
Abstract
High mountains are climate change hotspots. Quickly rising temperatures trigger vegetation shifts such as upslope migration, possibly threatening mountain biodiversity. At the same time, mountain slopes are becoming increasingly unstable due to degrading permafrost and changing rain and snowfall regimes, which favour slope movements such as rockfall and debris flows. Slope movements can limit plant colonization, while, at the same time, plant colonization can stabilize moving slopes. Thus, we here propose that response of high mountain environments to climate change depends on a ‘biogeomorphic balance’ between slope movement intensity and the trait-dependent ability of mountain plants to survive and stabilize slopes. We envision three possible scenarios of biogeomorphic balance: (1) Intensifying slope movements limit vegetation shifts and thus amplify instability. (2) Shifting ecosystem engineer species reduce slope movement and facilitate shifts for less movement-adapted species. (3) Trees and tall shrubs shifting on stable slopes limit slope instability but decrease biodiversity. Previous geomorphic, ecological and palaeoecological studies support all three scenarios. Given differences in ecologic and geomorphic response rates to climate change, as well as high environmental heterogeneity and elevational gradients in mountain environments, we posit that future biogeomorphic balances will be variable and heterogeneous in time and space. To further unravel future biogeomorphic balances, we propose three new research directions for joint research of mountain geomorphologists and ecologists, using advancing field measurement, remote sensing and modelling techniques. Recognizing high mountains as ‘biogeomorphic ecosystems’ will help to better safeguard mountain infrastructure, lives and livelihoods of millions of people around the world.
Keywords
High mountain landscapes and ecosystems in a changing climate
High mountains are geomorphologically and ecologically highly diverse and dynamic environments. Their extreme relief with steep slopes, high tectonic activity, glacial imprint and climatic extremes promotes rapid erosion rates through a variety of geomorphic processes (Figure 1; Barsch and Caine, 1984; Herman et al., 2021; Hinderer et al., 2013). Mountain plants, arranged in elevation-dependent vegetation zones, are adapted to these extreme conditions (Humboldt and Bonpland, 1807; Körner, 2003) and contribute to the extraordinarily high biodiversity in high mountain environments (Antonelli et al., 2018; Rahbek et al., 2019). High mountain environments in a changing climate. Warmer temperatures and more intense precipitation are expected to increase slope movements through various geomorphic processes. The same changes also push plants to shift through key ecological processes. Thereby, slope movements can increase and biodiversity decrease, with significant impacts on people, their livelihoods and infrastructure in mountain environments.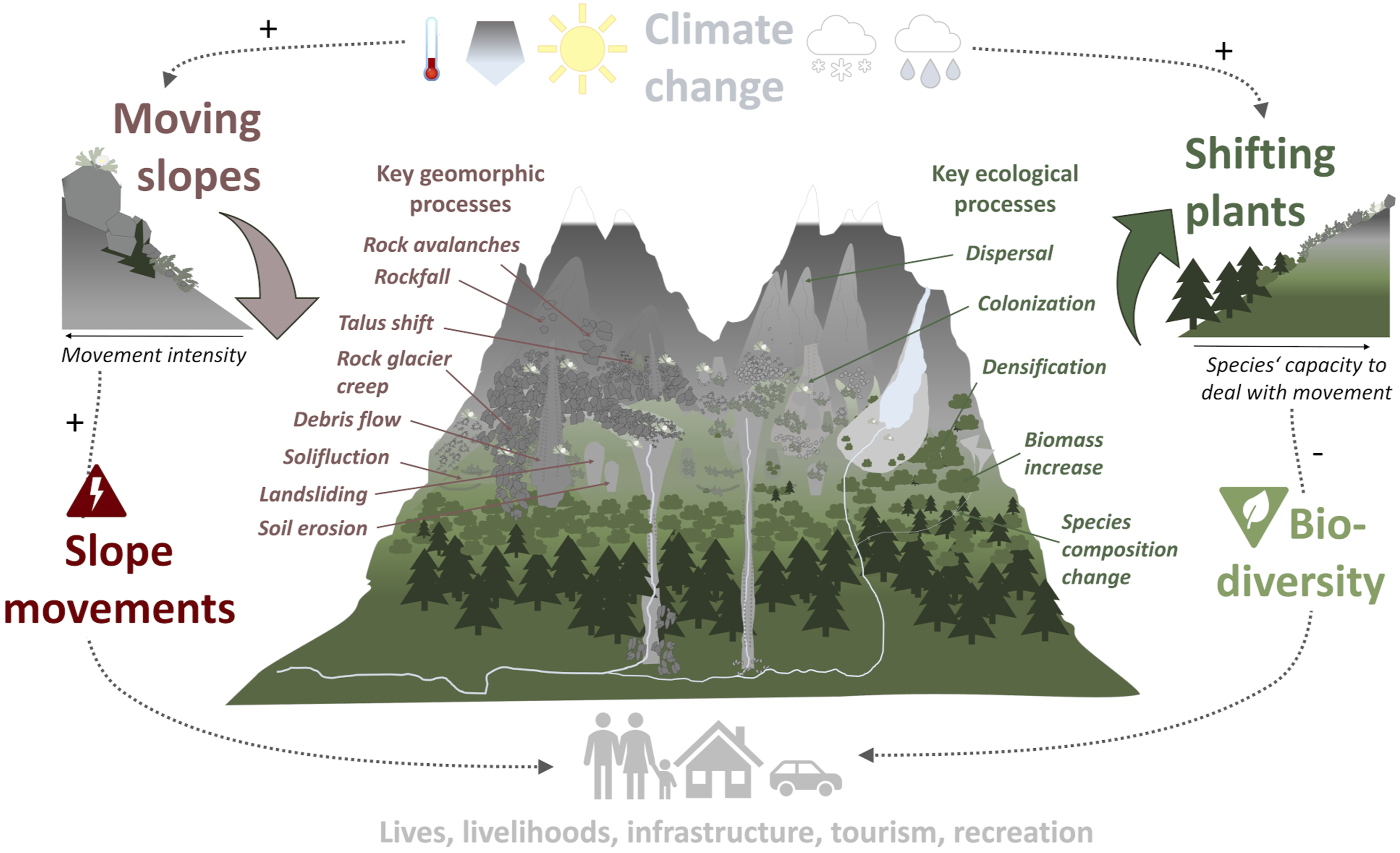
In high mountain regions, warming occurs at a much faster pace than in lowlands (Pepin et al., 2015, 2022), making them climate change hotspots (Hock et al., 2019a, 2019b). This accelerated warming causes fast mountain glacier melt, permafrost decrease and ground ice loss (Biskaborn et al., 2019; Rounce et al., 2023). Together with changing rain and snowfall regimes (Beniston et al., 2018; Gobiet et al., 2014), those cryospheric changes are intensifying mountain slope movements and associated slope instability (Figure 1; Arenson and Jakob, 2017; Clague et al., 2012; Stoffel and Huggel, 2012). Rockfall and landslide activity tend to increase in permafrost environments and to shift to increasingly higher elevations (Allen and Huggel, 2013; Draebing et al., 2022; Ravanel and Deline, 2011; Savi et al., 2020). Large and potentially very destructive rock-ice avalanches become more common (Jakob, 2022; Shugar et al., 2021) and rock glaciers tend move more quickly (Marcer et al., 2021) until the ice they contain has disappeared. Debris flows are expected to decrease in frequency but to increase in magnitude (Hirschberg et al., 2021; Stoffel et al., 2013) and to occur over most months in the future (Stoffel and Corona, 2018). More frequent extreme rainfall events can increase shallow landslide processes (Alewell et al., 2020; Geitner et al., 2021). When affecting high mountain communities, these intensifying slope movements turn into natural hazards.
