
Abstract
We studied species composition and spatial distributions of tree species, and the underlying topography and soil, in subtropical forests of northwest Belize, a region in the Maya Lowlands. Our goal was to learn how much the spatial distributions of species vary and are predictable over the landscape. The study was done in old-growth, subtropical moist forest on limestone-derived topography and soil. We identified to species all trees ≥10 cm DBH in 209 400-m2 plots. For each plot, we characterized topographic setting and analyzed soil nutrients and texture. We recorded 3,984 individual trees of ∼140 tree species and used the 3,775 individuals of the 69 species occurring in ≥5 plots in multivariate analyses, including Nonmetric Multidimensional Scaling (NMS). NMS showed that 73% of the variation in species composition per plot was associated with the first three ordination axes. Sixteen out of the 34 quantitative variables we measured were correlated at R 2 > 10% with the axes. Of the categorical variables, Topographic Class was strongly associated with species composition, and USDA Texture Class less so. Of the 69 focal tree species, the abundances of 21 were correlated at R 2 > 10% with one or more axes of the NMS ordination. Importantly, these 21 species accounted for 68% of all individual trees sampled in the 209 plots. Twenty-three species were indicators of particular topographic and soil classes. We conclude that patterns of tree species distribution are strongly and predictably associated with different topographic and soil conditions in this landscape. In the past, the ancient Maya could have used this type of predictable plant–soil relationship to optimize their agriculture. In the future, our results are a basis for predicting local shifts in tree species distributions due to climate change.
Keywords
Introduction
The Maya Lowlands extend from the northern end of the Yucatan Peninsula south through northern Guatemala and parts of Belize and Honduras. Research on the Lowlands’ vegetation has a long history (e.g., Bartlett, 1932), but modern quantitative studies are few in such a large region for which there is great interest in ancient Maya land use and modern management (Islebe et al., 2015). Especially informative are multivariate analyses that link data on tree species composition to data on environmental variables in forests. These analyses reveal the numerical strength of relationships between tree composition and the environment. Such results are needed for modeling the response of forest composition to changes in the environment due to disturbance and climate change (e.g., Mod et al., 2016).
In numerous studies across the tropics, the spatial distributions and abundances of a significant portion of tree species are correlated with gradients of topography and soil (reviewed in Baldeck et al., 2013). These correlations likely reflect differential adaptations among species to the variety of topographic settings and soil characteristics in forest landscapes (e.g., Brenes-Arguedas et al., 2008; Palmiotto et al., 2004). Several studies in the Maya Lowlands have described gradients of tree species on the topographic continuum but did not quantitatively link tree and environmental data (Furley and Newey, 1979; Lambert and Arnason, 1978; Lundell, 1937; Shepherd, 1975; Testé et al., 2020; Wright et al., 1959). For the Lowlands, we know of just three previous multivariate studies that do link these data, one of them unpublished (Hightower, 2012; Schulze and Whitacre, 1999; White and Hood, 2004). In this paper, we present multivariate analyses of tree species composition, soil characteristics, and topography in the varied terrain of the Rio Bravo Conservation and Management Area (RBCMA), in northwestern Belize. Our research questions were: Are tree species in the RBCMA distributed randomly, or are they distributed in predictable patterns on the landscape, and, how are tree species distributions and abundances on the landscape quantitatively associated with topographic position and/or soil characteristics?
Based on our results, we comment on how ancient Maya land use may affect the patterns we observe today, and we discuss the significance of those patterns for visualizing ancient Maya land use as a “managed mosaic,” in which the Maya tailored their land use to specific soil types and topographic settings (Fedick, 1996; Ford and Nigh, 2015). We also discuss how these quantitative results can help to predict future spatial distributions and abundances of tree species in a changing climate.
