
Abstract
Objectives: Migraine pathophysiology involves several pathways. Our aims were to explore a possible role of the brain-derived neurotrophic factor gene (BDNF) in migraine susceptibility; to study, for the first time, the calcitonin gene-related peptide gene (CGRP); and a possible interaction between the two.
Methods: Using a case-control approach, four tagging single nucleotide polymorphisms (SNPs) (rs7124442, rs6265, rs11030107, and rs2049046) of BDNF and one tagging SNP—rs1553005—of CGRP were analyzed in 188 cases and 287 controls. A multivariable logistic regression was performed, adjusting for gender. Allelic and haplotypic frequencies were estimated. Interaction was assessed by a stepwise multivariable-logistic regression and confirmed by a multifactor dimensionality reduction analysis.
Results: No significant main effects were found; however, a significant interaction was found between BDNF and CGRP, showing an increased risk for the AT-genotype of rs2049046 and the GC-genotype of rs1553005 (odds ratio = 1.88, 95% confidence interval: 1.20–2.93) for migraineurs.
Conclusion: Our data support the hypothesis of an interaction between BDNF and CGRP in migraine susceptibility that should be further explored.
Introduction
Migraine is a common multifactorial disease (1,2). Several different processes are involved in its pathophysiology, such as alteration of pain and sensory input, increased sensitivity of the cortex leading to aura, central pain facilitation, neurogenic inflammation and brain stem nociceptor sensitization (3,4).
BDNF (brain-derived neurotrophic factor) is important for neuronal growth, development and survival in the central nervous system, and is the most abundant neurotrophin in the brain (5,6). BDNF is expressed in the nociceptive sensory neurons, acting as a central pain modulator at both the spinal and supra-spinal level, contributing to central sensitization and modulating the activation of glutamatergic, N-methyl-d-aspartate (NMDA) receptors (7–9). A significant decrease in the levels of BDNF in platelets was found in migraineurs, showing that it may also have a role in migraine pathophysiology (8). One of the mechanisms involved in migraine is central sensitization, characterized by activity-dependent plastic changes in second-order trigeminal neurons (10). BDNF has been pointed out as a mediator of the trigeminal nociceptive plasticity (9,11). A previous study evaluated several genes that can be expressed after cortical spreading depression (CSD), the mechanism hypothesized to underlie migraine pathophysiology, especially migraine with aura (MA) (10,12). BDNF was one of the genes found to be differentially expressed after CSD (12).
Several studies showed that neurogenic inflammation is associated with pathogenesis of migraine (13). When neuronal excitability occurs, leading to primary brain dysfunction, the perivascular trigeminal sensory nerve fibers are activated, resulting in the release of vasoactive peptides, such as substance P and calcitonin-gene related peptide (CGRP), from trigeminal fibers (13). CGRP is the most abundant neuropeptide in perivascular sensory trigeminal nerve fibers and one of the mediators of neurogenic inflammation; it is a potent vasodilator (14,15), enhancing the activation of second-order neurons, thus contributing to pain transmission (13,16). CGRP has an active role in the activation and sensitization of nociceptors at the peripheral and central level (17), modulating nociceptive transmission in the trigeminovascular system (15).
The involvement of CGRP in migraine pathophysiology has been supported by the finding that the serum levels of CGRP are increased during migraine attacks and there is a decrease after treatment with triptans, although this was not observed in a more recent study (18–21).
CGRP is co-expressed with BDNF in trigeminal ganglion neurons in rats. In addition, CGRP enhances BDNF release from trigeminal neurons in vitro; this is independent of extracellular calcium and is abolished by a CGRP-receptor antagonist (11).
A case-control study found no evidence of a BDNF single nucleotide polymorphism (SNP) (rs6265) being associated with migraine susceptibility (22). The aims of this study were (i) to further explore a possible role of BDNF in migraine susceptibility and (ii) to study, for the first time, CGRP as a candidate gene. Furthermore, since both genes are co-expressed and involved in migraine pathophysiology (3), we also assessed their possible interaction in migraine’s susceptibility.
Subjects and methods
Subjects
A sample of 188 unrelated migraine patients from the neurology clinic, at (Hospital de Santo António) HSA, Porto, was sequentially enrolled in this study. Patients with familial hemiplegic migraine were excluded. Control subjects (287), with no personal history of migraine, were ascertained among healthy blood donors and from the Obstetrics and Gynecology Department of HSA. Controls were from the same ethnic and geographical origin (north of Portugal) as cases, and were age-matched to these. A diagnostic interview was performed both in cases and controls, based on the operational criteria of the International Headache Society (IHS), using the same structured questionnaire. The first edition of these criteria (ICHD-I) (23) was used before 2004; revising the diagnosis using the second edition (ICHD-II) (24) showed no differences in patients’ diagnosis (data not shown). Participants gave their written informed consent and the project was approved by the Ethics Committee of HSA.