At the same time, climatic changes also shift the conditions under which mountain plants can survive and thrive to higher elevations. Consequently, mountain plant species move their altitudinal distribution upwards, leading to increases in vegetation richness and density at high elevations (Gottfried et al., 2012; Grytnes et al., 2014; Steinbauer et al., 2018; Wipf et al., 2013). However, mountain plant species were also found to shift downslope, for example, in relation to changing climatic water balance (Crimmins et al., 2011; Rapacciuolo et al., 2014). Through dispersal, formerly bare ground, especially at higher elevations, can become colonized, and vegetation grows denser and taller. This happens both due to biomass increases of established species, and due to establishment of relatively taller and more competitive species from lower elevations (Bjorkman et al., 2018a, 2018b; Jaroszynska et al., 2023). Upslope shifts of woody species such as trees and tall shrubs cause reafforestation and ‘shrubification’ (Bader et al., 2020; Myers-Smith et al., 2011; Myers-Smith and Hik, 2018), which are key processes affecting ecosystem composition, structure and functioning. The entity of these processes are visible as widespread mountain greening observed from space (Choler et al., 2021; Rumpf et al., 2022). While vegetation productivity may rise overall, however, a number of species may fail to persist under altered biotic and abiotic conditions, or not shift successfully – or quickly enough – with climate change (Dullinger et al., 2012; Steinbauer et al., 2018).
While mountain geomorphologists investigate slope movements on mostly unvegetated, highly unstable alpine slopes, such as rockwalls, talus slopes and active rock glaciers (Arenson and Jakob, 2017; Ravanel and Deline, 2011), classic vegetation ecology tends to focus on stable and homogeneous conditions (Braun-Blanquet, 1964). Yet, in high mountain environments, slope movements are a very common and widespread phenomenon. Feedbacks between slope movements and shifting plants could play a strong role for the response of high mountain landscapes and ecosystems to climate change. For instance, mountain biogeomorphic research showed that strong slope movements can limit plant colonization and development (Eichel et al., 2016; Giaccone et al., 2019; Pérez, 2012). However, once vegetation manages to establish and grow, it can strongly reduce slope movements (Eichel et al., 2017; Haselberger et al., 2021; Marston, 2010). Yet, only few studies have hitherto considered the impact of slope movements on upslope plant migration (Macias-Fauria and Johnson, 2013; Randin et al., 2009; Resler, 2006), or how shifting plants may stabilize moving slopes (Greenwood and Jump, 2014; Moos et al., 2021; Sebald et al., 2019). Consequently, biogeomorphic feedbacks between moving slopes and shifting plants remain rather inadequately understood today. Yet, such an understanding would indeed be vital for securing mountain infrastructure, tourism, recreation and the lives and livelihoods of 671 million people living in mountain regions worldwide (Hock et al., 2019a, 2019b; Huggel et al., 2019).
In this progress report, we therefore develop the concept of ‘biogeomorphic balance’ to improve our understanding of how downward slope movements and upwards shifting plants interact in the context of climate change in high mountain environments. Based on previous geomorphic, ecological and palaeoecological research, we identify three biogeomorphic balance scenarios and discuss their variability in time and space. Finally, we propose three new research directions for joint ecological and geomorphologic research, which will help to advance understanding of climate change impacts on high mountain landscapes and ecosystems.
