The study aimed to detect CD8+CD25+FoxP3brigh Tregs and investigate their possible association with selected lung function values. CD8+CD25+FoxP3brigh Tregs were detected by flow cytometry in the peripheral blood of 25 patients with severe asthma (SA), 25 patients with mild-to-moderate asthma (MA), and 25 age-matched healthy donors (NC). The percentages of CD8+CD25+FoxP3brigh Tregs of the patients with severe (3.4 ± 4.55), and mild-to-moderate asthma (7.5 ± 8.15), were markedly lower than those of controls (12.1 ± 13.2). The mean forced expiratory volume in 1 s (FEV1) % predicted value in severe asthma subpopulation was significantly lower (67.05 ± 15.98%) when compared with that of mild-to-moderate asthma subgroup (87.71 ± 16.12%). Interestingly, the percentages of CD8+CD25+FoxP3brigh Tregs correlate with mean peak expiratory flow (PEF)% predicted values in severe (r = 0.7, P <0.01) and mild-to-moderate (r = 0.73, P <0.01) asthma. In contrast, this parameter was positively correlated with FEV1% predicted values in the severe asthmatics only (r = 0.71, P <0.01). In summary, this study establishes a link between the percentage of CD8+CD25+FoxP3brigh Tregs and selected lung function parameters, suggesting that this parameter has potential as a marker for inflammation and airflow obstruction.
Asthma is a chronic inflammatory disorder characterized by airway hyper-responsiveness, persistent inflammation, and increased influx of inflammatory cells into the airways. While the role of T helper (Th) cells in allergic asthma has already been well established, convincing lines of evidence suggest that a subpopulation of CD8+ T cells secretes T helper (Th)-2 cytokines upon antigen challenge,1 and has crucial implications for the development of chronic airways inflammation. Furthermore, it has been previously hypothesized that the persistent airflow limitation during asthma exacerbation affects mainly the central airways. Recent reports indicate that this process affects the entire lower respiratory system.2 Spirometry and peak flow monitoring are widely used to assess or monitor lung health. However, over the past few decades, credible lines of evidence indicate that most lung function parameters such as forced expiratory volume in (1 s) (FEV1) and peak expiratory flow (PEF) are not reliable,3 in that they do not guarantee a thorough assessment of the entire lower respiratory tract4 or are subject to intraindividual variation. These limitations might account for approximately 30% falsely diagnosed5 and 54% undiagnosed6 asthma cases. In addition, some patients with mild or poorly controlled asthma may have ‘normal’ pulmonary physiological indices. Furthermore, PEF does not always ensure an accurate clinical assessment of asthma severity7 and forced expiratory flow does not accurately reflect small airway inflammation.8 These unmet clinical needs of existing lung function tests have motivated a search for better strategies to assess the degree of airflow obstruction. In this perspective, several non-invasive markers of airway limitation, including serum levels of eosinophil cationic protein (ECP),9 exhaled gases10 have been targeted. Notwithstanding, to date, no gold standard for assessing small airway limitation has ever been established. The report of the relevance of multiple Tregs and related transcription factors as the cornerstone of the maintenance of immune tolerance and the development of chronic inflammation has provided another major breakthrough in our understanding of asthma pathobiology. In this contest, a plethora of studies have revealed that various Tregs can suppress the development and the function of other effector cells. A marker of T-cell activation, the high affinity receptor for IL (interleukin)-2 receptor alpha subunit (IL- 2Rα) also known as CD25, is a phenotypic marker for naturally occurring Tregs. Constitutive expression of CD25 either alone or in combination with the transcription factor FoxP3 has been widely used to identify this subset of T cells endowed with the capacity to suppress effector T cells. Cow’s milk protein-specific T cells expressed higher expression levels of CD25 than similar cells from milk-tolerant children.11 Circulating CD4+CD25+ Tregs have been shown to circumvent or reverse the allergic inflammation.12 Heme oxygenase-1 (HO-1) appears to protect against allergic airway inflammation through enhancement of CD4+CD25high Tregs.13 Although most of these studies have mainly unlocked the active role of CD4+ Tregs subset in maintaining immune homeostasis, large-scale studies on the implication of CD8+CD25+FoxP3brigh Tregs subset in allergic inflicted inflammation are rather, relatively scarce. We have previously reported the diagnostic value of Fractional exhaled nitric oxide (FENO) in childhood asthma.14 In a subsequent study, we claimed that the decreased relative proportion of CD8+CD25+FoxP3brigh Tregs in peripheral blood results in asthma.15 Because of the relevance of this subset to persistent airway inflammation, we have extended our work to establish a positive correlation between the relative proportion of CD8+CD25+FoxP3brigh Tregs and two selected lung physiological indexes in asthma. With regard to clinical implications, we hypothesize that flow cytometric assessment of CD8+CD25+FoxP3brigh Tregs may represent a novel biomarker for airway inflammation in routine clinical setting and might have diagnostic value in asthma.