Methods
Study area
We conducted this study in the Rio Bravo Conservation and Management Area (RBCMA) in northwest Belize (Figure 1). The 100,400 ha RBCMA is managed by the Programme for Belize for biodiversity conservation, sustainable uses, and environmental education (Programme for Belize, 2022). The reserve is centered at about 17°50’ N latitude. At La Milpa Lodge in the RBCMA, the mean annual temperature for the period 2006–2014 was 25°C (Programme for Belize, 2022). Total annual rainfall averaged 1499 mm during 2006–2014, with most of the rain falling between May and December, the remaining months being significantly drier. The RBCMA is in the “subtropical moist” life zone (Holdridge Life Zone System, Hartshorn et al., 1984). Locations of the western area of the Rio Bavo Conservation and Management Area (RBCMA) and of the Chawak But’o’ob (CB) and Dos Hombres to Grand Cacao Transect (DG) study sites. The RBCMA boundary is in red; the western area is c. 34,000 ha of the 104,000 ha RBCMA. Land surface in dark green is at c. 20 m asl; surface in darker brown near Guatemala border is at c. 200 m. Inset shows approximate coverage of main map. RBCMA boundaries on the map are not official. Study sites are not to scale.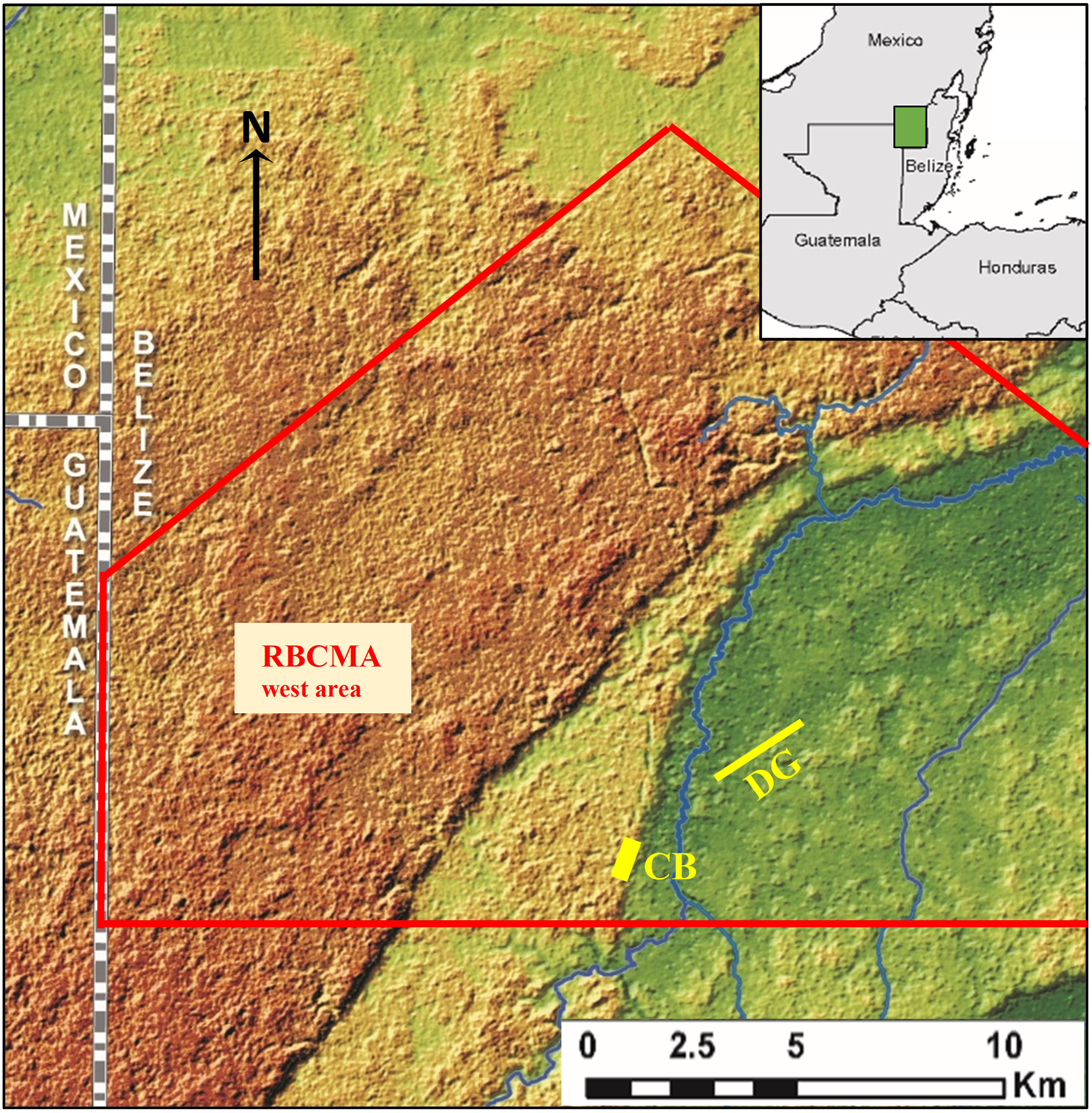
The RBCMA is on the Yucatan Platform, consisting of marine sediments consolidated during the Eocene (Dunning et al., 2003). The Platform began rising from the sea and exposing the area of northern Belize in the Pliocene. Faulting has produced escarpments 30–60 m high, and dissolution of the parent limestone and sedimentation has produced hills and valleys and gently rolling or level areas. Elevation of the RBCMA ranges from about 20 m asl in the northeast to 200 m asl near the Guatemala border. Soil parent materials are “fragmented limestone and chalk” (Wright et al., 1959).