Response of high mountain environments to climate change – a question of biogeomorphic balance?
Biogeomorphic research on alpine lateral moraines slopes found close relationships between the magnitude and frequency of geomorphic processes and plant species response and effect traits that determine species’ survival of (resilience) and effect on (resistance) geomorphic disturbances (Eichel et al., 2016). Similar relationships between slope movement intensities and plant traits are found on mountain slopes (Figure 2). Relationships between slope movement intensity and plant species’ response and effect traits towards slope movements on high mountain slopes.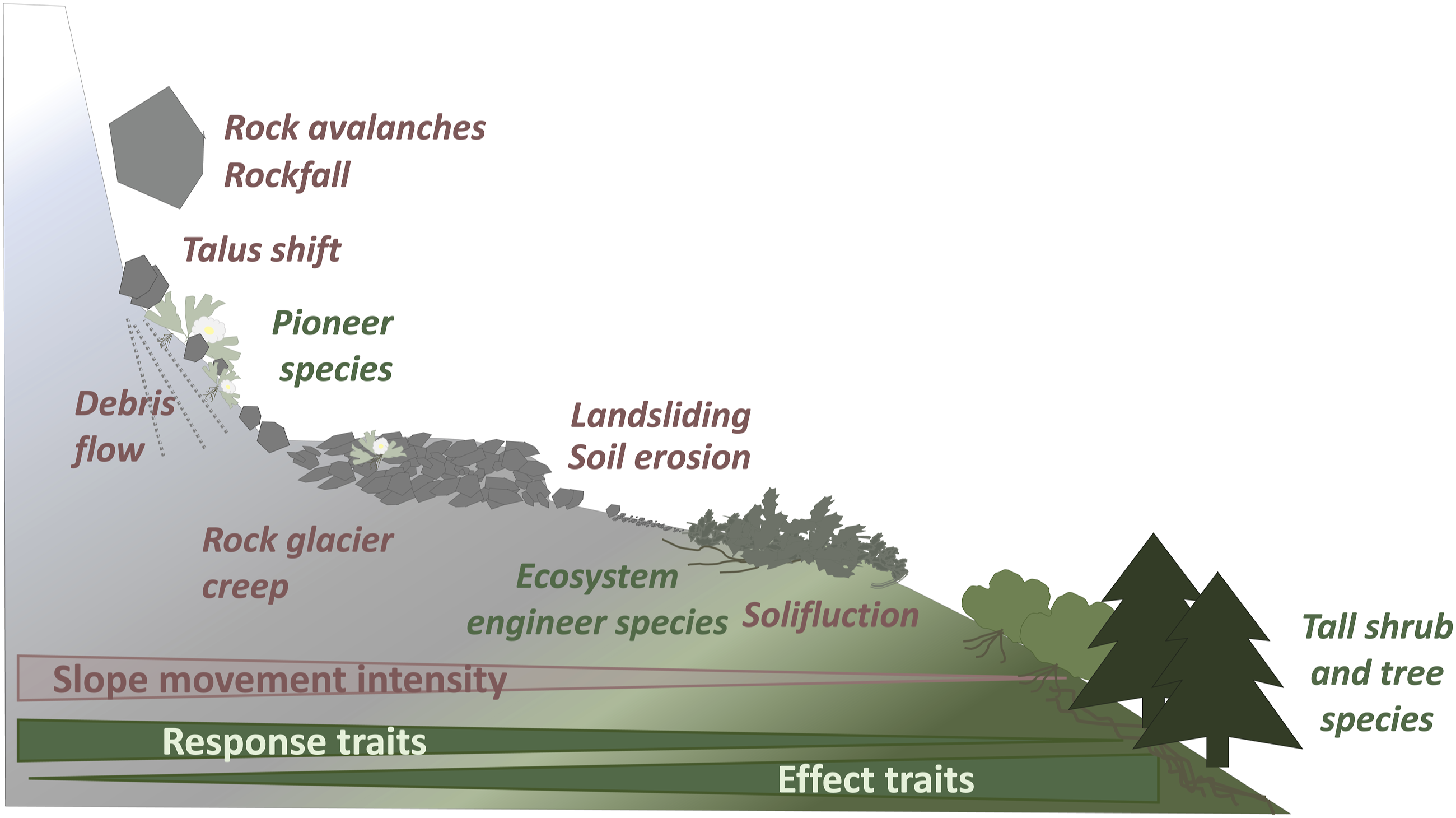
High magnitude or high frequency processes, such as rockfall, debris flows or rock glacier creep favour resilient pioneer species that tolerate movement or quickly regenerate after a disturbance (Cannone and Gerdol, 2003; Eichel et al., 2016). Flexible stems, dense fine root systems, quick growth rates and short life spans are typical traits enabling high resilience of pioneer species such as Linaria alpina or Ranunculus glacialis (Kutschera and Lichtenegger, 2013; Schröter et al., 1926). On slopes moving less intensely, for example, by solifluction (Eichel et al., 2017; Matsuoka, 2001; Price, 1974), soil erosion (Burylo et al., 2014; Frankl et al., 2020) and shallow landsliding (Löbmann et al., 2020; Pérez, 2012), a dominance of plant species that are less resilient but more resistant towards slope movements can be observed. By forming low lying mats, dense tussocks, rosettes or cushions, or by having close-set, low-lying stems, often extending through clonal growth, these ‘biogeomorphic ecosystem engineer’ species can not only survive but even reduce slope movement (Eichel et al., 2023; Haussmann et al., 2009; Pérez, 2009). Examples include prostrate shrubs (Dryas octopetala, Muehlenbeckia axillaris, Salix serpillifolia), graminoids (Festuca spp.), cushion plants (e.g. Azorella spp.) as well as certain herb (Anthyllis vulneraria) and shrub species (Coriaria angustissima, Salix hastata). On stable slopes, one observes a dominance of grassland, tall shrub and tree species that cannot deal well with slope movement, for example, during establishment or if soil moves in their rooting zone (Cannone and Gerdol, 2003). However, specifically montane shrubs and trees, such as Fagus sylvatica and Picea abies, have strong effect traits that stabilize moving slopes. Their large stems and biomass add vegetation resistance to flow and protect from rockfall, while their extensive root systems add root cohesion (Rickli et al., 2019; Stokes et al., 2005).
