
Abstract
Background: After viewing dynamic noise surrounding a homogeneous grey patch (artificial scotoma), observers perceive a prolonged twinkling-noise after-image within the unstimulated area. It has been suggested that noise-stimulated neurons induce a long-range inhibition in neurons within the artificial scotoma, which generates a rebound signal perceived as twinkling noise following noise termination. We used this paradigm to test whether migraineurs have enhanced excitability or weakened inhibition.
Methods: Twinkling-noise duration was measured in 13 headache-free volunteers, 13 migraineurs with aura and 13 migraineurs without aura.
Results: The durations of the after-image were significantly shorter for both migraine groups compared to controls.
Discussion: Enhanced excitation of noise-activated neurons in migraineurs would produce stronger rebound activity and longer after-image durations, while weakened inhibitory mechanisms would diminish the rebound activity and shorten the after-image durations compared to control subjects. The results suggest that cortical inhibitory mechanisms might be impaired in migraineurs with and without aura.
Background
A general characteristic of migraine is hypersensitivity to visual stimuli, which may be due to augmented excitability or reduced inhibition of migraine visual cortex (for review see (1)). Recently, we revealed substantial external noise-exclusion deficits in migraine-with-aura (MA) and a minor impairment of noise exclusion in migraine-without-aura (MO) (2). Both migraine groups had higher contrast thresholds for detecting a small luminance disk in the presence of high luminance noise levels, with performance of MA subjects significantly poorer than that of control subjects. Equivalent input noise analysis indicated that the internal (neuronal) noise, induced by the external noise, was significantly higher in both migraine groups compared to control subjects. This finding suggests that migraineurs may experience an abnormally high variability in the neuronal activity involved in the processing of a luminance stimulus, perhaps due to reduced neuronal suppression within the visual cortex in migraineurs with aura.
These findings led us to question whether migraineurs may also demonstrate deficits when stimulated within a large noisy environment. Ramachandran and Gregory (3) have described a non-invasive paradigm that allows the investigation of long-range interactions between neurons within the visual system. If observers view a stimulus consisting of a homogeneous grey patch (artificial scotoma), surrounded by dynamic luminance noise, the central artificial scotoma appears to be filled in by dynamic noise from the surrounding field. When the dynamic noise is switched off, observers perceive a prolonged twinkling after-image – reminiscent of the twinkling ‘snow’ seen on an untuned analogue television – in the region of the artificial scotoma only (as seen in Figure 1A, B).
(A) Illustration of the artificial scotoma stimulus used in this experiment. Subjects viewed a homogeneous grey field (circle) surrounded by dynamic binary noise, for 7 s on each trial. (B) Appearance of the twinkling after-image seen by subjects following the cessation of the artificial scotoma stimulus. (C) Model predictions for the strength of hypothetical twinkling after-images as a function of time after the termination of the artificial scotoma stimulus. Lines show the temporal course of the twinkling after-image strength assuming enhanced (dashed line), normal (solid thick line) and reduced (solid thin line) long-range inhibition. The horizontal (dotted) line illustrates a threshold determining the cessation of the perception of the twinkling after-image (denoted by the arrows). Enhanced long-range inhibition would be predicted to give rise to increased twinkling after-effect strength and longer after-image duration. Reduced long-range inhibition would give rise to reduced after-effect strength and shorter after-image duration.
A possible mechanism underlying the perception of twinkling after-images, arising within the unstimulated area, could be post-inhibitory rebound activity in the cortical representation of the artificial scotoma after the cessation of the surrounding noise (4). The stimulated visual neurons are depolarized during adaptation to the surround noise. This induces a long-range inhibition (hyperpolarization) in neurons, within the unstimulated area of the artificial scotoma. This hyperpolarization is released after the dynamic noise is terminated, generating a rebound signal that may be perceived as twinkling noise.
Hypersensitivity to external visual stimuli in migraine could be due to either enhanced excitability or weakened inhibitory mechanisms (1). Which of these mechanisms is impaired in migraine is still under debate. Studies, using backward masking, contrast gain control and orientation discrimination thresholds have tested the hypothesis that inhibition in migraine is impaired, but the findings were mixed (1).