Material and methods
Ethics approval and screening
An institutional review board approved the study protocol (RNN/17/09/KE), before study initiation. All participants provided written informed consent. This randomized study enrolled non-smokers who were previously diagnosed and treated for asthma. All patients were asymptomatic on study entry. Patients with cardiovascular disorders, respiratory infection, chest or abdominal pain, and those who could not undergo spirometry have also been excluded from the study. A paper and pencil version of Asthma Control Test (ACT)16 was used to assess the level of asthma control. Patients underwent clinical examination and spirometry with airways reversibility test before the blood samples collection. The degree of airflow limitation was assessed by means of spirometry as previously described elsewhere.17 Lung function indexes namely forced vital capacity (FVC), FEV1, and FEV1/FVC were measured three times, and the best results were selected. The methacholine challenge test was used to assess patients with asthma symptoms but normal lung function values. Patients also underwent skin-prick test to common allergens and specific serum immunoglobulin E (IgE) was measured. Atopy was defined by either a positive skin test to at least one of a selected panel of allergens (Allergopharma), or detectable allergen-specific IgE. We performed a drop-out analysis. Drop-out was defined as failure to appear for a scheduled follow-up visit after initial screening and study enrolment.18
Study population
We retrospectively enrolled 50 allergic asthma (AA) subjects registered with the Asthma and Allergy outpatient clinic of the Medical University of Lodz, Poland. Twenty-five age-matched healthy non-smokers served as controls (NC). Diagnosis of bronchial asthma was made based on history of asthma symptoms, and reversible airflow limitation. We used the ENFUMOSA criteria to assess disease severity as previously described elsewhere.19
Antibodies and reagents
The following monoclonal antibodies: anti-human CD8-AmCyan (clone SK1, Mouse IgG1, k), anti-Human CD25-FITC (clone M-A251, Mouse IgG1, k) were purchased from BD Pharmingen. Anti-Human FoxP3-APC (clone 236A/E7, Mouse IgG1, k) and permeabilization buffers used for intracellular staining, were procured from eBioscience. Histopaque 1077 was procured from Sigma-Aldrich. Antibody capture micro-beads and the human CD8+ T cells enrichment kit were obtained from BD Biosciences.
Isolation of peripheral blood mononuclear cells (PBMC)
PBMC were purified from heparinized peripheral blood using density gradient centrifugation (2500 rpm, 30 min) over Histopaque-1077. PBMC were collected from the buffy coat located at the Histopaque/serum interface and washed in PBS.
Extraction of CD8+ T cells from PBMC
CD8+ T cells were negatively isolated from PBMC using Human Naive CD8+ T Cell Enrichment kit. A total of 100 μL (106 cells) were incubated with 5 μL biotinylated monoclonal antibodies directed against all types of PBMCs except the CD8+ T-cells population, at 4°C for 15 min. The labeled cells were subsequently washed and centrifuged for 10 min. Thereafter, the cells suspension (106) was incubated with streptavidin micro-beads (7.5 μL) at 4°C for 30 min. CD8+ T cells were purified by immunomagnetic separation using the BD IMagnet™ system. The isolated cells were incubated with 100 µL of anti-CD8 antibodies for 20 min on ice and analyzed by flow cytometry. The purity of the enriched CD8+ T cells fraction was more than 93%.