Genotyping
Genomic DNA was extracted from peripheral blood leucocytes, using a standard salting-out method (25); or from saliva using ORAGENE kits and DNA extraction according to the manufacturer’s instructions (DNA Genotek, Kanata, Ontario, Canada).
SNPs were selected based on a data dump from the International HapMap Project (Release 24, November 2008, on NCBI B36 assembly, dbSNP build 126) (www.hapmap.org) and tagging SNPs were selected using Haploview 4.1 (Broad Institute, Cambridge, MA, USA) (26), capturing all the variation for both genes studied at an r2 threshold of 0.80 and with a minor allele frequency (MAF) of 0.10, by an aggressive tagging approach. For BDNF (NM_170732), these included rs712442 (located on chromosome 11: 27,634,117; on 3′ UTR), rs6265 (27,636,992; a functional SNP: p.V66M), rs11030107 (27,650,911; on intron 1) and rs2049046 (27,679,851, according to the Ensembl database; on 3′ gene region). Two of the SNPs were combined as a two-marker haplotype, allowing coverage of a broader region of SNPs tagged. For CGRP (NM_001033953), rs1553005 (located in chromosome 11: 14,950,566; on 3′ gene region) was selected.
Allelic discrimination was performed using Real-Time PCR (iQ5 Real-Time PCR Detection System, Bio-Rad Laboratories). Primers and molecular beacons were designed using Beacon Designer 6.0.
For BDNF, Real-Time PCR reactions were performed using 20 ng/µL DNA, 10 µL of iQTM Supermix or iQTM Multiplex Powermix, 1.2-8.0 mM MgCl2 and 0.5 µM of each primer, in 20 µL final reaction volume. Concentration of the probes used was 0.06 µM of probe 1 (labeled with FAM) and 0.25 µM of probe 2 (labeled with Cy3), for rs6265; 0.04 µM of probe 1 (labeled with FAM) and 0.16 µM of probe 2 (labeled with Cy3), for rs2049046; 0.03 µM of probe 1 (labeled with FAM) and 0.15 µM of probe 2 (labeled with Cy3), for rs7124442; 0.03 µM of probe 1 (labeled with FAM) and 0.30 µM of probe 2 (labeled with Cy3), for rs11030107. For the rs1553005 SNP of CGRP, Real-Time PCR reactions were performed using 20 ng/µL DNA, 10 µL of iQTM Supermix, 8 mM MgCl2, 0.5 µM of each primer, 0.03 µM of probe 1 (labeled with FAM) and 0.15 µM of probe 2 (labeled with Texas Red), in 20 µL final volume.
Primer and probe sequences are available upon request. Cycling conditions used were: an initial denaturing at 95°C, for 3 minutes and 30 seconds, followed by 35 cycles of 30 seconds, at 95°C, 60 seconds at 60°C (for annealing, and at this point fluorescence was measured), and 30 seconds at 72°C. Genotypes were determined automatically by the signal processing algorithms of the iQ5 Optical System Software, version 2.0 (Bio-Rad Laboratories); results were compared with the genotypes visually scored, based on the fluorescent emission data depicted (relative fluorescent units [RFUs]). To validate the Real-Time PCR reaction, we included three individuals previously genotyped by sequencing (a heterozygote, a homozygote for variant 1 and a homozygote for variant 2). If genotypes were uncertain, they were additionally sequenced, using the Big Dye Terminator Cycle Sequencing v1.1 Ready Reaction (Applied Biosystems), according to the manufacturer’s instructions, and an ABI-PRISM 3130 XL genetic analyzer (Applied Biosystems).
Statistical analysis
The Genetic Power Calculator (http://pngu.mgh.harvard.edu/∼purcell/gpc/) was used, assuming a high-risk allele frequency of 0.1, a relative risk for a homozygous genotype of 2.25 and 1.5 in heterozygosity. Hardy-Weinberg equilibrium was tested, using HWE software (http://linkage.rockefeller.edu/ott/linkutil.htm). Demographic data of patients and controls were compared using a chi-square test for qualitative variables. To compare allele frequencies between cases and controls, a chi-square test was used and odds ratios (OR) were estimated, with 95% confidence intervals (CIs). Our initial significance level was set at α = 0.01.
