
Abstract
Background
Cluster headache is a primary headache disorder characterized by bouts with circadian and circannual patterns. The CLOCK gene has a central role in regulating circadian rhythms. Here, we investigate the circannual CLOCK expression in a population of cluster headache patients in comparison to matched controls.
Methods
Patients with cluster headache were sampled two to four times over at least one year, both in or outside bouts, one week after each solstice and equinox. The expression of CLOCK was measured by quantitative real-time polymerase chain reaction (RT-PCR) in the peripheral blood.
Results
This study included 50 patients and 58 matched controls. Among the patient population, composed of 42/50 males (84%) with an average age of 44.6 years, 45/50 (90%) suffered from episodic cluster headache. Two to four samples were collected from each patient adding up to 161 samples, 36 (22.3%) of which were collected within a bout. CLOCK expression for cluster headache patients was considerably different from that of the control population in winter (p-value mean = 0.006283), spring (p-value mean = 0.000006) and summer (p-value mean = 0.000064), but not in autumn (p-value mean = 0.262272). For each season transition, the variations in CLOCK expression were more pronounced in the control group than in the cluster headache population. No statistically significant differences were found between bout and non-bout samples. No individual factors (age, sex, circadian chronotype, smoking and coffee habits or history of migraine) were related to CLOCK expression.
Conclusions
We observed that CLOCK expression in cluster headache patients fluctuates less throughout the year than in the control population. Bout activity and lifestyle factors do not seem to influence CLOCK expression.
Introduction
Clock genes are responsible for maintaining the intracellular timekeeping system, throughout the body, generating physiological rhythms, among other behavioral and brain functions (1,2). In particular, the CLOCK (Circadian Locomotor Output Cycles Kaput) gene encodes a transcription factor that interacts with BMAL1 (brain-and-muscle ARNT-like protein 1) protein and with the heterodimer PERs (period proteins)/CRYs (cryptochrome proteins) (3,4). These heterodimers activate and inhibit each other, producing a cell-autonomous oscillation of approximately 24 hours (4,5). CLOCK is strongly expressed in the brain regions responsible for controlling circadian rhythms, including the suprachiasmatic nucleus (SCN) in the hypothalamus (5 –7). SCN synchronizes the cellular clocks in the periphery generating circadian rhythms, mainly through the autonomic nervous system (6). Polymorphisms in circadian clock genes have been associated with several neuropsychiatric disorders such as autism spectrum disorder, schizophrenia, anxiety, major depressive disorder, bipolar disorder, attention deficit hyperactivity disorder and sleep architecture perturbation (7,8).
While the exact pathophysiology of cluster headache (CH) is still unknown, one of its main clinical features is the striking rhythmicity of attacks, with a circadian and circannual pattern (3). CH patients report a predictable circadian and circannual rhythmicity of attacks (82% and 56%, respectively) (9). In fact, most patients have cluster attacks at the same time each day, most often at night between midnight and 3 am, and at the same time every year, usually in the spring or autumn (10). The periodicity of attacks and the association with sleep disorders suggest that CH is a chronobiological disorder that might involve the SCN and circadian clock transcription-translation feedback loops (11,12). The underlying mechanism of how CLOCK could modulate CH via the hypothalamus is not known. However, complex genetic factors are likely involved in CH etiology (12), and thus CLOCK polymorphisms may affect the internal clock and lead to a mismatch between the internal clock and external time cues (3). Genome-wide association studies have implicated specific gene variants in CH susceptibility, although the candidate genes found await validation, and the molecular pathways and physiologic circuits involved remain poorly understood (1,13 –16).
We have previously found that levels of expression of the CLOCK gene in peripheral blood are altered in obstructive sleep apnea syndrome (OSAS) (17). Because patients with CH have a higher prevalence of OSAS, and the link between these conditions is observed during active cluster episodes (18,19), we asked whether CLOCK expression could also be compromised in CH. To test this possibility, we compared the seasonal pattern of CLOCK expression between CH patients and healthy controls, in peripheral blood as before (17), due to the known circannual rhythmicity of bouts (9). Data on the regulation of clock genes that accompany the variations of human circannual rhythms is scarce, but a postmortem study in human brains, which tested the expression of a large pool of genes in dorsolateral prefrontal cortices, identified seasonal patterns for the PER family of genes (20).
Here we explore the patterns of the circannual expression of CLOCK in a population of CH patients and matched controls. We hypothesize that differences in CLOCK gene expression may be associated with increased risk of CH, considering the impact of CLOCK in the hypothalamus and as a potential modulator of chronosynchronicity.
Methods
Population
A cohort of adult patients (>18 years) with episodic or chronic CH according to the ICDH-III beta (21) followed in a neurology outpatient clinic at a tertiary care hospital between 2016 and 2019 were included. Controls were healthy volunteers recruited from a checkup clinic, matched by sex and age. Exclusion criteria included severe neurological or psychiatric comorbidity (defined as conditioning labor and/or daily activities at the time of study), bipolar disorder, active infection, active oncological disease, any autoimmune disease, current allergen immunotherapy, pregnancy, lactation and inability to provide informed consent. Controls were recruited one to two days per month and had only one evaluation. Differences in age between patients and controls were tested with a chi-squared test and differences in sex between patients and controls were tested by Fisher's exact test.