Immunostaining
CD8+ T cells (106 cells/100 µL) were washed twice in BD Pharmingen Stain Buffer (FBS) and incubated with 10 μL anti-human CD8 and anti-Human CD25 for 30 min at 4°C in the dark. Cells were re-suspended in wash buffer and then incubated with 2 mL of 1× Human FoxP3 Buffer A for 10 min in the dark at 4°C. After incubation, cells were washed twice with 2 mL of Stain Buffer (FBS) and centrifuged. The pellet was washed and incubated with 0.5 mL of 1× Human FoxP3 Buffer C for 10 min. Cells were washed with 2 mL Stain Buffer (FBS) and incubated with anti-Human FoxP3 for 30 min at 4°C. Cells were re suspended in 500 μL staining buffer, and fluorescent events were analyzed by flow cytometry.
Flow cytometry
CD8+ T cells were isolated using Magnetic-activated cell sorting (MACS), and stained with a panel of antibodies according to manufacturer’s protocol, as previously described above. The average TAT for the clinical samples was 12 h. Cells were acquired on a FACS CANTO II flow cytometer (BD Biosciences). A minimum of 50,000 fluorescent events were captured for each sample. Lymphocyte gates were set based on forward scatter vs. side scatter, excluding cell debris. To analyze surface CD25 and intracellular FoxP3 expression on circulatory CD8+ T cells, cells were gated as CD8+CD25+ events and the expression of FoxP3 was quantified within CD8+CD25+ cells. Quadrants of dot-plots were set for each antibody (Figure 1). Automatic compensation for spectral overlap was performed electronically to minimize fluorescence spillover, using antibody capture beads. Gating strategy was determined by comparison to Fluorescence Minus One (FMO) control. Gating was standardized within individual samples to generate a fully comparative data. Data were analyzed using FACSDiva and FCS softwares (BD Biosciences), and are representative for experiments performed with cells obtained from 50 patients with allergic asthma and 25 healthy controls. All experiments were run by the same operator, with the same instrument and compensation settings, to ensure data comparability.
Gating strategy for CD8+CD25+FoxP3brigh cells. To define and quantify circulating CD8+CD25+FoxP3brigh cells, CD8+ T cells from allergic asthma (AA) and healthy (NC) subjects were purified from peripheral blood samples and stained for coexpression of CD8, CD25, and FoxP3. CD8+ T cells were gated out of lymphocytes population, and the percentage of CD8+ cells co expressing CD25+ and FoxP3brigh cells among the selected CD8+ T cells was determined.
Statistical analysis
Data were analyzed as either, mean ± standard deviation (SD), or median ± interquartile (IQR). Analysis was performed using Statistica Pl. v. 8.0 software (StatSoft Sp. z o. o). The Mann-Whitney U test was used for testing differences among variables. Correlations between parameters were assessed using the Spearman rank correlation coefficient. Statistical significance was set at P <0.05.