The RBCMA has a mosaic of forest types, including upland forests on hills and other well-drained sites, with upper canopy height mainly 18–20 m and emergent trees 25–30 m tall; “bajos,” which are shallow, poorly drained basins in upland areas, with canopy 6–10 m; riparian forests with a low main canopy 10–15 m but scattered large trees to 30 m; palm-dominated forests with trees to 25 m; and transition forests among these (Brokaw et al., in review). Our study plots (Section 2.2) were mainly in upland forest, with some in transition forests between upland and poorly drained sites. The structure and composition of the RBCMA forest is similar to that of the forest at Tikal, Guatemala (96 km SW of RBCMA), described by Schulze and Whitacre (1999). Dunning et al. (2003) provide an overview of the regional environment.
Environmental history
The climate described above has prevailed since about 4,000 BC and supported subtropical forest (Frappier, 2016; Leyden, 2002). However, this forest has been subjected to hurricanes (Friesner, 1993), decades-long droughts (Hodell et al., 2005), deforestation by the ancient Maya (Beach et al., 2018; Jones, 1994), and natural reforestation after the Maya decline (Anselmetti et al., 2007). Apparently, the most recent hurricane that may have strongly affected the study area passed 65 km to the northeast in 1955 (Friesner, 1993). Hurricane Richard, in 2010, affected areas in the RBCMA but did not significantly affect the study area (Map 1 in Programme for Belize, 2022). Research nearby in Mexico shows that hurricanes typically cause dramatic structural damage but have little effect on dominant tree species composition (Navarro-Martínez et al., 2012; Vandecar et al., 2011).
Judging from land use history and from forest and soil characteristics, we believe that the RBCMA largely supports old-growth forest, having attained great age, with little disturbance. Ancient Maya activity is well-documented in the region (Guderjan, 2007; Scarborough et al., 2003), but since the abrupt decline of the local Maya, c. 950 AD (Hammond et al., 1998) there has been little human disturbance. Early logging for mahogany (Swietenia macrophylla) with axe and oxen transport would have affected a limited area (Weaver and Sabido, 1997). By the era of chainsaws and trucks, the Belize Estate & Produce Company owned the area and logged just a few tree species, causing relatively little forest damage (cf. Whitman et al., 1997). And the Company limited other uses: in 1953, there were only about 10 farming families, clustered in one area of the present RBCMA (Figure IX in Wright et al., 1959). Since the RBCMA was established in 1988 there has been no logging in our study area. During the 1980s, farmers cultivated small plots in three areas of the future reserve but not in our study area (N. Brokaw, personal observation). Present day characteristics that indicate old-growth status (Budowski, 1965) of forest in the RBCMA include (1) tall and large-diameter trees for this environment, (2) complex three-dimensional forest structure, (3) large lianas, and (4) and large fallen trees (N. Brokaw, personal observation). There are also abundant seedlings and saplings of species that will replace abundant conspecifics in the canopy, suggesting stability of species composition (unpublished data). In each of two 1-ha plots in representative upland forest in the RBCMA, eight tree species are among both the 10 most abundant upper canopy species (stems ≥10 cm DBH: diameter at breast height) and among the 10 most abundant species with stems 1–10 cm DBH. Lastly, soil characteristics also indicate old-growth forest (Beach et al., 2018).
Study sites and plots
We established two study sites in old-growth forest in the RBCMA to examine tree distributions in relation to topography and soil. The sites were at locations being mapped and excavated by archaeologists: Chawak But’o’ob and the Dos Hombres to Gran Cacao Transect. The sites are c. 3 km from each other at their closest approach (Figure 1). At both sites, we used a systematic, rather than random, arrangement of sample plots to capitalize on archaeological surveys and to more easily position and relocate points in this dense forest on sometimes steep terrain.
Chawak But’o’ob (CB) extends north–south for 1300 m along the 400-m wide slope (horizontal distance) of the east-facing Rio Bravo Escarpment. The escarpment drops 55 m at an average inclination of 13.7%, which includes steeper slopes and level benches. Near the base of the escarpment, there is an abrupt transition to the Rio Bravo floodplain. Soils are “grey brown or brown grey rocky clay loam” and “grey brown or brown grey gravelly clay loam on rocks,” according to the soil map of Wright et al. (1959). The site was occupied by ancient Maya “commoners” (Walling et al., 2020). Archaeological remains are prevalent but modest. The forest has a main canopy 18–20 m in height and emergent trees to about 25 m, about 52 tree species ha−1 (among all trees ≥10 cm DBH; unpublished data), frequent lianas (large, woody vines), occasional epiphytes, and an understory with many shrubs and small palms.
At CB, we established five parallel north–south transects, each separated by 100 m. The most westerly transect was near the top of the escarpment and the easterly transect was near the escarpment base. The transects varied in length from 150 to 1300 m, to maximize coverage of the irregular shape of the escarpment face. Every 50 m along each of the transects, we established a 400-m2 circular tree sample plot, for a total of 88 plots.