Consequently, we suggest that response of mountain ecosystems and landscapes to climate change will depend on a ‘biogeomorphic balance’ between slope movement intensity and the capacity of shifting plant species to survive and reduce slope movement, depending on their functional traits (Figure 3(a)). A ‘biogeomorphic balance’ concept for high mountain environments with three possible scenarios (A–C) in a changing climate. (a) Illustration of the biogeomorphic balance between moving slopes and shifting plants, depending on slope movement intensity and the trait-dependent ability of plant species to survive and reduce slope movements. (b) Scenario A: Intensifying slope movements limit vegetation shifts and thereby amplify. (c) Scenario B: Shifting biogeomorphic ecosystem engineer species reduce slope movement and facilitate vegetation shifts. (d) Scenario C: Shifting trees and tall shrubs on and onto stable slopes protect from slope movements but outcompete alpine species and reduce biodiversity.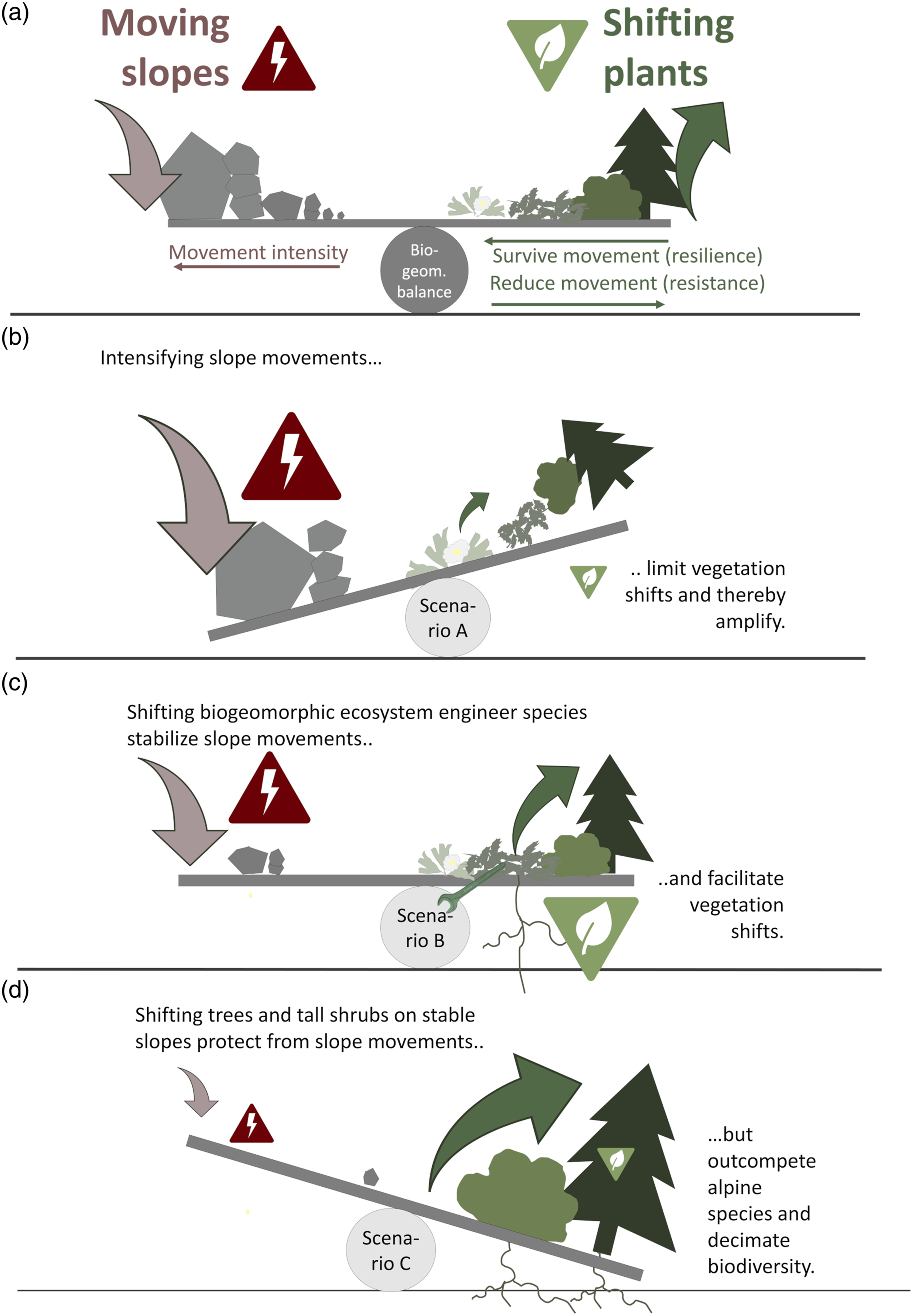
Three scenarios of biogeomorphic balance in high mountain environments
In a changing climate, we expect three possible scenarios of future biogeomorphic balance (Figure 3(b)–(d)). All scenarios are currently observed and were reconstructed to have happened during and after the Younger Dryas (∼12,500–5800 BP).