Results
Population characteristics
To investigate whether levels of CD8+CD25+FoxP3brigh Tregs correlate with lung function parameters, adults with allergic asthma (AA) in stable condition, under as needed (PRN) asthma drug regimens, were selected, and completed a baseline visit. Age-matched healthy subjects served as controls (NC). The patients were divided equally and randomly into two groups: severe asthma (SA) (n = 25) and mild-to-moderate asthma (MA) (n = 25). Overall, 48% of severe asthma patients were men, 52% were women. Similarly, 52% of mild-to-moderate asthma patients were men, 48% were women. Likewise, 52% of healthy subjects were men whereas 48% were women. The severe and mild-to-moderate asthma groups had a mean age of 48.0 ± 14.00 years and 42.0 ± 13.00 years, respectively. The healthy controls group was characterized by an average age of 48.5 ± 14.5 years. The flow cytometry analysis and lung function test were performed between two consecutive visits. The mean percentage changes of FEV1 from baseline in severe asthma and mild-to-moderate asthma patients were 67.05 ± 15.98 and 87.71 ± 16.12%, respectively. Drop-out of clinical trials has been associated with higher degrees or frequencies of distress associated with disease-related symptoms such as dyspnea. We hypothesized that patients with severe asthma exacerbation are more likely to drop out of the study than their peers with mild-to-moderate asthma phenotype. Surprisingly, all patients with asthma, irrespective of disease severity had all follow-up visits. Wheal flare with a mean diameter of at least 3 mm (or 7 mm2 area), after 15 min reading was considered positive. Allergen-specific IgE values of ⩾0.35 kU/L were considered positive. Atopy was ruled in based on positive reaction to at least one allergen or evidence of increase specific serum IgE. Allergen poly-sensitization was defined as sensitization to more than two allergens-specific IgE, regardless of allergen class. The presence of persistent obstructive airflow limitation was confirmed whenever FEV1/FVC was lower than 0.70 and FEV1 was reduced more than FVC. All patients had airway hyper-responsiveness, and reversible airflow obstruction, which was defined as an increase in FEV1 or FVC of at least 200 mL and 12% from patient’s baseline values, in response to inhaled bronchodilator administration. Moreover, an improvement in PEF of at least 60 L/min or 20% after inhalation of a bronchodilator indicates asthma. Patients with mild-to-moderate stable asthma were receiving budesonide or beclamethasone 800 μg once daily and fluticasone 500 μg once daily as needed. Patients with severe asthma were receiving high doses of inhaled corticosteroids, namely budesonide ⩾1600 μg/day or beclomethasone, and futicasone 800 μg/day or equivalent. Some severe asthma patients were receiving long-term therapy with inhaled long-acting beta-receptor agonist (LABA), including formoterol 9 μg bid or salmeterol 50 μg bid, alternatively oral theophylline (200–300 mg ⩽2 daily).
Decreased relative count of CD8+CD25+FoxP3righ Tregs subset in peripheral blood is associated with asthma and determines diseases severity
Numerous markers have been used to delineate natural and inducible Treg cell subpopulation. Particularly, CD25 and the transcription factor, FoxP3, have predominately and conventionally been used to define natural Tregs. We and others20 have demonstrated previously that several CD8+ Tregs have a role in allergic asthma development. We extend these preliminary findings by showing that CD8+CD25+FoxP3brigh Tregs populate the peripheral blood of healthy and allergy asthma subjects. The levels of CD8+CD25+FoxP3brigh Tregs were significantly decreased in severe asthmatics (3.4 ± 4.55) and mild-to-moderate asthmatics (7.5 ± 8.15) compared to controls (12.1 ± 13.2). Subjects with severe asthma had lower percentages of CD8+CD25+FoxP3brigh Tregs than those with mild-to-moderate asthma. In comparison with controls, a statistically significant, lower frequency of CD8+CD25+FoxP3brigh Tregs was seen in the severe asthma subjects (P <0.0001) (Figure 2).
Comparative analysis of the relative levels of circulatory CD8+CD25+FoxP3brigh Tregs between allergic asthma subjects and healthy controls.