Standard protocol approvals, registrations, and patient consent
The protocol was approved by the Hospital da Luz Ethical Committee and compliance with data protection regulations was obtained with the local Data Protection Officer. All patients and controls signed a written informed consent.
Evaluation protocol
After informed consent, we collected information regarding the clinical characterization of the disease for all patients, including the circadian and circannual pattern of the bouts in episodic CH and headache impact test (HIT-6) (22). Relevant information on potential influencing factors such as habits and routines, coffee, alcohol and tobacco consumption, sleep patterns (Epworth scale, Pittsburgh Sleep Quality Index and Horne-Ostberg Morningness-Eveningness Questionnaire) (23 –25), mood (Beck Depression Inventory, State-Trait Anxiety Inventory STAY-Y) (26,27) and suicidal ideation (Columbia Suicidality) (28), comorbidities and current medication were collected for patients and controls. Differences between patients and controls regarding regular smoking consumption, regular coffee consumption and regular alcohol consumption were tested by Fisher's exact test.
Sample collections
Patients had at least two and up to four assessments over the minimum period of one year. In each assessment, information concerning disease activity and current medication was compiled, and blood samples were collected. Patient assessments were scheduled for a three-day period starting from the 6th to the 8th day after each solstice and equinox in the northern hemisphere. Healthy volunteers were assessed and selected as controls in a period of one to two days in every month. Blood samples were collected from 8 am to 10 am for both patients and controls.
CLOCK expression
Quantitative real time polymerase chain reaction (RT-PCR) was used to quantify the expression of the gene CLOCK in peripheral blood cells of CH patients and compared with the expression of matched control volunteers. Blood was collected and peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll gradient (FICOLL-PAQUE PLUS GE Healthcare, catalog ner 17144002) and immediately resuspended in Trizol reagent (Invitrogen, catalog ner 15596026). Ribonucleic acid (RNA) was later extracted using the RNeasy Micro Kit (Qiagen, catalog ner 50974004) and synthesis of cDNA was performed with SuperScript II reverse transcriptase (Invitrogen, catalog ner 18064014). RNA levels were quantified in the QuantStudio 7 Flex Real-Time PCR machine (Applied Biosystems) and relative CLOCK expression of each sample was determined using the expression of the housekeeping gene GAPDH (glyceraldehyde 3-phosphate dehydrogenase). CLOCK levels for each patient were normalized to the mean CLOCK expression of the control population. The CLOCK primers used were: forward, TGCGAGGAACAATAGACCCAA; and reverse, ATGGCCTATGTGTGCGTTGTA. The GAPDH primers used were: forward, GAGTCAACGGATTTGGTCGT; and reverse, TTGATTTTGGAGGGATCTCG.
Data analysis
Differences in CLOCK expression between controls and patients for each season
Due to the lack of previous studies on this subject, we did not have any expectations regarding the size effects and therefore we did not perform power analysis on our cohort. We started by assessing, in the control population, the variation in CLOCK expression during the year. Control individuals had only one blood sample collected because we assumed that the levels of CLOCK expression throughout the year in healthy individuals could be used as the baseline for the expression of the gene. Values of expression were then indexed by the day of the year of their collection, and we used a Gaussian Process to assess the temporal variation of the CLOCK gene expression over the year. The correlation time scale that was estimated by the Gaussian Process was between 26 and 65 days (mean of 44 days). We used this statistical method to interpolate, in a non-parametrical way, the values of CLOCK expression and to retrieve a prediction of CLOCK expression for any particular day of the year, together with a measure of the uncertainty associated, referred to as natural variation. After fitting the Gaussian Process to the control population, we tested for differences in CLOCK expression between patients and controls. We asked whether CH patients would have any deviations to the predicted expression at any particular time of the year. For each season, we derived an equal number of synthetic controls (one set) to compare with the expression levels of the patients. p-values were calculated by Mann-Whitney tests by sampling 1000 sets of synthetic controls against the expression levels of the patients. We present the average of the p-values obtained for the 1000 tests, as well as the lower and upper limits of the confidence intervals.
Testing the effects of bouts, of being episodic patients or of having circannual patterns for CLOCK expression
We used multivariate regression analysis to assess, within the patient group, the effect of being in bout, as well as the effect of being an episodic patient or a patient with circannual pattern of disease on CLOCK gene expression. We present the p-values of the t-tests performed for each season and the adjusted p-values whenever appropriate. Multiple test corrections were performed using the Holm-Sidak method.