This study used the paradigm of an artificial scotoma. This paradigm offers a way to distinguish between effects related to enhanced excitability and those associated with weakened cortical inhibition in migraine. The duration of the twinkling noise after-image (Figure 1B), produced by adaptation to dynamic noise surrounding an artificial scotoma (Figure 1A), in migraineurs and headache-free subjects was measured. Figure 1C illustrates a hypothetical time-course of the after-image strength in headache-free subjects with normal long-range inhibition (thick solid line). According to the rebound hypothesis, the duration of the twinkling after-image would be determined by a given threshold level of the rebound activity (Figure 1C, horizontal dotted line) below which the perceived after-image disappears. We hypothesized that enhanced excitation and weakened inhibition of neurons, activated by the noisy surround, may lead to opposite effects on the after-image durations. Elevated long-range inhibition in migraineurs, resulting from enhanced excitability of the activated neurons, would result in stronger post-inhibitory rebound activity and consequently longer after-image durations than in control subjects (Figure 1C, dashed line). On the other hand, impaired long-range inhibition in migraineurs, due to weakened inhibition of the activated neurons, would produce weaker post-inhibitory rebound activity and hence shorter after-image durations than in control subjects (Figure 1C, solid thin line).
Methods
Participants
Thirteen headache-free volunteers (mean age ± SD: 30 ± 4.7 years) were recruited from among the staff and students at Glasgow Caledonian University, Scotland. These volunteers reported no migraine headaches ever, verified by a structured questionnaire in accordance with IHS-II classification (5). Thirteen MA (30 ± 5.2 years) and thirteen MO subjects (28 ± 7.8 years) were recruited from patients attending the Headache Clinic at the Neurology Department, University Clinic Jena, Germany, where they had been previously diagnosed by a consultant neurologist. Volunteers using preventative medication or reporting a migraine episode 3 days prior or following testing were excluded. The median frequency of migraine episodes in the month preceding the test was one for the MA group (range 1–2); all subjects in the MO group reported only one episode. The median headache duration was 10 years (range 1–32) for the MA group and 5 years for the MO group (range 2–16 years); this difference was not significant (Mann–Whitney U-test, z = −0.697; p = 0.486). The median MIDAS score was 17 (range 3–53) and 16 (range 0–30) for the MO and MA groups, respectively. These did not differ significantly between these groups (Mann–Whitney U-test, z = −1.540; p = 0.124). Subjects had normal visual acuity (>20/30), contrast thresholds (<1%), IOP (<21 mmHg) and visual fields (assessed by automated perimetry). Ethics committees of Glasgow Caledonian University and University Clinic Jena approved the study. The research adhered to the tenets of the Declaration of Helsinki.
Apparatus
Stimuli were generated by a PC running custom-written software and presented on a 19″ high-resolution RGB Iiyama Vision Master Pro 450 (Iiyama, Tokyo, Japan). The mean luminance of the stimulation field was 27.9 ± 0.07 cd/m2 and the mean ambient luminance of the test room was 5.1 ± 3.5 cd/m2.
Stimuli
The stimulus display contained dynamic binary noise samples of 90% contrast and noise-pixel size of 4.4 arcmin. The artificial scotoma was created by a circular grey patch of 4 deg in diameter surrounded by dynamic noise (see Figure 1A). The viewing distance was 85 cm.
Procedure
Participants were instructed to fixate on a dot in the centre of a circular grey patch surrounded by dynamic noise. Subjects adapted to the artificial scotoma stimulus for 7 s, after which a uniform screen of the same mean luminance was immediately presented. This leads to perception of a strong twinkling after-image within the area of the artificial scotoma, whose strength gradually reduces. Subjects were instructed to press a button when the twinkling after-image disappeared. Data for 25 trials were collected for each subject.
Statistical analysis
Prior to group analyses, individual data were tested for normality (Shapiro–Wilk test). The distributions of the after-image durations for 12 out of 39 subjects were not normal (Shapiro–Wilk test, p < 0.05), and so we used the median of the after-image durations for each subject as a measure of central tendency of the individual data. The data distribution for each group was also tested for normality. The distributions of the data for each migraine group did not significantly depart from a normal distribution (Shapiro–Wilk test, p = 0.08), while the data for the control group did not follow a normal distribution (Shapiro–Wilk test, p = 0.017). Using ± 3 SD from the mean as a criterion, a single participant in the control group was classified as an outlier (Figure 2, cross) and was omitted from the data analysis. Excluding this individual, the control group data had a normal distribution (Shapiro–Wilk test, p = 0.92). The group means were compared using independent sample t-tests.