Correlation between CD8+CD25+FoxP3brigh Tregs and lung function parameters
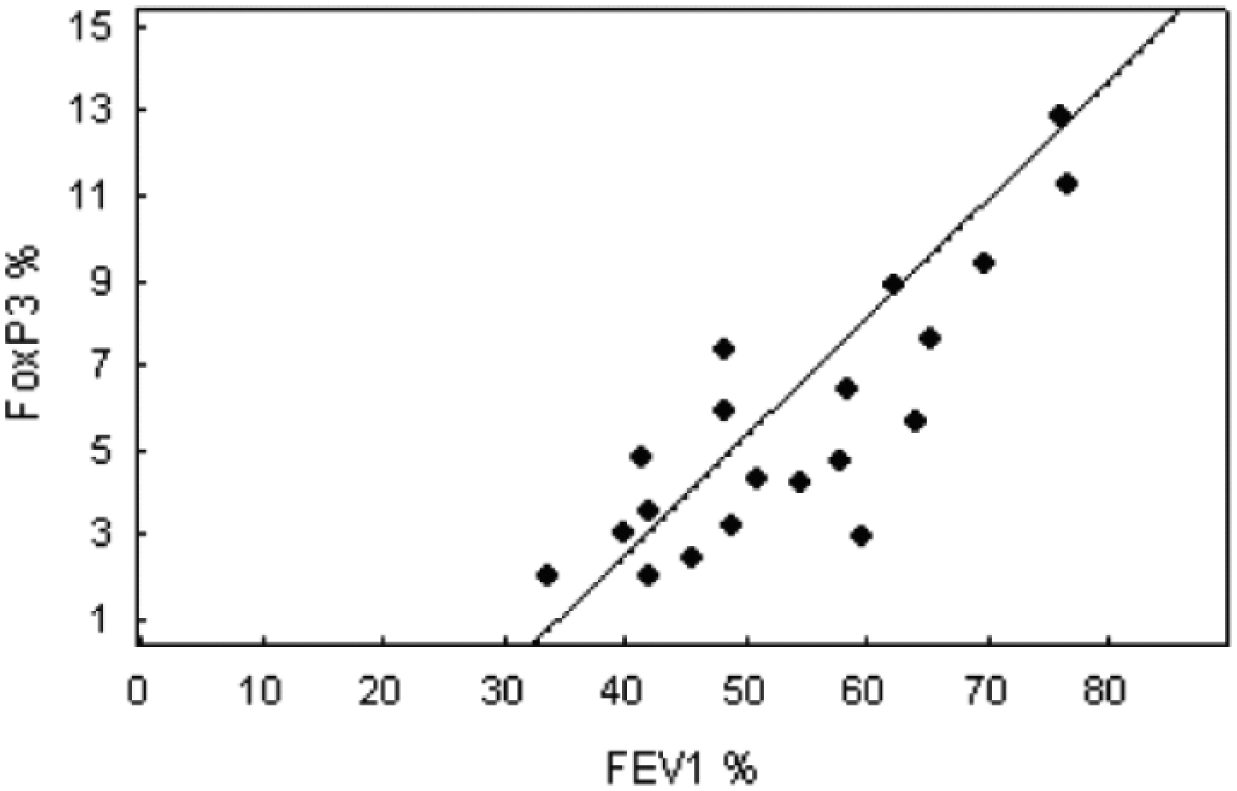
Several reports have demonstrated the effects of allergy-mediated inflammation on the dynamic of several circulatory CD8+ T-cells profile, and lung function variables, such as the mean forced expiratory volume in 1 s (FEV1) % predicted values, and the mean peak expiratory flow (PEF) % predicted values. The present study complements and extend these preliminary findings by revealing a positive correlation between the level of circulatory CD8+CD25+FoxP3brigh Tregs and FEV1% predicted values (r = 0.71, P <0.01) (Figure 3) in the severe asthma group. This parameter correlated positively with mean peak expiratory flow (PEF) % predicted values (r = 0.7, P <0.01) in this population (Figure 4). Furthermore, we observed a positive association between %PEF and %CD8+CD25+FoxP3brigh Tregs (r = 0.73, P <0.01) in the mild-to-moderate asthma subgroup (Figure 5).
Positive correlation between the relative value of CD8+CD25+FoxP3brigh cells and the mean forced expiratory volume in 1 s (FEV1) % predicted values in severe asthma (r = 0.71, P <0.01).