A multivariable-logistic regression was performed (with the most frequent homozygote as the reference), to evaluate association between SNPs and migraine, including the four SNPs of BDNF and the CGRP SNP in the model, and adjusting for gender. We assessed interaction between BDNF and CGRP by a stepwise multivariable-logistic regression. For the logistic regression analyses, significance was set to α = 0.005, using a Bonferroni correction for multiple comparisons. These analyses were performed using SPSS (version 16.0 for Windows).
Gene-gene interactions were also evaluated using multifactor dimensionality reduction (MDR) software (version 2.0) (27,28). MDR is a non-parametric and genetic model-free approach that can identify combination of SNPs involved in disease susceptibility (27). We performed a MDR analysis of the rs2049046*rs1553005 interaction using a forced approach. By using this forced analysis, we evaluated a specific set of attributes and we obtained an unbiased estimate of the testing accuracy. We used a tenfold cross-validation to avoid false-positives (29). The significant results obtained were corrected for multiple testing using the permutation test implemented on the MDR Permutation Tool (version 1.0) (30).
Haplotype frequencies were compared between cases and controls, for the BDNF gene, using Haploview 4.1 (26) with all parameters set at the default values. Frequencies of haplotypes analysed were above 1%, according to the Haploview threshold; to correct for multiple comparisons, when estimating allelic and haplotype frequencies, 10,000 permutations were used.
Results
Demographic and clinical data of migraine patients and controls

SD = standard deviation.
A migraine prevalence of 16% has been estimated in a Portuguese population (31,32). We found that we had a power of 64% to detect an association with our sample (for a nominal significance level of 0.05).
Both case and control groups were in Hardy-Weinberg (H-W) equilibrium for the four SNPs of the BDNF gene (rs712442, rs6265, rs11030107 and rs2049046) and for rs1553005 of CGRP. The correlation between the BDNF SNPs was small, denoting the weak linkage disequilibrium (LD) between them; in our sample, these SNPs were also in weak LD (data not shown). To confirm Real-Time PCR efficiency genotyping we have sequenced a random sample. For BDNF SNPs rs6265, rs2049046 and rs712442 all genotypes were concordant and thus 100% efficiency was considered. However, for rs11030107, some genotypes were incorrect; therefore, we sequenced all individuals for this SNP. Regarding CGRP, we also sequenced all individuals and we found 100% of identity with the previous results from Real-Time PCR.
Allele frequencies of SNPs studied in migraine patients and controls

SNP = single nucleotide polymorphism. BDNF = brain-derived neurotrophic factor (gene). CGRP = calcitonin gene-related peptide (gene). OR: odds ratio; CI: confidence interval.
Results from the multivariable logistic regression, gender-adjusted

(ref) = reference category. OR = odds ratio. CI = confidence interval.
Interaction results found between CGRP*BDNF, assessed by a stepwise logistic regression analysis
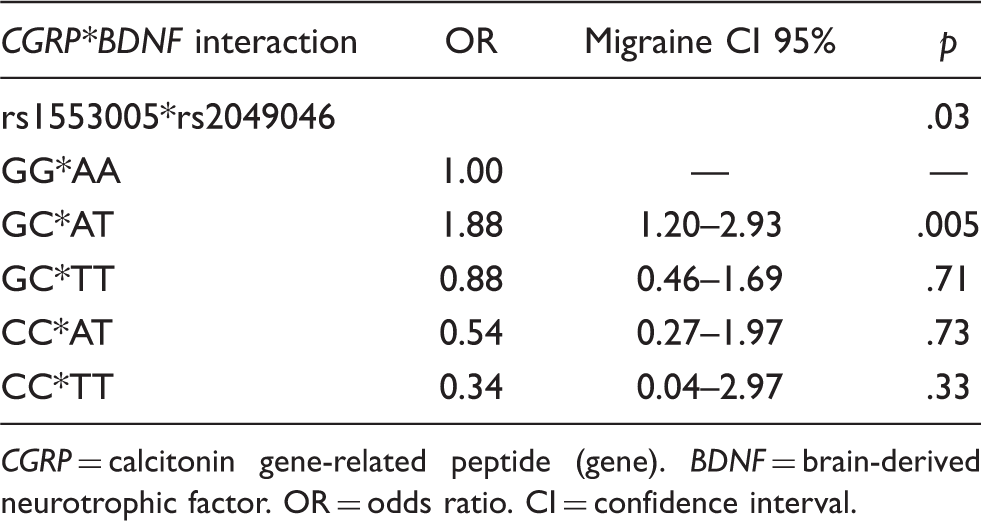
CGRP = calcitonin gene-related peptide (gene). BDNF = brain-derived neurotrophic factor. OR = odds ratio. CI = confidence interval.