Mean twinkling after-image durations for headache-free (filled circle) and migraine (filled square) subjects. Open circles represent individual data for headache-free subjects; open triangles show individual data for MA subjects; filled triangles denote individual data for MO subjects. * Represents significant significance at p < 0.05. The cross shows the after-image duration of a control subject (outlier) whose result was larger than 3 SD from the mean and was excluded from the data analysis. Error bars illustrate 95% confidence intervals.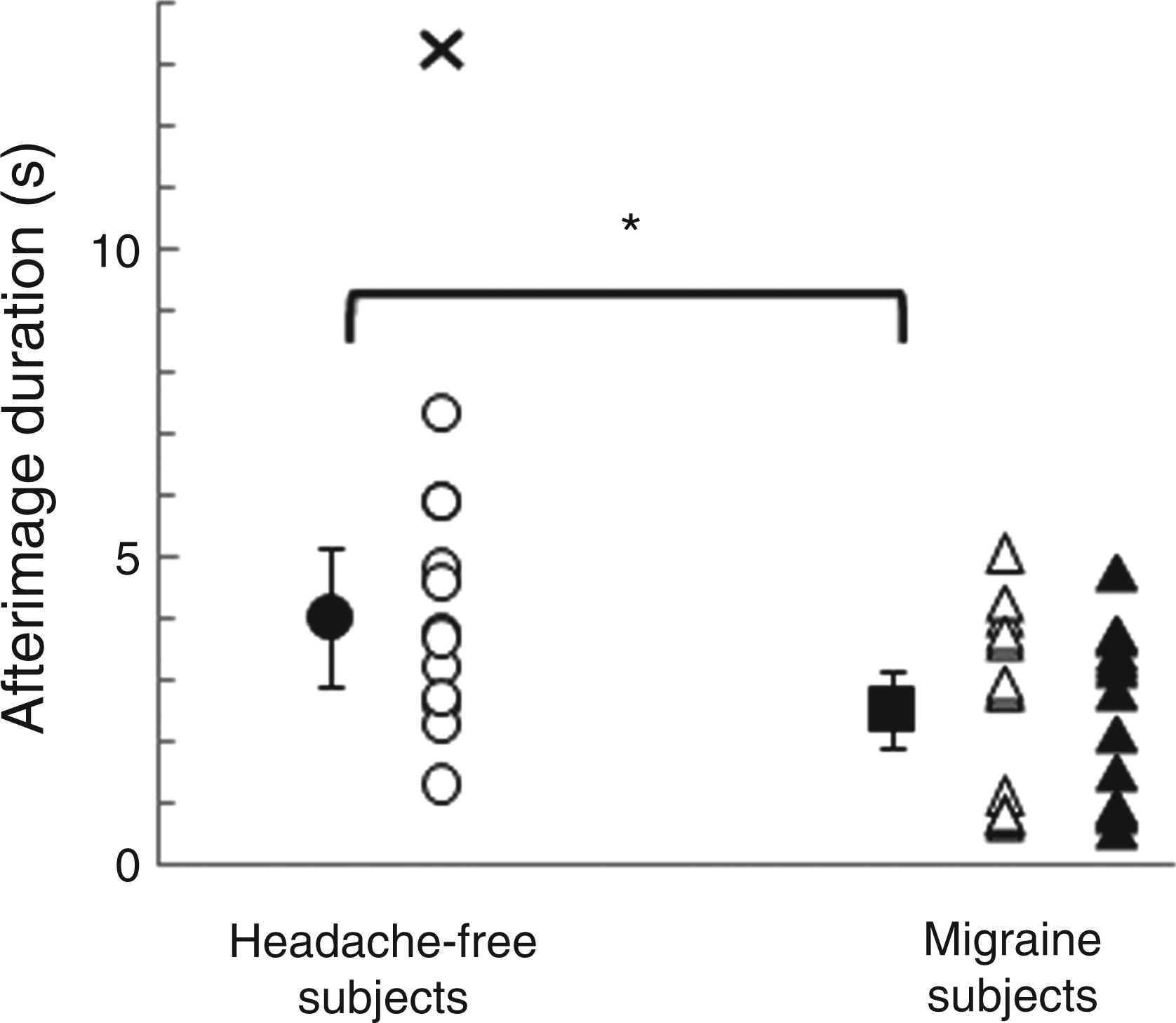
Results
The mean after-image durations of the two migraine groups (MO: mean ± 95% CI, 2.57 ± 0.93 s; MA: 2.47 ± 0.83 s) did not differ significantly (t24 = 0.19, p = 0.85); hence all migraine data were pooled for comparison with headache-free subjects. The effect size, describing the ratio of the difference between the data for both migraine groups and the pooled standard deviation was very small (d = 0.07). Post hoc power analysis indicates that for a power of 0.8, a very large sample (n > 2000 in each migraine group) would be required to detect a significant difference between the migraine groups (6).