Positive correlation between the percentages of CD8+CD25+FoxP3brigh cells and the mean peak expiratory flow (PEF) % predicted values in patients with severe asthma (r = 0.70, P <0.01).
Positive correlation between the frequencies of CD8+CD25+FoxP3brigh cells and the mean PEF % predicted values in patients with mild to moderate asthma (r = 0.73, P<0.01).
Discussion
Here we detect CD8+CD25+FoxP3brigh Tregs in peripheral blood and suggest the potential importance of this CD8+ T-cell subset in allergies. In this context, we add new findings to existing data on the importance of Treg-cell subsets in asthma and atopic diseases to suggest that CD8+CD25+FoxP3brigh Tregs subset has diagnostic and prognostic value as a biomarker for asthma. First, this study demonstrated that the relative count of CD8+CD25+FoxP3brigh Treg of total peripheral blood mononuclear cells is decreased in asthma subjects compared with healthy subjects. These findings support and extend results of previous studies, including ours describing the relevance of Treg cell dysfunction in allergy and asthma. Defects in the proportion and function of Treg cells are thought to result in the development of immune dysregulation, polyendocrinopathy, enteropathy, X-linked (IPEX), a disorder characterized by an increased serum IgE level and a skewed Th2-mediated immune response. Lately, CD8+ Tregs cells have emerged as inhibitor of the cellular proliferation of naive and effectors CD4+ T cells.21 In the same contest, a recent study has demonstrated that the Toll-like receptors agonists can suppress allergen-induced Th2 immune response by increasing the frequency of CD8+Foxp3+ Tregs.20 Although our findings lend support to these preliminary investigations, they contrast with other studies that reported normal proportion of Tregs in the peripheral blood of asthmatic patients in stable condition.22 Further retrospective, cohort studies are necessary to confirm or disprove our findings. Another main finding from our study is the significant decrease in CD8+CD25+FoxP3brigh Tregs level in the severe asthma population when compared with that of the group of subjects with mild-to-moderate asthma, suggesting that the decreased frequency of CD8+CD25+FoxP3brigh Tregs probably determines disease severity. Giving that inhaled corticosteroids improve asthma control through increased Treg cell frequency,23 it is rational to reason that the higher the steroid dose and the longer the frequency and duration of therapy, the greater the proportion of Treg cells. Thus, consistent with a previous study that have demonstrated a reduced frequency of CD4+CD25high FoxP3+ Tregs in steroid-naive asthmatics compared with those receiving steroid treatment,24 the unexpected decrease in the proportion of Treg cells in severe asthma let us suggest that disease status, rather than different treatment protocols, is responsible for discrepant level of Tregs. Although few studies have revealed an association between Treg cells proportion and asthma pathobiology, in terms of abnormal lung function values and IgE levels,25 to the best of our knowledge, the link between CD8+CD25+FoxP3brigh Tregs and lung function values has not yet been established. Along with other colleagues, we have previously reported the correlation between fractional exhaled nitric oxide (FENO) and airway hyper-responsiveness as measured by lung function values in childhood asthma.14 The present study extends these findings by demonstrating a positive correlation between the frequencies of ex vivo CD8+CD25+FoxP3bright Tregs from asthma subject and either one or two lung physiological indexes. Although our data further extend those of a previous study,26 they are in contrast to findings from other studies which have failed to establish a correlation between these cells and pulmonary function values.27 Further clinical studies comparing the pathological mechanisms of severe and mild-to-moderate asthma have revealed differences between these phenotypes, with regard to patterns of abnormal inflammatory defects and related structural changes affecting the airways.28 Extending these previous works, here, we show that the relative proportion of CD8+CD25+FoxP3brigh Tregs correlates positively with the two selected lung function values in severe asthma subjects, supporting a previous study that examined the link between PEF and FEV1 % predicted values in most asthmatic patients, mainly those with severe asthma.29 Surprisingly, the percentage of CD8+CD25+FoxP3brigh Tregs correlated with PEF % predicted values only in the mild-to-moderate asthmatic subpopulation. These data further support an earlier report that demonstrated that FEV1 and PEF may not be used interchangeably as markers of less severe airflow limitation.30 Moreover, they suggest that clinical targeting of CD8+CD25+FoxP3brigh Tregs may be challenging. We argue that, as a single biomarker, this parameter has only weak diagnostic value in mild-to-moderate asthma. The main strength of the present study is the relatively even distribution of the study population with regard to age, sex, and asthma phenotypes. Another strength of our case-control study is that all procedures were performed by the same physician or scientist. It is also worth mentioning that our study has few caveats. In terms of methodology, one disadvantage is that the study was restricted to the Polish population, and a fairly small area, Lodz, thus, the data cannot be generalized to the larger population in another real-world setting. Despite these limitations, we believe that our findings are robust, and worth sharing.