At the time of our study, the Dos Hombres to Gran Cacao Transect (DG) was a 2150 m long, 150 m wide “belt” transect, established for an archaeological survey conducted by M. Cortes-Rincon (McFarland and Cortes-Rincon 2019). It passed through scattered hills supporting upland forest, and low-lying terrain mainly supporting forest transitional between upland forests and bajos. Upland forest soils and forest physiognomy are similar to those described above at CB. Bajo soils are “pale brownish-grey or pale grey clay” soils that are sometimes flooded in the wet season and edaphically dry (holding water too tightly for effective plant use) in the dry season (Wright et al., 1959). The wetting and drying creates “gilgai,” a hummocky micro-relief. Bajos support a low forest of dense stems and often a sedge (Cyperaceae) ground cover. Archaeological sites are absent along most of the DG transect, but there are some small plazas with low remains (generally c. 1 m height).
At DG, we established a 400-m2 circular sample plot every 50 m along the 2150 m baseline and, with a few exceptions, similar plots at 50 m distance from baseline plots at points on axes perpendicular to and on both sides of the baseline. This made 121 plots at DG and 88 at CB, for a total of 209 400-m2 plots, their area summing to 8.36 ha.
Field methods
We defined a tree as a self-supporting, woody plant ≥10 cm DBH (diameter at breast height; i.e., at 130 cm along the tree stem from rooting point). (Archaeological workers did not cut trees ≥10 cm DBH.) At each plot, we counted and identified all trees whose bases were at least half inside the plot. Trees with multiple stems were counted as one individual, but all stems ≥10 cm DBH were measured. We estimated the DBH of palms that had stems sheathed in leaves. Basal area was calculated as the cross-sectional area of stems (including multiple stems), using DBH and the formula area = πr 2 . Most trees were identified to species in the field based on experience and previously collected voucher specimens (Brokaw et al., 2021). We collected unknowns for identification.
We classified the topographic setting of each 400-m2 plot as hilltop, rolling to level, slope, slope bottom (on slope but near bottom), hill base (just off slope), lowland (low-lying and level, no sign of flooding), bajo (gilgai and/or sedge evident), or floodplain (sometimes flooded). We measured the predominant inclination (degrees) of the plot (if any) with an inclinometer.
Soil sampling and analysis
We took one soil sample in each 400-m2 plot, at 1 m SE of the center point. We removed the surface litter and dug a ∼20 cm wide hole, to the depth of bedrock or a maximum of 20 cm, from which we removed soil from the top to the bottom of the hole, to sample more completely the soil zone used by tree roots. We mixed the sample in the field and extracted from the mix about one cup (∼237 cc) of soil. Soil samples were air dried, complete drying being impossible under field conditions. (Remaining moisture in the samples may have limited the precision of soil parameter measurements. With more precision, we might have seen stronger associations of environmental variables with plot species composition.) We did not measure soil water since it is highly variable temporally, and thus is difficult to measure meaningfully in the field (Kursar et al., 2005).
We sent the 209 samples to the Cornell Nutrient Analysis Laboratory (Ithaca, New York) for analysis. Analysis of soil particle composition (soil texture) (Kettler et al., 2001 in Moebius-Clune et al., 2016) included determination of percent sand, silt, and clay, and USDA Soil Texture Class (USDA, 1987). Total nitrogen and total carbon were determined following Zimmerman et al. (1997). Loss on ignition (LOI) and organic matter were determined after Schulte and Hoskins (2009). The Mehlich III extraction (Mehlich, 1984; Wolf and Beegle, 2009) was used for standard nutrient analysis, which was carried out following U.S. EPA (1996).
Analysis
Our analysis followed this sequence, explained in more detail below: (1) we arranged the 209 sample plots in a primary matrix of plot species composition (presence and abundance of each tree species per plot) and a secondary matrix of the measured values of environmental variables per plot; (2) using the primary matrix we created an ordination that arrayed the sample plots along several axes in proportion to their similarity in tree species composition (comparing each plot with all other plots); (3) using the secondary matrix, we identified which quantitative environmental variables were correlated (Pearson) with the axes of the tree plot ordination; (4) we associated individual tree species abundances with the ordination axes and with the quantitative and categorical environmental variables; and (5) we determined which species were Indicator Species (strongly associated with certain categorical variable classes).
Data arrangement
Data from the two study sites were combined for analysis, because the sites were near each other in space and on the same parent material, only differing in that DG had more plots in low-lying topographic classes than did CB. For the primary matrix, we included only tree species that occurred in ≥5 of the 209 plots and that were identified at least to family level. We constructed three primary matrices, one each based on: the number of individuals, presence versus absence, or basal area, for each species in each plot.
Environmental variables included in the analysis.