Differences in CLOCK expression across seasons
We made use of the paired nature of the CLOCK expression measurements of the patients to assess differences across consecutive seasons. For each season transition, we estimated an incremental change in expression, considering that the same individuals were measured. For the patient group, we used the average of the measurements. We used regression analysis to determine the slopes of the changes in expression associated with the transitions from one season to the consecutive season and we present the p-values of the t-tests performed, as well as the lower and upper limits of the confidence intervals.
Data availability
Data (anonymized) not published within the article is available and may be shared by request from any qualified investigator.
Results
Characterization of the CH patient and control populations
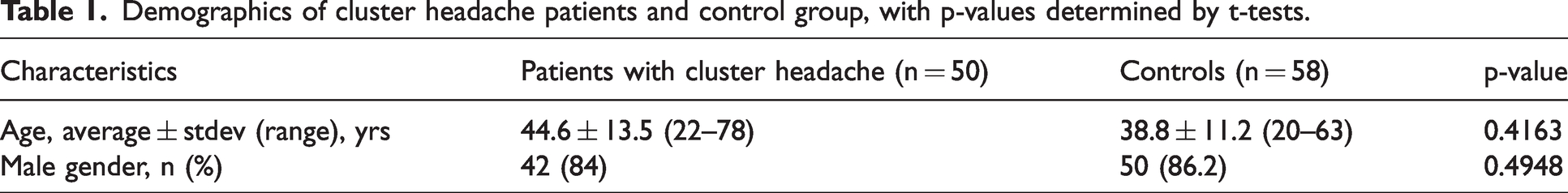
We included a total of 50 patients (84% male, average age of 44.6 ± 13.5 years) and 58 matched controls (86.2% male, average age of 38.8 ± 11.2 years) (Table 1). Our CH cohort had a very high male-to-female ratio (5.2:1), most likely because all the patients that were enrolled in our study were diagnosed before 2016 using criteria that preceded the 3rd edition of the International Classification of Headache Disorders (ICHD-3) in 2018 (29). Moreover, it was only in 2017 that differences in chronobiology between men and women with CH were identified (30). Patient and control groups were not statistically different in terms of age and sex (Table 1, p = 0.4163 and p = 0.4948, respectively). The age of the patients ranged from 22 to 78 years whereas the age of the controls ranged from 20 to 63 years (Table 1). When we distributed the ages of patients and controls per decade, we observed that the highest percentage of patients were in the 40–49 years group (28%) and the highest percentage of controls were in the 30–39 years group (32.3%) (Online Supplementary Table 1). More than half of the patients (62%) were current or previous smokers, 28% reported regular consumption of alcohol and 86% reported regular consumption of coffee (Table 2). Patients and controls had similar smoking, alcohol and coffee consumption habits (Online Supplementary Table 2, p = 0.1584, p = 0.1684 and p = 0.1685, respectively).
Demographics of cluster headache patients and control group, with p-values determined by t-tests.
Clinical characterization of cluster headache patients.
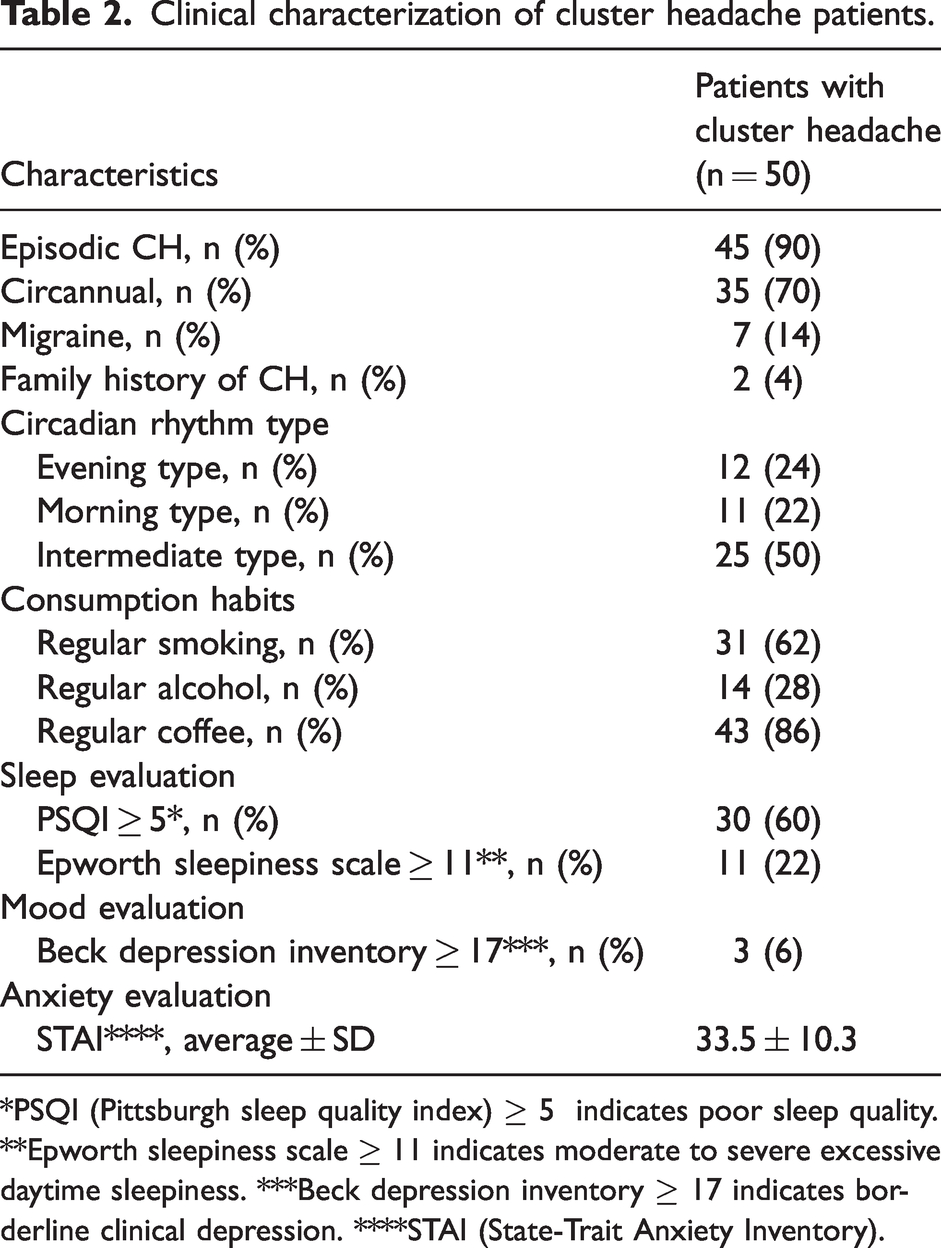
*PSQI (Pittsburgh sleep quality index) ≥ 5 indicates poor sleep quality. **Epworth sleepiness scale ≥ 11 indicates moderate to severe excessive daytime sleepiness. ***Beck depression inventory ≥ 17 indicates borderline clinical depression. ****STAI (State-Trait Anxiety Inventory).