Conclusion
In sum, the present study establishes the link between CD8+CD25+FoxP3bright Tregs and selected lung function parameters in asthmatics. Furthermore, it compiles our own findings with the limited existing data regarding CD8+Treg cell and potential inflammatory markers of asthma to suggest that assessing CD8+CD25+FoxP3brigh Tregs proportions, by means of flow cytometry, as add-on marker, may improve current approach to detect and monitor asthma. Further prospective studies with a larger sample size are underway to confirm or challenge our findings.
Footnotes
Acknowledgements
The authors thank all the participants of this study.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported by grant from the Polish Academy of Sciences grant KBN nr N N402 517339.
References
1.
CoulterEMJenkinsonCFarrellJ. (2010) Measurement of CD4+ and CD8+ T-lymphocyte cytokine secretion and gene expression changes in p-phenylenediamine allergic patients and tolerant individuals. Journal of Investigative Dermatology130: 161–174.
2.
Ramos-BarbónDParra-ArrondoA (2011) Inflammation and remodeling of the distal airways: Studies in humans and experimental models. Archivos de Bronconeumologia47: 2–9.
3.
NewtonMFO’DonnellDEForkertL (2002) Response of lung volumes to inhaled salbutamol in a large population of patients with severe hyperinflation. Chest121: 1042–1050.
4.
ControliMBousquetJFabbriLM. (2010) The small airways and distal lung compartment in asthma and COPD: A time for reappraisal. Allergy65: 141–151.
5.
FunstonWHiggingsB (2014) Improving the management of asthma in adults in primary care. Practitioner258: 15–19.
6.
JoséBPCamargosPACruz FilhoÁA. (2014) Diagnostic accuracy of respiratory diseases in primary health units. Revista de Associacao Medica Brasileira60: 599–612.
7.
ReddelHJenkinsCWoolcockA (1999) Diurnal variability—time to change asthma guidelines?BMJ319: 45–47.
8.
SutherlandERMartinRJBowlerRP. (2004) Physiologic correlates of distal lung inflammation in asthma. Journal of Allergy and Clinical Immunology113: 1046–1050.
9.
NiimiAAmitaniRSuzukiK. (1998) Serum eosinophil cationic protein as a marker of eosinophilic inflammation in asthma. Clinical & Experimental Allergy28: 233–240.
10.
WarkPAGibsonPG (2003) Clinical usefulness of inflammatory markers in asthma. American Journal of Respiratory Medicine2: 11–19.
11.
SchadeRPVan Ieperen-Van DijkAGVersluisC. (2002) Cell-surface expression of CD25, CD26, and CD30 by allergen-specific T cells is intrinsically different in cow’s milk allergy. Journal of Allergy and Clinical Immunology109: 357–362.
12.
OstroukhovaMRayA (2005) CD25+ T cells and regulation of allergen-induced responses. Current Allergy and Asthma Reports5: 35–41.
13.