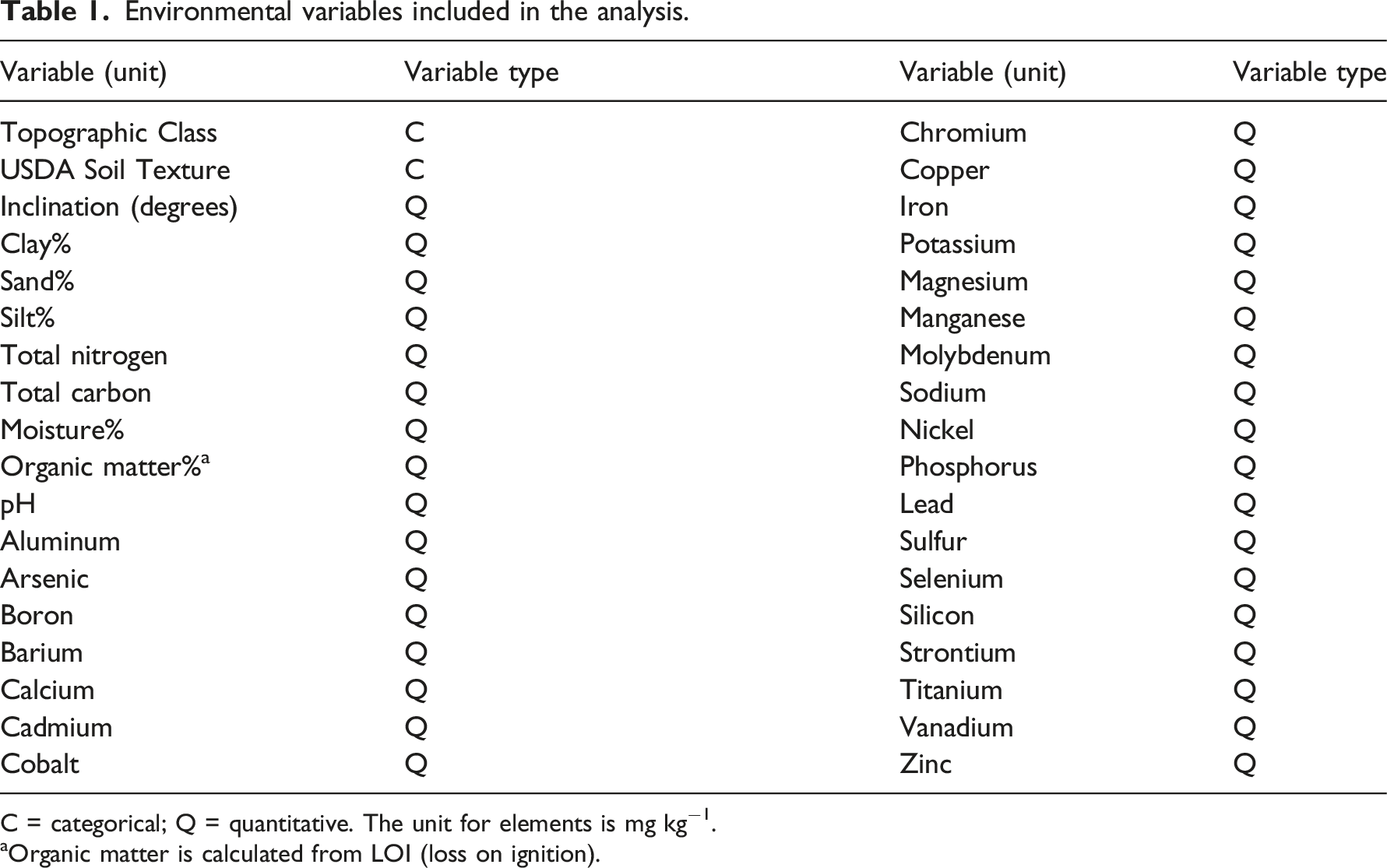
C = categorical; Q = quantitative. The unit for elements is mg kg−1.
aOrganic matter is calculated from LOI (loss on ignition).
Number of plots in each class of categorical variables (209 total plots).

Ordination of plots based on tree species composition
To determine if tree species are distributed randomly or in consistent, predictable patterns on the landscape, we created ordinations of the 209 tree plots. Ordination is a technique that arranges sample plots along axes in one or more dimensions, in proportion to plot similarity (each plot compared to all other plots) in tree species composition in primary matrices. Axes are orthogonal, that is, not correlated with each other, and represent dimensions that account for major fractions of the differences among plots. The first axis accounts for the highest proportion of variation among plots, with the second axis accounting for part of the residual variation, and so on for subsequent axes. For our ordinations, we used Nonmetric Multidimensional Scaling (NMS), which is suitable for non-normally distributed data. We used PC-ORD 7.08 (McCune and Mefford, 2018) for all ordinations in this paper.
The first step in our NMS was to determine Sørenson (Bray-Curtis) “distances” between plots (Peck, 2016), based on the degree of plot similarity in the three primary matrices: tree species composition, frequency, and basal area, respectively. The second step used these distances to perform the ordinations and determine the number and importance of axes for each of the three primary matrices.