Scenario A: Intensifying slope movements limit vegetation shifts and thereby amplify
An increase in either the magnitude or frequency of rockfall, rock glacier creep, debris flows and other landslides over the coming decades (Hock et al., 2019a, 2019b; Ravanel and Deline, 2011; Stoffel and Huggel, 2012) could limit required plant shifts on local scales. High intensity slope movements can disturb or even remove existing vegetation (Pérez, 2010) or prevent its densification and biomass increase (Aalto et al., 2021). Plant species with traits that are not adapted to slope movements, such as tree and some tall shrub species (Macias-Fauria and Johnson, 2013; Myers-Smith and Hik, 2018; Resler, 2006), grassland and snowbed species (Bürli et al., 2021; Pérez, 2009) cannot establish on talus slopes affected by rockfall, snow avalanches and debris flows (Pérez, 2012) or on active rock glaciers (Burga et al., 2004; Stefano et al., 2021). Thus, when trying to migrate into upslope areas characterized by the most intense slope movements (Slaymaker and Embleton-Hamann, 2018), these species may not succeed and could thus be lost. Vegetation regression and declining population sizes of arctic-alpine species due to permafrost degradation, rockfall and landslide activity are currently observed at high elevations (Cannone et al., 2007; Carlson et al., 2017; Watts et al., 2022) and also occurred in the past. Pollen analysis found that during the Younger Dryas between 11,000 and 9800 BP, abundances of tree, tall and prostrate shrub species and overall vegetation cover decreased, while abundances of pioneer species increased (Tinner et al., 1996). High sedimentation rates reconstructed from alpine lake sediments indicate that this was caused by increasing slope movement activity. Thus, intensifying slope movements due to climate change could create a positive feedback loop further amplifying slope movements due to the removal of vegetation. Amplified slope movements could turn into natural hazards and enhance risk for mountain communities and infrastructure.
Scenario B: Shifting biogeomorphic ecosystem engineer species reduce slope movements and facilitate vegetation shifts
While several high intensity processes such as rock avalanches and rock glacier creep are not affected by plants, vegetation can stabilize lower intensity processes such as talus shift, soil erosion, landsliding and solifluction (Burylo et al., 2011; Eichel et al., 2017; Geitner et al., 2021; Pérez, 2012). When slope movement intensities decrease, for example, once rock glacier permafrost is gone (Marcer et al., 2021) or rockfall intensities decrease due to changing snow cover and duration (Draebing et al., 2014), biogeomorphic ecosystem engineer species could establish, densify or increase their biomass. Positive response of prostate engineer shrubs such as D. octopetala to elevated temperatures suggest that densification and biomass increase could happen quickly (Welker et al., 1997), while establishment, for example, of engineer cushion plants was found to take more time (Matteodo et al., 2013). With their dense above ground biomass close to the ground, engineer species protect the soil surface by intercepting rainfall and obstructing runoff (Burylo et al., 2011; Kervroëdan et al., 2021) and trap sediments (Eichel et al., 2023). Their roots and root associated mycorrhiza can play a key role for soil stability (Beeli et al., 2011; Norris et al., 2008a, 2008b; Vannoppen et al., 2015) and hold moving sediments in place (Eichel et al., 2023). Thus, especially densification, increase in biomass and cover, for example, by merging ecosystem engineer patches, would be efficient for slope stabilization (Eichel et al., 2016; Marston, 2010). Besides local stabilization, engineer species also improve soil conditions due to organic matter provision and nitrogen fixation (Eichel et al., 2023; Pérez, 2009) and even create small scale landforms such as solifluction steps, terraces and lobes (Eichel et al., 2017). In combination, those engineer effects promote establishment for other species (Butler et al., 2004; Cavieres et al., 2016; Resler, 2006; Zuber, 1968), which is often necessary for upslope migration of species from lower sites, for example, for treeline advance (Brodersen et al., 2019; Choler et al., 2001; Resler, 2006). Pine trees, for example, need facilitation by established plants or favourable microsites in the first 1–2 years to successfully establish (Batllori et al., 2009). Consequently, successful upslope migration of ecosystem engineer species could reduce slope movements and facilitate survival and migration for other species, thereby preserving biodiversity.
Remote sensing analysis shows evidence for increasing colonization and biomass at rocky habitats >2500 m in the Écrins (France; Carlson et al., 2017), while field studies using repeated plot sampling found that decreasing rock glacier movement encourages vegetation development (Cannone and Piccinelli, 2021). At the onset of the Younger Dryas (12,000–11,000 BP), lake sedimentation decreased while dwarf willow pollen increased (Salix herbacea, Salix retusa, S. serpillifolia; Tinner et al., 1996), suggesting that those prostrate shrubs acted as stabilizing ecosystem engineers on scree slopes.