XiaZWZhongWWXuLQ. (2006) Heme oxygenase-1-mediated CD4+CD25high regulatory T cells suppress allergic airway inflammation. Journal of Immunology177: 5936–5945.
14.
GrzelewskiTWitkowskiKMakandjou-OlaE. (2014) Diagnostic value of lung function parameters and FeNO for asthma in schoolchildren in large, real-life population. Pediatric Pulmonology49: 632–640.
15.
EusebioMKunaPKraszulaL. (2014) Allergy-related changes in levels of CD8+CD25+FoxP3(bright) Treg cells and FoxP3 mRNA expression in peripheral blood: the role of IL-10 or TGF-beta. Journal of Biological Regulators & Homeostatic Agents28: 461–470.
16.
ShatzMSorknessCALiJT. (2006) Asthma Control Test: Reliability, validity, and responsiveness in patients not previously followed by asthma specialists. Journal of Allergy and Clinical Immunology117: 549–556.
17.
NèveVMatranRBaquetG. (2012) Quantification of shape of flow-volume loop of healthy preschool children and preschool children with wheezing disorders. Pediatric Pulmonology47: 884–894.
18.
YennurajalingamSKangJHChengHY. (2013) Characteristics of advanced cancer patients with cancer-related fatigue enrolled in clinical trials and patients referred to outpatient palliative care clinics. Journal of Pain and Symptom Management45: 534–541.
19.
PietruczukMEusebioMKraszulaL. (2012) Phenotypic characterization of ex vivo CD4+CD25highCD127low immune regulatory T cells in allergic asthma: Pathogenesis relevance of their FoxP3, GITR, CTLA-4 and FAS expressions. Journal of Biological Regulators & Homeostatic Agents26: 627–639.
20.
TsaiYGYangKDNiuDM. (2010) TLR2 agonists enhance CD8+Foxp3+ regulatory T cells and suppress Th2 immune responses during allergen immunotherapy. Journal of Immunology184: 7229–7237.
21.
SinghAMDahlbergPBurmeisterK. (2013) Inhaled corticosteroid use is associated with increased circulating T regulatory cells in children with asthma. Clinical and Molecular Allergy11: 1.
22.
ShiHZLiSXieZF. (2004) Regulatory CD4+CD25+ T lymphocytes in peripheral blood from patients with atopic asthma. Clinical Immunology113: 172–178.
23.
SinghAMPaul DahlbergPBurmeisterK. (2013) Inhaled corticosteroid use is associated with increased circulating T regulatory cells in children with asthma. Clinical and Molecular Allergy11: 1.
24.
ProvoostSMaesTvan DurmeYM. (2009) Decreased FOXP3 protein expression in patients with asthma. Allergy64: 1539–1546.
25.
NguyenKDFohnerABookerJD. (2008) XCL1 enhances regulatory activities of CD4+ CD25(high) CD127(low/-) T cells in human allergic asthma. Journal of Immunology181: 5386–5395.
26.
KotsianidisINakouEBouchliouI. (2009) Global impairment of CD4+CD25+FOXP3+ regulatory T cells in idiopathic pulmonary fibrosis. American Journal of Respiratory and Critical Care Medicine179: 1121–1130.
27.
LeyBBrownKKCollardHR (2014) Molecular biomarkers in idiopathic pulmonary fibrosis. American Journal of Physiology – Lung Cellular and Molecular Physiology307: L681–691.
28.
CaramoriGPanditAPapiA (2005) Is there a difference between chronic airway inflammation in chronic severe asthma and chronic obstructive pulmonary disease?Current Opinion in Allergy and Clinical Immunology5: 77–83.
29.
GruptaDAggarwalANSubalaxmiMV. (2000) Assessing severity of asthma: Spirometric correlates with visual analogue scale (VAS). Indian Journal of Chest Disease and Allied Science42: 95–100.
30.
AggarwalANGuptaDJindalSK (2006) The relationship between FEV1 and peak expiratory flow in patients with airways obstruction is poor. Chest130: 1454–1461.