The importance of an axis is based on the percent variation among plots accounted for by that axis, measured as R2. How well the NMS ordination preserves the observed rank-order distances between plots is evaluated as “stress,” or distortion in the data (Peck, 2016), with lower values indicating less distortion. NMS performs numerous iterations of the ordination, in which one to six axes are sequentially computed and compared for the amount of variation explained and the amount of stress.
In our study, ordination based on species composition resulted in lower final stress than ordinations based on species frequency or basal area, that is, species composition provided the best association of plot distances with the axes; so we used species composition for all analyses. For the species composition data, three axes had the best combination of variation in species composition and low stress. Stress decreased from 38.009 to a minimum of 18.453, or to below 20.0, as recommended (Peck, 2016). The inclusion of a fourth axis decreased stress by 3.45, below the recommended minimum stress reduction of 5, so adding a fourth axis was not justified (Peck, 2016). We repeated the NMS ordinations based on species composition five times to verify the consistency of the stress calculations. The final stress for all five runs ranged from 18.453 to 18.459, indicating consistency of the NMS. One NMS, with final stress = 18.458, is featured in the results of this paper. The selected NMS ordination also yielded “plot scores,” that is, plot positions on each axis, to be used in further analyses.
The third step in NMS is to test whether the ordination of observed data could have been produced at random. We used a Monte Carlo test to compare the results of 250 iterations of the observed data with 249 iterations of randomized data (no penalty for ties, and using time as the random number seed [Peck, 2016]). The p value indicates the probability of obtaining a lower final stress in an iteration using randomized data (i.e., by chance alone) than using the observed data (McCune et al., 2002). A p ≤ 0.05 indicates that species composition differs significantly (is not random) among the plots.
In our ordination diagrams, the points represent the 209 plots, and the distance from one point to another in the diagram indicates the degree of similarity among plots in tree species composition (close points more similar, distant points less similar). The ordination axes can be correlated with independently measured environmental factors.
Species composition and quantitative variables
To assess the association between the primary matrix of species composition and the quantitative environmental variables in the secondary matrix (Table 1), we used a Mantel Test (Peck, 2016). Because the quantitative environmental variables were measured on different scales, we relativized their values as percent of the maximum value for each variable (0–100%). For the Mantel test, we used Sørenson distances for both the species data (primary matrix) and quantitative environmental data (secondary matrix), and we used a Monte Carlo test for significance of the association.
To further characterize the relationships between species composition and quantitative environmental variables, we also used the Mantel test to assess the strength of association between plot scores on each axis in the NMS ordination and the matrix of quantitative environmental variables. For this test, we reversed the matrices. For the primary matrix, we used Sorenson distances among plots in terms of relativized quantitative environmental variables. For the secondary matrix, we use plot scores on each axis separately. We adjusted negative values in the NMS plot scores and potential outliers by using relative Euclidean distances (Peck, 2016). We also determined the Pearson correlations (R2 >10%) between individual quantitative variables per plot and each of the three axes of the NMS ordination. Tests of significance were not carried out, since with a sample size of 209 plots, correlations will be significant at a low value, possibly below ecological relevance (McCune and Grace, 2002).
Species composition and categorical variables
The association of tree species composition per plot with Topographic Class or USDA Soil Texture (Tables 1 and 2) was evaluated using Multi-response Permutation Procedure (MRPP; Peck, 2016), a non-parametric test of differences among groups (Mielke, 1984; McCune and Grace, 2002). MRPP yields A values, a statistic that describes effect size. When within-group heterogeneity equals that expected by chance, A = 0; when sample units within groups are identical, A = 1. The p value is for δ, a measure of within-group distance, which is used to calculate A (McCune and Grace, 2002). For this test, rank-transformed Sørenson distances between plots were used with the recommended weights. MRPP was also used to determine if plot scores for each axis from the NMS ordination differed among classes of the categorical variables Topographic Class and USDA Soil Texture. For these MRPPs, plot scores on axes were adjusted using relative Euclidean distances, with the recommended weights on distance (Peck, 2016).
The association between quantitative environmental variables per plot and the categorial variables of Topographic Class and USDA Soil Texture (Tables 1 and 2) were also evaluated using MRPP. The quantitative variables were relativized as described for the Mantel test, and Sørenson distances were used with the recommended weights.
Individual species analysis
We determined the Pearson correlations (R2 >10%) between individual tree species abundances per plot and each of the three axes of the NMS ordination, as stated above for quantitative environmental variables.
To evaluate the association of individual tree species with the categorical variables Topographic Class and USDA Soil Texture, we used Indicator Species Analysis, based on relative abundance and constancy (Peck, 2016). Constancy is the proportion of sample units in each class in which a species occurs. Indicator values (IVs) were calculated for each species for each categorical variable class (Peck 2016) to yield a maximum IV (IVmax) among those classes (Dufrêne and Legendre, 1997). The significance of the IVmax for each species was determined with a Monte Carlo randomization test. The significance of the sum of all species IV (Sum IVmax) for a given categorical variable was also determined with a Monte Carlo test.