Scenario C: Trees and tall shrubs shifting on and onto stable slopes protect from slope movements but outcompete alpine species and decimate local biodiversity
Slope stabilization, for example, by loss of permafrost and periglacial processes over the next decades (Aalto et al., 2014), could enable shrub and tree species to successfully shift, for example, by densifying and increasing their biomass in existing positions or shifting upslope onto previously moving slopes (Burga et al., 2004; Myers-Smith and Hik, 2018). Established trees facilitate establishment for other tree seedlings by providing protection and a favourable microclimate (Bader et al., 2020; Butler et al., 2007; Resler, 2006). Densifying forests protect downslope communities and infrastructure from debris flows, rockfall and snow avalanches (e.g. Lingua et al., 2020; Malik et al., 2013; Moos et al., 2018, 2019). Efficient debris flow and rockfall protection, for example, by reducing runout length is linked to high stem diameters or stem densities of tree species (Bettella et al., 2018; Guthrie et al., 2010; Michelini et al., 2017). However, upslope advance by competitive tall shrub and tree species and slope stabilization will reduce habitat area for light-demanding alpine species and limit downslope movement for those species (Choler et al., 2021; Rixen et al., 2007; Watts et al., 2022), potentially reducing biodiversity locally. Rapid shrub and tree upslope expansion is frequently reported for stable slopes at lower elevations (Filippa et al., 2019; Myers-Smith and Hik, 2018) or for stabilizing slopes at intermediate elevations, such as inactive and relict rock glaciers (Burga et al., 2004; Cannone and Gerdol, 2003). A few centuries after the Younger Dryas (9200–5800 BP), tree pollen started to dominate as temperatures increased, while pioneer and prostrate shrub pollen declined and lake sedimentation decreased (Tinner et al., 1996).
Future mountain biogeomorphic balances in time and space
High heterogeneity and elevational gradients in high mountain environments, coupled with geomorphic and ecological processes acting on multiple spatial and temporal scales (Malanson et al., 2019), suggest that future biogeomorphic balances will be variable in time (Situations T1–3, Figure 4) and space (Situations S1–3, Figure 4). Variability of future biogeomorphic balances with different possible situations in time (T1–3) and space (S1–3) on spatiotemporal scales from local/years-decades to regional/centuries-millennia.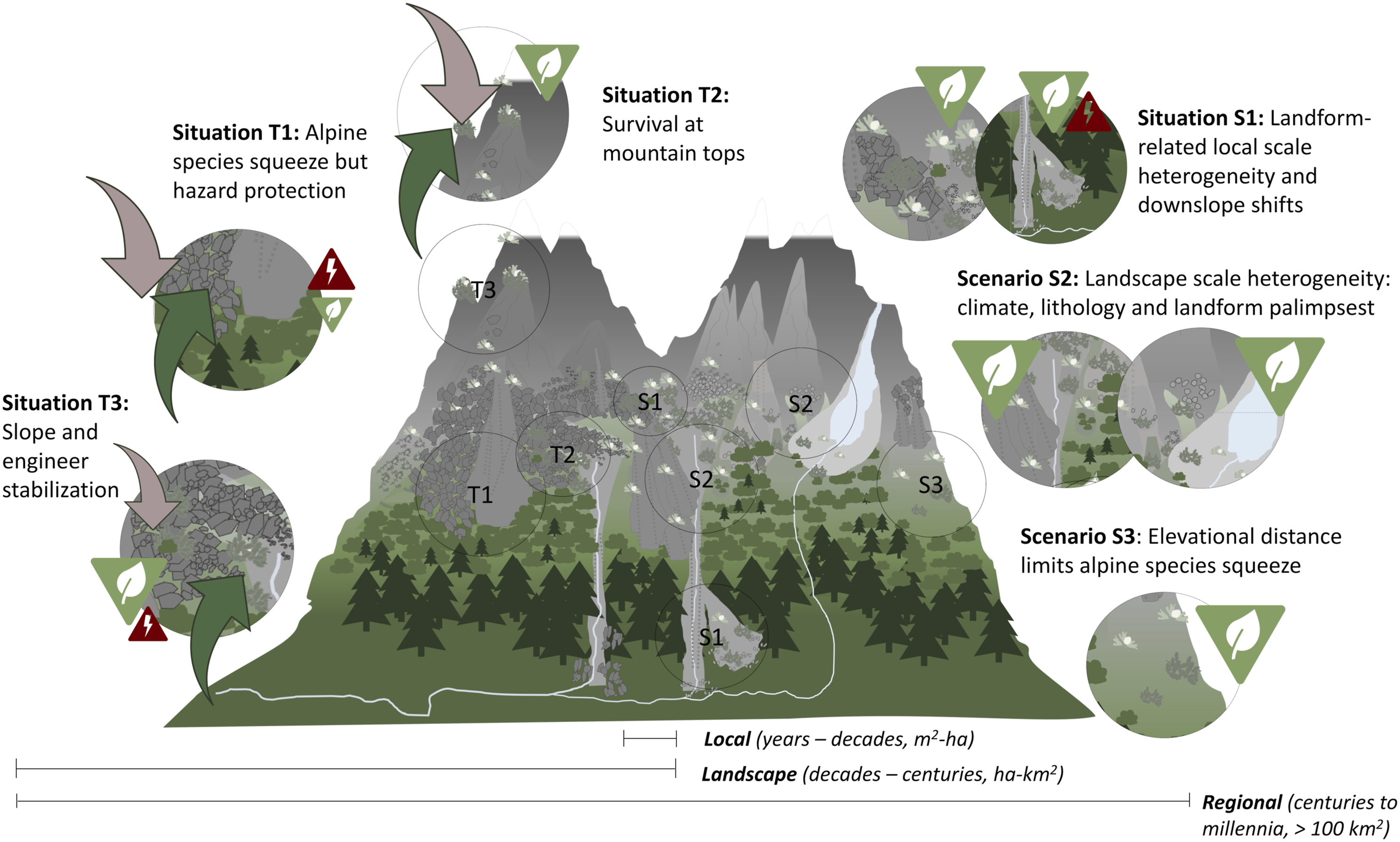
Biogeomorphic balances in time: The role of geomorphic and ecological response rates
Variable response rates and sensitivity of geomorphic and ecological processes to climate change (Dullinger et al., 2012; Knight and Harrison, 2023) could influence which biogeomorphic balances will dominate where over the coming decades to centuries on a landscape scale (Figure 4). Permafrost thaw and glacier melt are expected to accelerate further with rising temperatures over the next decades (Aalto et al., 2014; Rounce et al., 2023), directly causing increase in rock slope destabilization, rockfall frequencies and rock glacier destabilization (Marcer et al., 2021; McColl and Draebing, 2019; Savi et al., 2020). Likewise, shrubs and trees species respond quickly to climate change, visible in the form of tall shrub cover expanding into the alpine tundra (Formica et al., 2014) and widely upslope shifting treelines (Hansson et al., 2021). In combination, quickly responding unstable slopes and shifting competitive shrub and tree species could strongly reduce habitats for less competitive ecosystem engineer and possibly even pioneer species across the landscape (Situation T1, Figure 4). Thus, biodiversity could reduce rapidly. Yet, shifting tall vegetation could well counteract increasing rockfall and debris flow activity, thereby decreasing slope movements. Alpine engineer and possibly also pioneer species could survive at mountain tops (Situation T2, Figure 4), on which species richness strongly increased over the past decades (Steinbauer et al., 2018). However, this would require quick dispersal to the mountain tops, working well for propagules with achene or pappus appendages, such as Asteraceae species (Matteodo et al., 2013). If engineer species make it to the mountain tops, they might exclude less competitive pioneer