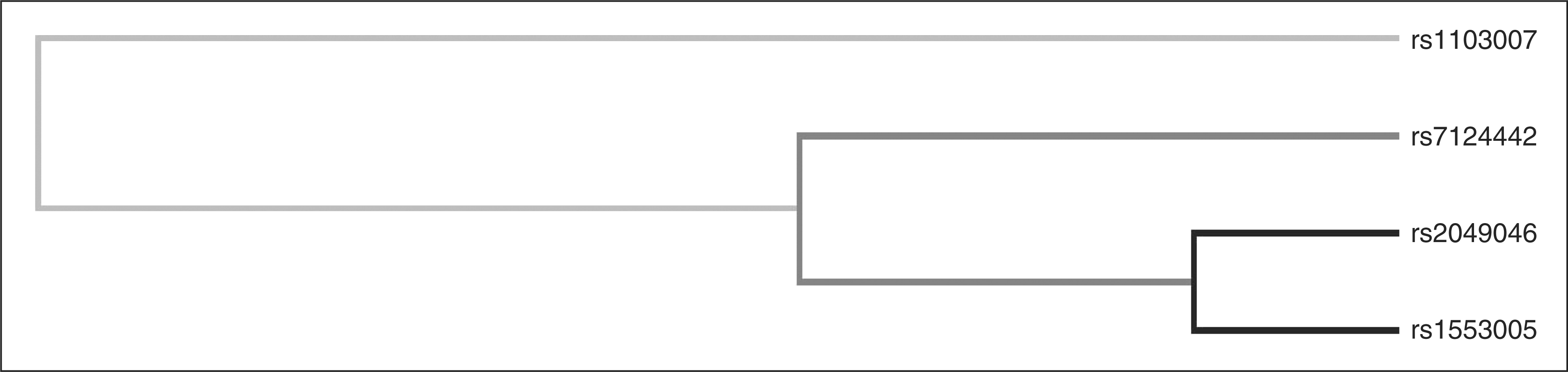
With the MDR analysis, we found that the best model for the interaction between rs2049046 and rs1553005 showed a testing balanced accuracy of 0.53 and a cross-validation consistency of 10/10 (Table 5). After permutation testing, this model was still significant (p = .002), which confirmed the interaction result found in the logistic regression analysis. Figure 1 shows the combinations associated with high and low risk. The varied patterns of high and low risk cells, with no linearity across each multifactor dimension, provide evidence of gene-gene interaction. The dendogram in Figure 2 shows a strong interaction between the two SNPs, which is indicative of a non-additive effect.
Interaction graphic for rs2049046 and rs1553005. High-risk combinations are depicted as dark-shaded cells, and low-risk combinations as light-shaded cells. For each cell the left bar indicates the cases and the right bar indicate the controls. Interaction dendogram for the SNPs genotyped. The dendogram clearly shows a strong interaction effect between rs2049046 and rs1553005 (darker lines suggest that there is a synergistic relationship: the shorter the lines, the stronger the interaction). SNPs = single nucleotide polymorphisms. Interaction results found between CGRP*BDNF, assessed by the MDR method
CGRP = calcitonin gene-related peptide (gene). BDNF = brain-derived neurotrophic factor. OR = odds ratio. CI = confidence interval. MDR = multifactor dimensionality reduction.

For BDNF, we also performed a haplotype-based analysis. No differences were found between cases and controls for any of the haplotypes evaluated (data not shown).
Discussion
The aim of this study was to assess the possible involvement of BDNF and CGRP in migraine susceptibility and their possible interaction. A previous study found no evidences of BDNF as a risk factor for migraine, regarding the functional SNP rs6265 (22). We now used a haplotype-block tagging approach, selecting SNPs that cover whole gene common variation. No significant differences were found between cases and controls regarding the SNPs analysed.
For CGRP, only one tagging SNP (rs1553005) was selected with the software used, covering also the whole variation within this gene, with the parameters specified for the European population. Several polymorphisms of CGRP are described as non-informative in dbSNP (http://www.ncbi.nlm.nih.gov/SNP) in European populations, although they may be informative for other populations. This may be a reason why these SNPs are not captured when searching for tagging SNPs.
For CGRP, no differences were also found between cases and controls. No differences were found regarding gender for BDNF and CGRP (data not shown).