The pooled migraine groups showed significantly (t36 = −2.82, p = 0.008) shorter after-image durations (2.52 ± 0.62 s) than those of headache-free subjects (4.01 ± 1.11 s). After-image durations and frequency of migraine events during the last month before testing showed no significant correlation (r24 = 0.164, p = 0.424). There was no significant correlation between after-image duration and the time since the onset of the migraine problem (r24 = 0.178, p = 0.384).
Discussion
This study found the duration of the twinkling-noise after-image, perceived within the area of an artificial scotoma following adaptation to surround dynamic luminance noise, to be significantly shorter in migraineurs compared with control subjects. This finding is in line with the notion of impaired long-range inhibitory mechanisms producing weakened post-inhibitory rebound activity within the neural representation of the artificial scotoma in migraine. The results do not support the hypothesis for neuronal hyperexcitability in migraine (7), which would be predicted to result in stronger rebound activity and hence, longer twinkling after-image durations in migraineurs than in headache-free subjects (8).
Impaired cortical inhibition in migraine could arise as a consequence of hypoxia induced by aura (9). Therefore, one might expect after-image durations in MA subjects would be significantly shorter than in MO subjects. However, no significant difference between the after-image durations for the two migraine groups was found. This could be due to the relatively small numbers tested. Increasing the statistical power by increasing the sample size does not seem promising, given the very small effect size (d = 0.07) and the very large sample size in each migraine group (N > 2000), which would be required to approach a significant difference. Another reason could be related to the mean age of the migraine participants, which was relatively young. Testing migraineurs of older age with longer duration of migraine history could potentially lead to statistically significant differences between after-image durations in people with MA and MO, however among the subjects tested there was no correlation between the onset of the migraine problem and the duration of the after-image. Verifying the hypothesis of specific impairment of cortical inhibition in MA is complicated, however, because of considerable overlap between MA and MO (10). In one study, 70% of the participants had experienced episodes of both migraine with and without aura episodes (11).
Migraine-related impairments of cortical inhibition could be widespread within the visual system (1). In a previous study (2), we found that migraineurs had higher levels of multiplicative internal noise when detecting a luminance disk embedded in luminance noise. A recent study (10) replicated our experiment and found similar noise-exclusion deficits in migraine. These noise-exclusion deficits in migraine could be related to the processing of localized stimulus features in early cortical areas (V1). Impaired ability to exclude visual noise in a pattern detection task could be due to untuned perceptual templates. The tuning characteristics of visual neurons are modulated by cortical suppression at the neuronal level, for example GABA-mediated suppression (12). Impairments of cortical inhibition could also result in increased spontaneous activity, as shown by a study applying GABAa receptor antagonists in the medial temporal cortex (MT) neurons (13). Increased additive noise in migraine, which could be related to increased spontaneous activity, was found in a radial-frequency discrimination task involving a higher cortical area (V4) of global contour coding (10). Another study found increased thresholds for detecting coherently moving dots embedded in motion noise, suggesting an increase in baseline neuronal noise at higher motion processing stages (MT) (14).
The twinkling after-image phenomenon could be cortical in origin or inherited from earlier visual processing stages. Tyler and Hardage (8) have suggested that the twinkling after-image may involve principally the magno-cellular visual pathway, given that the after-effect is not elicited by equi-luminant chromatic dynamic noise. While the broad orientation tuning of the twinkling after-effect has been taken as evidence for a pre-cortical origin locus for this perceptual phenomenon (15), Reich and colleagues found the perceived size of the twinkling after-effect to be smaller than the size of the inducing artificial scotoma following binocular adaptation (16). Dichoptic adaptation caused a reduction of the twinkling after-effect that was similar to that found after binocular adaptation, suggesting instead a cortical origin for the after-effect.
The reduced after-image durations in migraine could also be accompanied by a reduced strength of the after-image activity. The hypothesis that weakened inhibition leads to a reduced duration and strength of the after-image activity could be verified in vivo in animals using GABAa receptor antagonists (12,13).
In summary, this study investigated for the first time long-range interactions within the visual system of migraineurs. Migraine subjects showed significantly shortened twinkling after-image within the area of an artificial scotoma surrounded by dynamic luminance noise. This finding is suggestive of weakened inhibitory mechanisms within the early visual stages.
Footnotes
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Conflict of interest
None declared.