Results
Tree species composition among plots
The 69 tree species (stems ≥10 cm DBH and present in ≥5 of the 209 plots) used in NMS ordination tree plots in the Rio Bravo Conservation and Management Area, Belize.
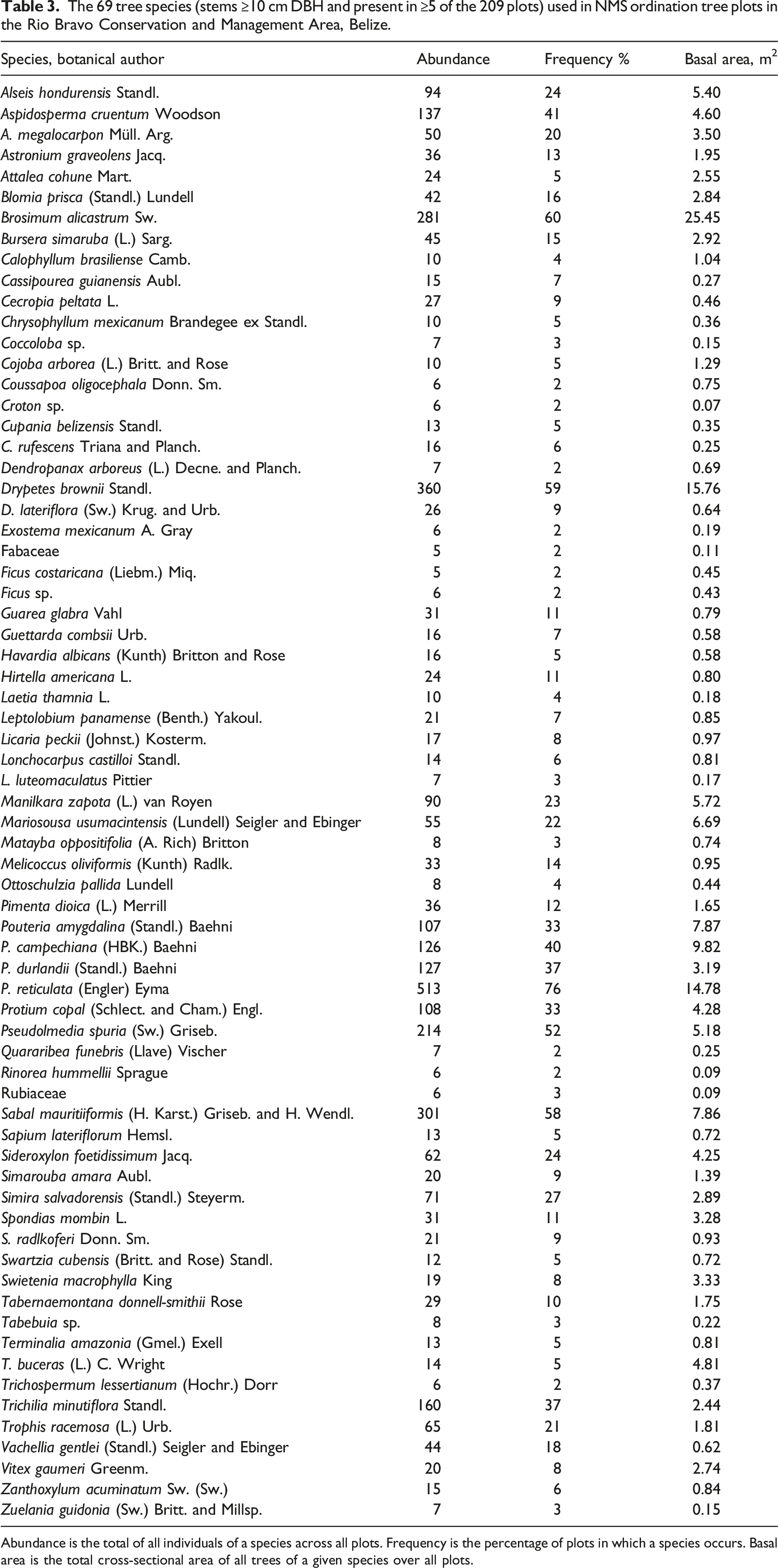
Abundance is the total of all individuals of a species across all plots. Frequency is the percentage of plots in which a species occurs. Basal area is the total cross-sectional area of all trees of a given species over all plots.