We characterized several clinical aspects relevant for the disease (Table 2). Forty-five patients (90%) suffered from episodic CH, with a mean number of bouts of 8.9 ± 9.2 and a frequency of attacks during a bout of 1.9 ± 1.4 per day. Thirty-five patients (70%) had a circannual rhythmicity of bouts. Seven patients (14%) had personal history of migraine and two patients (4%) had family history of CH. Half of the patients had an intermediate type of circadian rhythm chronotype, whereas 24% of the patients had evening and 22% had morning chronotypes. Most patients (60%) had poor sleep quality (PSQI score ≥ 5) and 22% had moderate to severe excessive daytime sleepiness, as assessed by the Epworth sleepiness scale (≥11). Depression severity was assessed by the Beck Depression Inventory (BDI) and 6% of the CH patients reported borderline clinical depression (≥17). Anxiety was measured by the State-Trait Anxiety Inventory (STAI) and the average score was 33.5 ± 10.3.
Less than half of the patients (27.5%) had high anxiety levels, and depression was even less prevalent (7.8%). Only four patients had suicide risk (mild), coinciding with bout activity.
CLOCK gene circannual pattern of expression
Control samples were distributed throughout the year, with at least one blood sample every month (Online Supplementary Table 3). Each donor provided only one sample. Blood was processed, RNA was extracted from PBMCs and CLOCK gene expression was quantified by RT-PCR in these PBMCs as done before in a previous study (17). We tested CLOCK gene expression for all control samples and the results were normalized (relative expression) to the expression of the housekeeping gene GAPDH. We then used a Gaussian Process to statistically model the temporal variation of the CLOCK gene expression in the control population, which allowed us to extrapolate CLOCK expression for any particular day of the year (Figure 1; measured control samples are represented as black dots). With this, we attempted to understand the “natural” variation of the control population concerning CLOCK expression during the year. We observed that the expected expression of CLOCK gene fluctuated throughout the year (Figure 1, thick red line) and we also determined the associated uncertainty (Figure 1, red areas above and below the thick red line). According to our model, the highest CLOCK expression is expected in spring, whereas the lowest is expected in summer. Spring is also the season where CLOCK expression changes the most, with the least variations observed in autumn.