We have also assessed a possible interaction between BDNF and CGRP. The Hierarchically Well-Formulated Rule (HWFR) is commonly used when a logistic regression is performed and assumes that lower order terms of the highest order interaction term are included in the model (33). With limited number of SNPs (as in our study), a stepwise logistic regression without HWFR may be a better approach to test for SNP-SNP interactions (33). Therefore, we performed a stepwise logistic regression, allowing that the final model included only the interaction terms between the BDNF and the CGRP SNPs, in an exploratory analysis of a putative interaction between the two genes. Further studies are now needed to confirm these results.
A significant interaction between rs2049046 and rs1553005 was found, showing an increased risk for the double heterozygotes AT and GC (OR = 1.88, 95% CI: 1.20–2.93) when compared with the most frequent double homozygotes (AA of rs2049046 and GG of rs1553005).
However, logistic regression parameters may not be completely reliable due to the limited number of subjects in each group (34).Therefore, we employed a MDR analysis. This method combines genotypes into a single dimension with two groups (high or low risk). MDR is also able to detect a high-order interaction in the absence of any statistically significant main effects. The combination of cross-validation and permutation testing minimizes the occurrence of false positive results (27). Furthermore, MDR is able to detect gene-gene interactions even in small sample sizes (35), which gives us confidence in our results. With the MDR analysis we also found a highly significant two-way interaction between those SNPs associated with an increased risk for migraine, confirming the results of the logistic regression analysis. Therefore, an interaction between these variants, or between variants in LD with them, may be involved in migraine susceptibility.
Our sample size is a limitation of our study and does not give us enough power to detect a variant with an OR < 1.5. Also, rare alleles may also be involved in the susceptibility of migraine (36), although we were interested in studying the role of common variants (MAF > 0.10). Therefore, we could not exclude that these genes may be independent risk factors for migraine, although this was not detected in our sample. However, some gene-gene interactions occur even without the presence of a significant main effect (37) and future association studies should take this into account.
Nonetheless, for common variation, we covered the whole gene region with the SNPs studied for both genes. Importantly, our high case:control ratio increases the power to detect any effects in our sample. In addition, cases and controls were matched for age-at-observation and gender, and were from the same population and geographic region; several studies using lineage markers, sensitive to population stratification, have showed there is no substructure within the Portuguese population (38–40). Also, we corrected for multiple testing, to prevent type-I errors, and used logistic regression analyses to incorporate all SNPs in the model and analyze their effects together. Additionally, we set our initial level of significance at p < .01, to use a stringent statistical criteria as suggested by Bird et al. (41). Furthermore, the MDR analysis strongly confirmed the interaction results found with the logistic regression analysis. The application of this method has already led to the identification of gene-gene interactions for several diseases, such as sporadic breast cancer, essential hypertension, schizophrenia and familial amyloid neuropathy (29).
Several findings support a role of CGRP in initiating and sustaining migraine episodes (42). CGRP has been extensively studied regarding its possible role in migraine treatment. Antagonists for CGRP receptors may act at the central level and modulate nociceptive trigeminovascular transmission in the cat, and are arising as a new generation of migraine drugs (42–44). Furthermore, studies in transgenic mice showed that modulation of one of the CGRP receptors (RAMP1) levels may contribute to migraine susceptibility and, in particular, to photophobia (45). More interestingly, a recent study showed that adenosine triphosphate (ATP)-gated P2X3 receptors may be involved in chronic pain (including migraine) and CGRP seems to enhance its transcription, involving also mediation by BDNF (46). BDNF is also a mediator of neuronal survival and plasticity of dopaminergic, cholinergic and serotonergic neurons, and its involvement in migraine seems clear as a mediator of the trigeminal nociceptive plasticity (9,11,47).
Our data support the hypothesis of an interaction between BDNF and CGRP in migraine susceptibility. The results obtained also reinforce the importance of replication studies with these genes in larger samples and in other populations. Interaction between candidate genes for migraine pathophysiology should be further explored, in order to have a broader knowledge of genes interplay in susceptibility for this common disease.
Footnotes
Acknowledgements
We would like to thank all patients and controls for participating in this study. We thank also Dr Serafim Guimarães and nurses Teresa Gomes and Palmira Gouveia for their assistance with sample collection, Paula Magalhães for technical assistance and Jorge Pinto-Basto and Zoltán Bochdanovits for their helpful suggestions.
This study was supported in part by grants of Fundação para a Ciência e Tecnologia (FCT) (POCTI-034390/99/FCT), Sociedade Portuguesa de Cefaleias (SPC) and the National Headache Foundation (NHF) (USA). CL was the recipient of an FCT fellowship (SFRH/BD/17761/2004). The authors report no conflicts of interest.