results of the current study showed enteric salmonellosis induced severe changes of TNF and IL-1β; they are produced by macrophages and play a major role in inflammation and tumor progression. 28 These results were in agreement with a previous study that reported apoptotic role of salmonella infection to macrophage in order to skip from the detection induced by host innate immune system. 29 Earlier studies reported that salmonellosis enhance production of TNF, IL-1, IL-8, and caspase-3 from human dendritic cells. 30 On the other hand, animals treated with lactic acid bacteria exhibit no significant difference in comparison to control group that indicated highly safety profile of probiotics. Mice treated with lactobacilli infected with S. typhi showed a significant reduction of TNF and IL-1β although still more than the control group. In addition, Lactobacillus paracsi showed pronounced restoration of serum level of TNF in comparison to Lactobacillus plantarum treated mice. These results were in the same line of previous studies that describe anti-inflammatory activity of Lactobacillus paracsi and alleviate symptoms of patients with rheumatoid arthritis. 31
The modulator effect of lactobacilli may be attributed to distinct maintenance of immune homeostasis and significant adaptation of immune response. 32 The histological results reported in the current study confirmed the biochemical results and indicated that salmonella typhi induced severe histological changes of hepatic and intestinal tissues. Similar changes in the hepatic and intestinal tissues have been previously documented. 33 On the other hand, lactobacilli succeeded to reverse severe histological changes in the intestinal tract that induced by typhoid infection more or less like control. These results were in agreement with previous studies that reported lactobacilli counteract the epithelial changes of the intestinal villi and modulate the liver architectures.34,35
It can be concluded that Egyptian dairy products are rich with lactobacillus strains and exhibit antimicrobial and immunomodulatory efficacy against a wide range of enteropathogenic effects of S. typhi due to its bactericidal and anti-inflammatory properties. These results suggest the importance of using dairy products to reduce the virulence of salmonella infection.
Footnotes
Acknowledgements
The authors thank Professor Nabila Hassan, Pathology Department, National Research Centre, Egypt for her precious support during investigation and writing comments of pathological examinations.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.