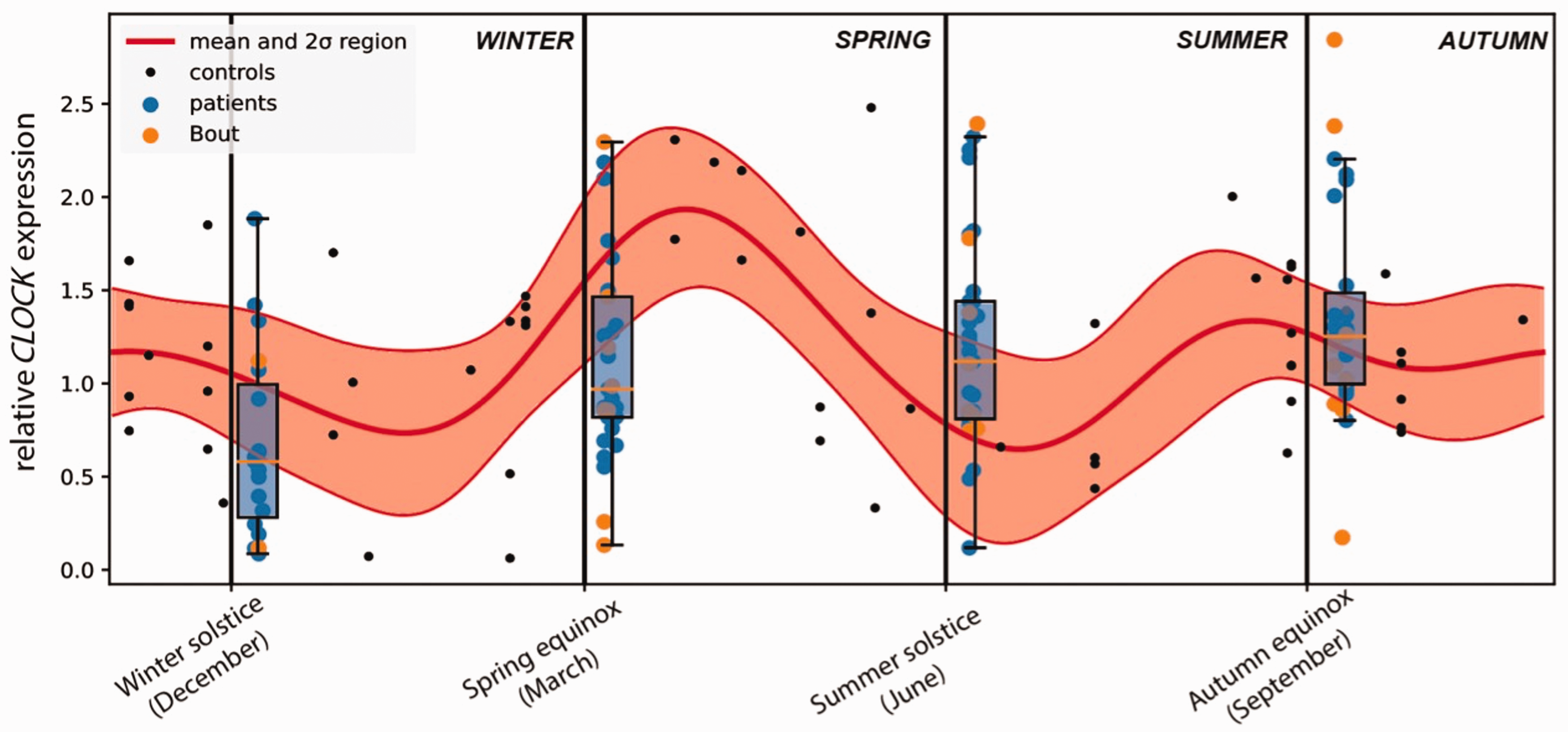
Circannual expression of CLOCK gene in controls and cluster headache patients.
CLOCK gene expression in CH patients
We collected 161 blood samples from CH patients in the week following each solstice and equinox in the northern hemisphere for a period of three years, with each patient being sampled two to four times over at least one year. The distribution of the samples per season was: 25 samples (15.5%) after winter solstices; 49 (30.4%) after spring equinoxes; 47 (29.2%) after summer solstices; and 40 (24.8%) after autumn equinoxes (Table 3). Twenty-four patients (48%) had four collections, 16 patients (32%) had three collections, whereas there were 10 patients (20%) with only one or two collections (Table 4). As for controls, CLOCK relative gene expression was determined by quantitative RT-PCR (detailed above).
Number of samples of cluster headache patients collected per season.

Number of sample collections for each cluster headache patient.
We then asked whether CLOCK expression of CH patients was different from the expression of the control population. For that, we compared CLOCK expression of the CH patients to the statistical model (Figure 1; box plots represent the CH patient population at the times of collections, with blue or yellow dots representing each patient sample). The average CLOCK expression of CH patients was either below the expression of the control population, in winter and spring, or above, in summer and autumn (Figure 1). We determined that CLOCK expression for CH patients is considerably different from that of the control population in winter (p-value mean = 0.006283), spring (p-value mean = 0.000006) and summer (p-value mean =0.000064), whereas no difference was detected in autumn (p-value mean = 0.262272) (Table 5). Our results suggest that expression of CLOCK gene in CH patients fluctuates less throughout the year than that of the control population.
Comparison of CLOCK gene expression between cluster headache patients and synthetic controls using a Gaussian process and p-values determined by Mann-Whitney tests. Adjusted p-values were determined by multiple test corrections using the Holm-Sidak method.