species through their dense, impenetrable cover (Malanson et al., 2019, Butler et al. 2009). Once permafrost and glaciers disappear, slope movements are expected to decrease within decades to centuries (Aalto et al., 2017; Ballantyne, 2002; Vivero and Lambiel, 2019). If ecosystem engineer species survived until slope movements start to decrease, they could colonize the stabilizing surfaces from their refugia and actively contribute to stabilization once they reach a certain biomass or cover (Situation T3, Eichel et al., 2016). This could amplify the reduction of slope movements in this situation and locally protect biodiversity if new landforms are created.
Biogeomorphic balances in space: The role of environmental heterogeneity and elevational gradients
High mountain environments are highly heterogeneous and geodiverse from local to regional scales and extend over large elevation gradients (Bollati and Cavalli, 2021; Gordon, 2018).
Their local to landscape scale mosaic of habitats offers opportunities for many species to survive locally. Local scale heterogeneity is, for example, created by periglacial and glacial landforms such as solifluction lobes, rock glaciers, patterned ground and moraines (Situation S1). Due to strong variations in sediment properties, microtopography, microclimate and movement rates within and between landforms (Eichel et al. 2020), landforms provide habitats for many different plant species (Tukiainen et al., 2019) and thereby safeguard local (α-)diversity. Downslope expansion of geomorphic processes such as debris flows and snow avalanches can create additional habitats and refugia at the local scale (Situation S1, Gentili et al., 2015; Körner, 2003) and also could also promote downslope dispersal and plant shifts (Crimmins et al., 2011; Raffl et al., 2006; Rapacciuolo et al., 2014). However, downslope expansion of geomorphic processes can also increase natural hazards (Stoffel and Huggel, 2012).
On a landscape scale, additional environmental heterogeneity is created, for example, by varying solar energy input at different aspects (Kulonen et al., 2018; Scherrer and Körner, 2010) and topography- and cryosphere-related climatic variability (Matthews, 1992; Patsiou et al., 2017) (Situation S2). Together with differently aged landform palimpsests resulting from previous glaciations and deglaciations (Stroeven et al., 2013), this will likely ensure that increasing slope movements will not act as a landscape scale barrier for upslope migrating plant species, preserving landscape-scale (γ-)diversity. Further attention should be given in this context to solifluction processes, which are moving soil extensively across the landscape (Del Vecchio et al., 2022; Rouyet et al., 2021) and could therefore act as a more widespread migration barrier than other geomorphic processes.
On a landscape to regional scale, the vertical distance between intensifying slope movements at highest elevations and the upslope migrating tall shrubs and trees from the montane to subalpine zone could serve as a buffer for biodiversity decline (Situation S3). Even if slope movements and upslope tree and tall shrub migration both intensify within the coming decades, there might still be sufficient space for pioneer and engineer species to survive in the shrinking alpine zone until slope movements decrease with loss of permafrost and glaciers.
A mosaic landscape preserving biodiversity was reconstructed to have occurred directly following the Younger Dryas (9800–9200 BP). Over several few centuries, a mosaic of open Larix decidua stands, Juniperus nana shrublands and alpine meadow, snowbed and debris communities (Caryophyllaceae, Rumex spp., Salix spp. and D. octopetala) characterized the alpine treeline (Tinner et al., 1996).
Go or grow? New research directions for high mountain environments
The biogeomorphic balance scenarios and their variability described in this contribution indicate that also when taking biogeomorphic feedbacks into account, increasing slope movements and biodiversity loss remain a possible consequence of climate change. At the same time, however, our biogeomorphic balances suggest that we might also see positive developments in terms of natural hazards protection by advancing treelines and/or the preservation of alpine biodiversity in geodiversity-created (micro)refugia due to biogeomorphic feedbacks.