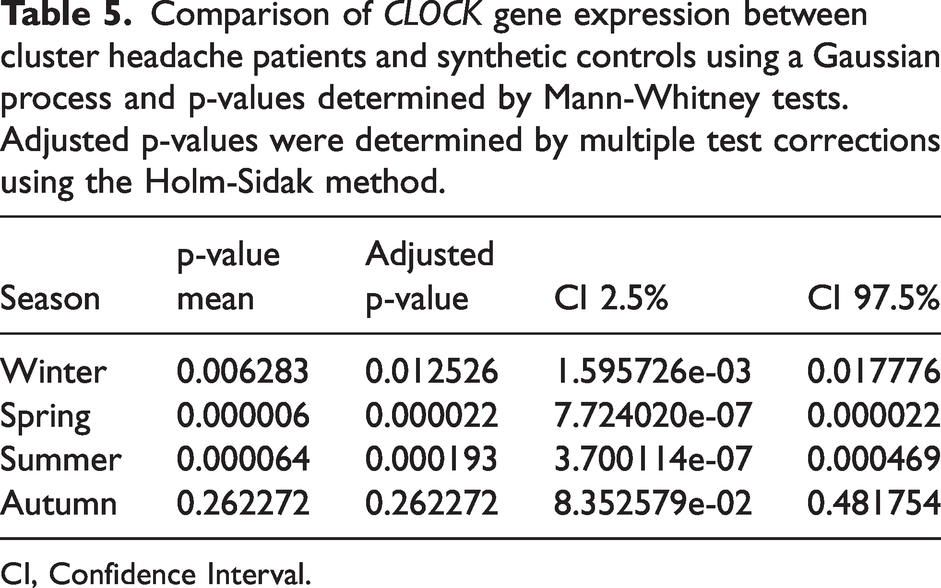
CI, Confidence Interval.
Variations in CLOCK expression across seasons for patients and controls
Additional tests were performed to assess whether CLOCK expression in CH patients changed less than in the control population. For each of the two groups, we determined the changes in expression occurring between two consecutive seasons. We observed that CLOCK expression in CH patients and controls tended to vary in the same direction, either both increasing or decreasing their expression from one season to the next (Figure 2). The only exception was the spring to summer transition, when CH patients had a very mild increase in CLOCK expression (slope = 0.017342, Online Supplementary Table 4), compared with an abrupt decrease in CLOCK expression for controls (slope = −0.97182, Online Supplementary Table 4). For each season transition, the variations in CLOCK expression were more pronounced in the control group than in the CH patient population, as assessed by the slopes of the curves linking consecutive seasons (Figure 2 and Online Supplementary Table 4). The exception observed was in the autumn to winter transition, where the patients’ changes in expression were slightly more noticeable (slope = −0.627580) than the changes in expression of controls (slope = −0.287782) (Figure 2 and Online Supplementary Table 4). In agreement, the up- or down-regulation of CLOCK gene expression between consecutive seasons was statistically significant for all season transitions in the control population but was only significantly different for the autumn to winter transition for CH patients (Figure 2 and Online Supplementary Table 4). Overall, CLOCK expression oscillated less from season to season for CH patients than for controls. These results further support the hypothesis that circannual changes in CLOCK expression are compromised in CH patients.
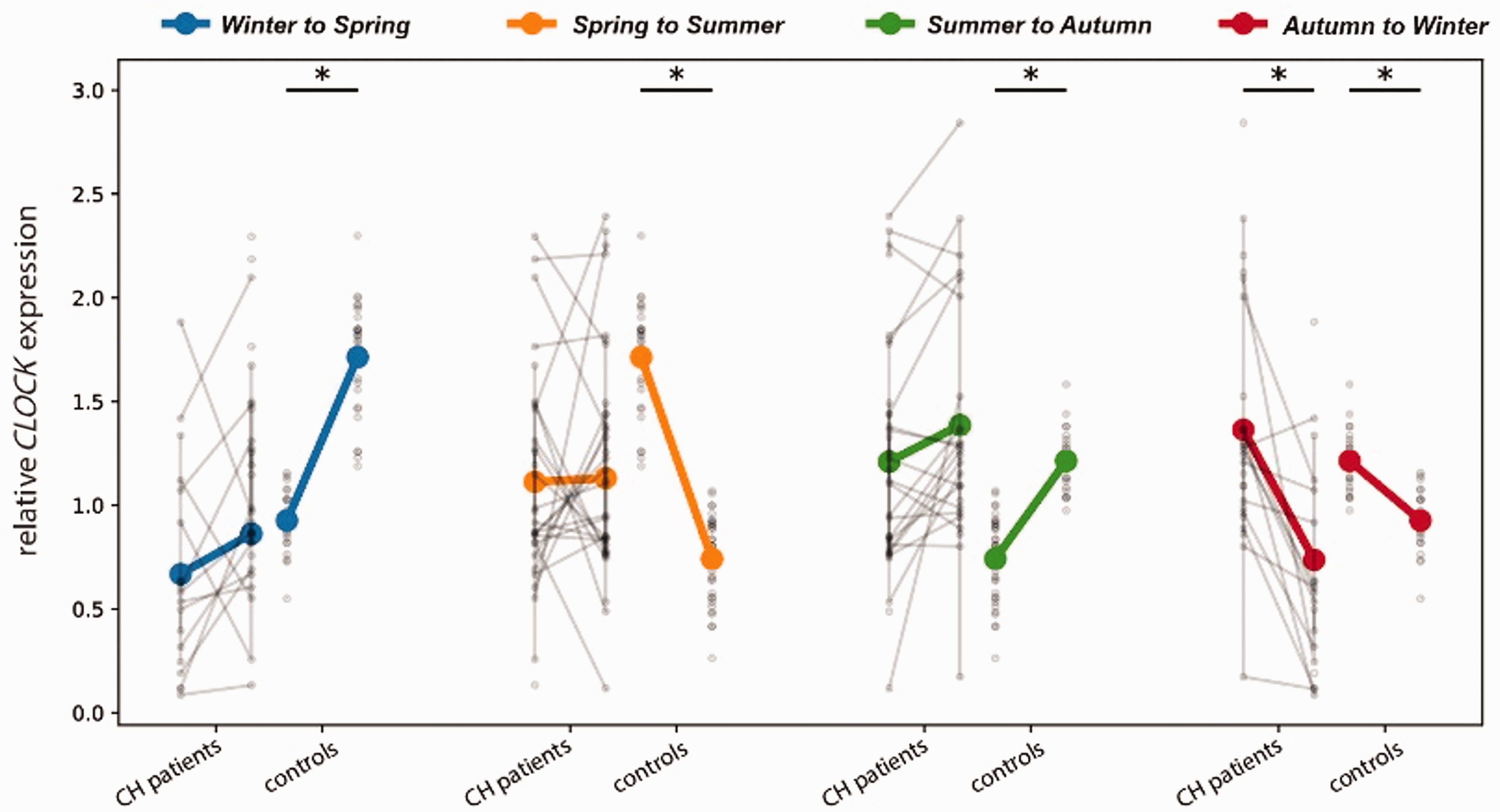
Changes in CLOCK expression across seasons for cluster headache patients and controls *p ≤ 0.05.
Effect of bouts, of being an episodic patient, and of having a circannual CH pattern on CLOCK expression
Of the 161 samples from CH patients, 36 samples (18.6%) were collected when the patients were in bout (Figure 1, yellow dots). We asked, for each season, whether being in a bout influenced CLOCK gene expression. Using multivariate regression analysis, we observed that bouts very modestly contributed to CLOCK gene expression only in spring (p-value =0.045755, adjusted p-value = 0.170838, Online Supplementary Table 5). There were no statistically significant effects due to bouts on CLOCK gene expression for the other seasons (Online Supplementary Table 5).
We also tested whether being an episodic patient contributed to CLOCK gene expression. Most of the CH patients in this study were episodic (90%), with only a small proportion of chronic patients (Table 2). Independently of the season, our results suggest no effects on CLOCK expression due to being an episodic patient (Online Supplementary Table 5). Finally, we asked whether having a circannual pattern of disease influenced CLOCK gene expression. In our cohort, the majority of the patients had a circannual pattern (70%, Table 2). For all seasons, our analysis showed that there are no effects on CLOCK expression due to the circannual pattern (Online Supplementary Table 5).
Discussion
In this study, we analyzed the circannual expression of the CLOCK gene among a population with cluster headache in comparison to a population of healthy controls. Our data suggests that expression of CLOCK in CH patients may fluctuate less throughout the year than that of the controls.
Several past studies have found a role for clock genes in specific brain functions including sleep, stress responses, learning and memory (7). Complex genetic factors are likely to play a role in CH etiology and pathophysiology (1), but no confirmed gene associations have so far been identified. Up to 22% of CH patients have a positive family history (31). The role of several gene polymorphisms was studied in CH patients, including hypocretin receptor 2 (HCRTR2), alcohol dehydrogenase 4 (ADH4), CLOCK, period circadian regulator 3 (PER3) and cryptochrome circadian regulator 1 (CRY1), yielding weak and occasionally conflicting results (4,16,32 –35). Insights from comparative genomic hybridization analysis of sporadic CH patients and controls identified a rearrangement in neurexin 3 (NRXN3) that was specifically found in some CH cases (36). In addition, genome-wide association studies (GWASs) pointed to genes potentially involved in CH susceptibility. The candidate genes identified have roles in pain processing, synaptic plasticity, neuroinflammation, immunological processes and circadian regulation (1,13,14,16). The most recent GWAS further identified genes associated with brain function, but also with arterial tissue function, as potentially causal for CH (15). The fact that distinct genes were identified in the different studies, and that no linkage to previous candidate genes involved in circadian rhythm has been found in the latest GWAS publication (15), may reflect the complexity of the genetic factors in CH, including the involvement of multiple genes and pathways, potential redundancies, and tissue-specific contributions. To strengthen the association between the candidate genes identified in genome-wide studies and the pathology of CH patients, further studies should test expression of those genes in well-characterized cohorts, in a similar way to what our study did with CLOCK gene expression, preferentially also performing subgroup analysis, as candidates may prove to be more relevant in certain CH patient subtypes. In addition, especially when testing circadian and circannual targets, the transversal collection of samples should be considered, so that the studies inform about variations along the day or across the year.