To successfully unravel (future) biogeomorphic balances in time and space and better apprehend response of high mountain environments to climate change, we suggest three new research directions. Resulting improved understanding of biogeomorphic balances in a changing climate will help to achieve natural hazard protection and biodiversity conversation at the same time.
Biogeomorphic balance mechanisms: Linking plant traits to slope movement intensities
The biogeomorphic balance is largely dependent on whether plant species can establish on and/or stabilize moving slopes. This in turn depends on response and effect traits of high mountain plant species (Eichel et al., 2023), their associated mycorrhiza (Graf et al., 2019) or their community (Pohl et al., 2009). Knowledge on adaptations of high mountain species to slope movements exists already for a long time (Schröter et al., 1926), but relationships between plant traits and movement intensities have rarely been quantified. To achieve this, measurements of mountain plant traits, facilitated by well-standardized methods (Freschet et al., 2021; Pérez-Harguindeguy et al., 2013), could be carried out on slopes with known movement rates increasingly provided by new techniques such as terrestrial laser scanning, uncrewed aerial vehicle (UAV) surveys and InSAR (Hartl et al., 2023; Hendrickx et al., 2020; Rouyet et al., 2021). While extensive plant trait databases exist (Bjorkman et al., 2018a, 2018b; Kattge et al., 2011, 2020; Maitner et al., 2018), data availability is often limited for alpine species and many important biomechanical traits, such as root tensile strength or modulus of elasticity, are far from being included routinely in ecological databases. Yet, the insights that one can gain on how and which mountain species deal with slope movements will ultimately help efforts of stabilizing moving slopes, for example, by using nature-based solutions (Norris et al., 2008a, 2008b; Viles and Coombes, 2022), but also to protect biodiversity by identifying species that cannot cope with very active or very stable slopes.
Biogeomorphic balance patterns: Detecting and mapping biogeomorphic balances in time and space
To unravel future biogeomorphic balances and their impacts on slope movements and biodiversity, we need to better understand which scenarios and situations are likely to happen when and where. Monitoring and reconstructing linked geomorphic and vegetation changes can help us to better understand biogeomorphic feedbacks and balances. Combining ecological and geomorphic field techniques, such as repeated vegetation surveys (Cannone and Piccinelli 2021; Grabherr et al., 2000) and continuous geomorphic monitoring (Belli et al., 2022; Mourey et al., 2022) can determine decadal scale biogeomorphic dynamics on local scales. Especially UAV surveys and dendroecology are of great value for biogeomorphic research as they can monitor or reconstruct geomorphic (De Haas et al., 2021; Favillier et al., 2018; Stoffel, 2010) and vegetation changes (Francon et al., 2020; Wei et al., 2021) at the same time. Satellite remote sensing can help to reconstruct and monitor biogeomorphic dynamics on landscape scales (Betz, 2021; Marchetti et al., 2020), with high spatial and temporal resolutions (e.g. 0.5 m resolution, multiple visits per day for Planet satellites) especially for the past few years, and lower spatial and temporal resolutions for the past decades (e.g. 30 m for Landsat satellites, images once a month). Forward simulation modelling can help looking into the future, though ecological focus on statistical models (e.g. Phillips et al., 2006) and geomorphic focus on process-based models complicates their integration. Recent studies incorporated geomorphic properties and processes into statistical species distribution modelling (e.g. Bailey et al., 2018; Randin et al., 2009) and vegetation dynamics into process-based geomorphic models (e.g. Karssenberg et al., 2017; Schmaltz et al., 2019). Given that statistical models rely on current data, their ability to predict the future might be limited (Del Vecchio et al., 2022), making combined process-based models, such as used by Moos et al. (2021) to assess plant shift effects on future rockfall risk, a better choice. Increasingly and freely available topographic, climatic, microclimatic and permafrost data (e.g. Kenner et al., 2019; Lembrechts et al., 2020; Michel et al., 2021) now allow assessment of local-to-regional scale factors influencing both timescales and distributions of biogeomorphic balances.
Biogeomorphic balance understanding: Recognizing high mountains as biogeomorphic ecosystems
To implement the first two new research directions and to successfully integrate their findings, an overarching biogeomorphic perspective is needed for high mountain environments. We are convinced that a ‘biogeomorphic ecosystems’ (Balke, 2013; Corenblit et al., 2015) approach to high mountain environments will help to unify and streamline geomorphic and ecological research that come with different viewpoints, terminology and methodology (Haussmann, 2011). Recognizing mountain environments as biogeomorphic ecosystems with inherent, frequent and regular physical (geomorphic) disturbances will mean that ecologists explicitly need to investigate geomorphologically disturbed areas instead of focussing on stable ground (e.g. Cullen et al., 2001). Likewise, geomorphologists need to realize that not only plants but also plant species matter as they respond to and affect geomorphic processes differently. Thus, it is well worth to assess which species are covering a site of interest to geomorphologists.
Footnotes
Acknowledgements
We thank Maarten Kleinhans, Daniel Draebing and Oscar van Vliet for many interesting discussions and feedback on climate change effects in mountain regions and the concept of biogeomorphic balance. We thank two anonymous reviewers for their constructive and helpful comments on our manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: J. Eichel acknowledges support for this study from the Nederlandse Organisatie voor Wetenschappelijk Onderzoek (NWO) grant VI.Veni.212.125. M. Stoffel and S. Wipf acknowledge the Research Commission of the Swiss National Park (FoK-SNP) for supporting his work on geomorphic process activity and its interaction with trees, and her work on alpine plant community shifts, respectively.