A series of potential markers of the time of the year have been identified, namely thyroid hormones and melatonin (37,38). Whereas CH patients do not appear to have altered levels of thyroid hormones (39), a possible link between CH and melatonin has been suggested (40). The authors reported lower melatonin levels in CH participants, which might indicate a possible dysregulation of circannual patterns in CH patients. Considering the periodical occurrence of CH attacks, clock genes are particularly interesting candidates for a role in this condition. Most studies have focused on single nucleotide polymorphism (SNP) and compared the allele or genotype frequency of mutations in CH patients versus control populations. One of the first studies to evaluate the occurrence of a particular allele or genotype of CLOCK in a population with CH was from Rainero et al. (41) and tested for 3092 T–>C CLOCK polymorphism (also known as 3111 T–>C or SNP rs1801260). In an association study in a cohort of Italian CH patients, rs1801260 has been linked to modifying diurnal preference (42). The main finding was that this polymorphism was similarly distributed in CH patients and controls. Further studies reported similar conclusions, not supporting the hypothesis that the T3111C CLOCK polymorphism is associated with CH (32,34,36,41). More recently, a Swedish study screened for three CLOCK SNPs, including rs1801260, and tested for association between each of the polymorphisms and CH. The authors found a significant association between the CLOCK gene variant rs12649507 and CH, with the less common A-allele of this SNP more frequent in CH patients (3). The authors further discuss that individuals with the rs12649507 AA genotype had shorter sleep duration compared to individuals with the GG or GA genotype, highlighting the association between sleep and CH. In addition, this study also showed that rs12649507 leads to increased CLOCK mRNA gene expression in human primary fibroblasts which points to a possible association between the AA genotype and the levels of CLOCK expression in CH patients.
Our study has several important strengths, including a large cohort of patients for a disease with an estimated prevalence below 0.1% of the general population. Importantly, this is a prospective study, with multiple collections in critical time points along the year when bouts are known to occur preferentially. Samples were obtained during and outside bouts. Patients were followed up for a long period and were well-matched to a control population that was used to establish a normal pattern of CLOCK gene expression during the year. The power and significance of the study may, however, be limited by several factors, including that: 1) the cohort size may underestimate some of the effects in gene expression differences; 2) no data on CLOCK gene expression is available to test for possible circadian differences because samples were only collected once (in the same two-hour window) daily; 3) samples for all four seasons were only collected for less than half of the CH patients (48%); 4) this is a single-center study; 5) CLOCK gene expression can vary in different cell types and different tissues, and PBMCs may not reflect CLOCK gene expression changes that may occur in the hypothalamus or other brain areas thought to be important for CH, and finally; 6) the CH patient population was heterogeneous, including subgroups that may prove to be distinct concerning CLOCK gene expression when larger cohorts are tested. We acknowledge that, in the subgroup analyses, the small numbers of patients in some of the subgroups may have masked any possible effects in CLOCK gene expression variations. Therefore, the findings are related to the population under study and may not be applicable to other populations.
In conclusion, in this study we found that CLOCK expression fluctuates substantially less in CH patients throughout the year as compared to matched controls, potentially reflecting a lower adjustability to circannual environmental factors in CH patients.
Clinical implications
Individuals with cluster headache (CH) show reduced seasonal variation in peripheral CLOCK gene expression compared to controls. This may contribute to the circannual clustering of headache bouts seen in CH patients because it may compromise their ability to adapt to circannual rhythm changes and environmental variation.
Supplemental Material
sj-pdf-1-cep-10.1177_03331024241247845 - Supplemental material for CLOCK gene circannual expression in cluster headache
Supplemental material, sj-pdf-1-cep-10.1177_03331024241247845 for CLOCK gene circannual expression in cluster headache by Renato Oliveira, Ana Neves-Costa, Dora Pedroso, Tiago Paixão, André B. Barros, Luís F. Moita and Raquel Gil-Gouveia in Cephalalgia
Footnotes
Acknowledgements
Dr. Ruth Geraldes, Dr. Raquel Gil-Gouveia, and Dr. Luis Filipe Moita conceived the original study concept, with Dr. Luis Filipe Moita securing project funding.
Dr. Isabel Pavão Martins and Dr. Elsa Parreira played a pivotal role in conceptualizing and drafting the initial study protocol, alongside Dr. Raquel Gil-Gouveia and Dr. Luis Filipe Moita.
Dr. Isabel Pavão Martins additionally recruited and included patients from the Hospital da Santa Maria Headache Clinic, and Dr. Inês Brás Marques assisted with the initial setup of the study at Hospital da Luz Lisboa.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Fundação Calouste Gulbenkian.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.