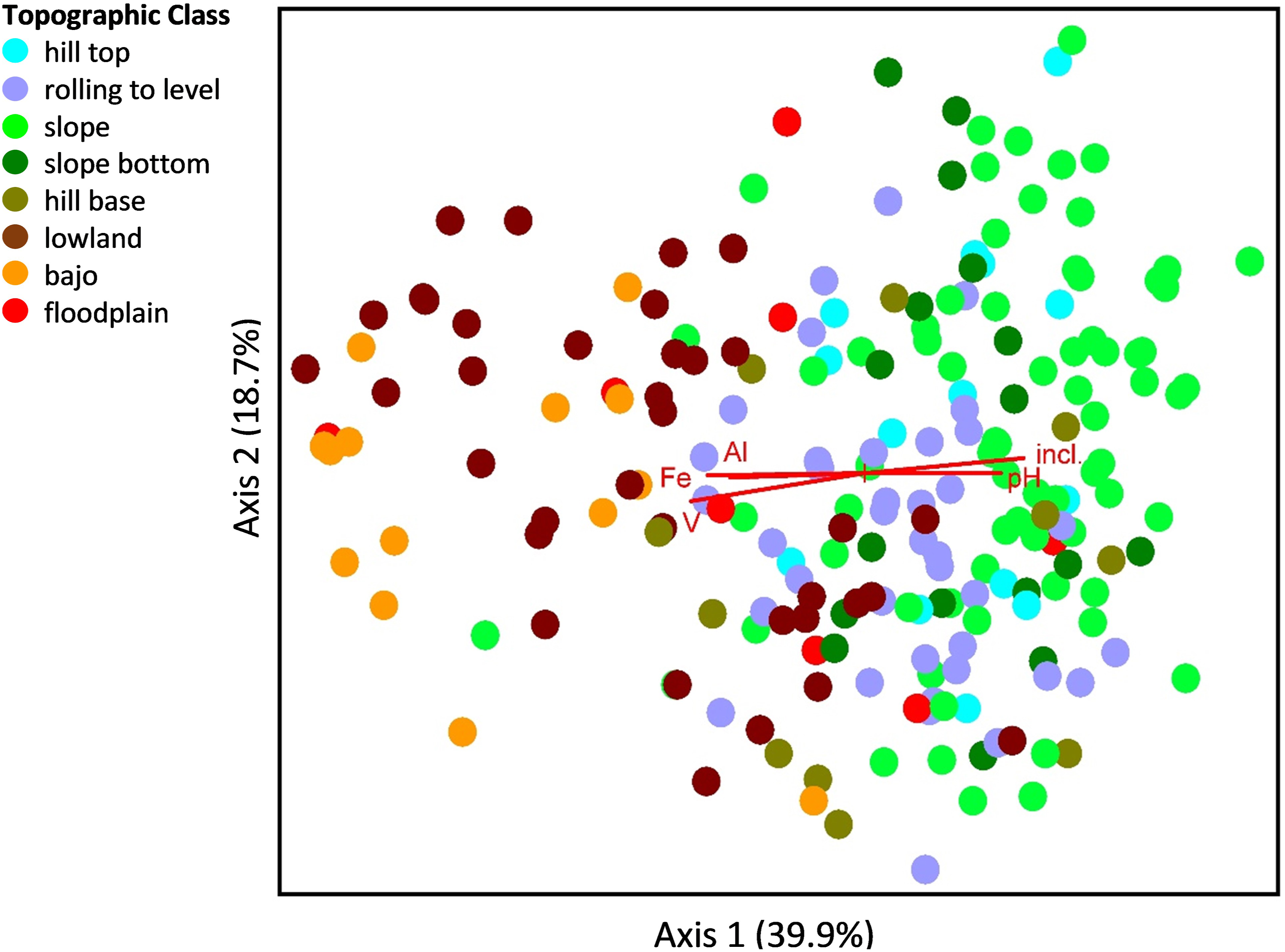
Ordination of tree species composition in 209, 400-m2 plots in Belize. Each plot is shown as a dot. The dots are colored according to their Topographic Class and arranged according to their similarity in tree species composition along two axes. A gradient in species composition along Axis 1 is evidently related to the topographic gradient, from lowland and bajo on the left to hilltop and slope on the right. The red lines show the quantitative variables pH, V (vanadium), Al (aluminum), Fe (iron), and degrees inclination (incl.) that were highly correlated with Axis 1 (R2 > 0.25). The percentage of variation accounted for by each axis is shown in parentheses on the axis label.
Species composition and quantitative variables
Several tests indicated strong relationships between species composition and quantitative environmental variables. In the NMS ordination, the pattern of species abundances was significantly related to the pattern of quantitative environmental variables (Standardized Mantel Statistic: r = 0.254, p for Type I error = 0.001). Other Mantel tests for the three different NMS axes showed that the matrix of relativized, quantitative environmental variables was most strongly associated with NMS plot scores on Axis 1 (Axis 1: Standardized Mantel Statistic: r = 0.184, p for Type I error = 0.001) and less so for Axis 2 (r = 0.0600, p = 0.001) and Axis 3 (r = 0.020, p = 0.017).
Pearson correlations (R2 > 0.10) of NMS axes with relativized nutrient values, degrees inclination, and soil particle composition.
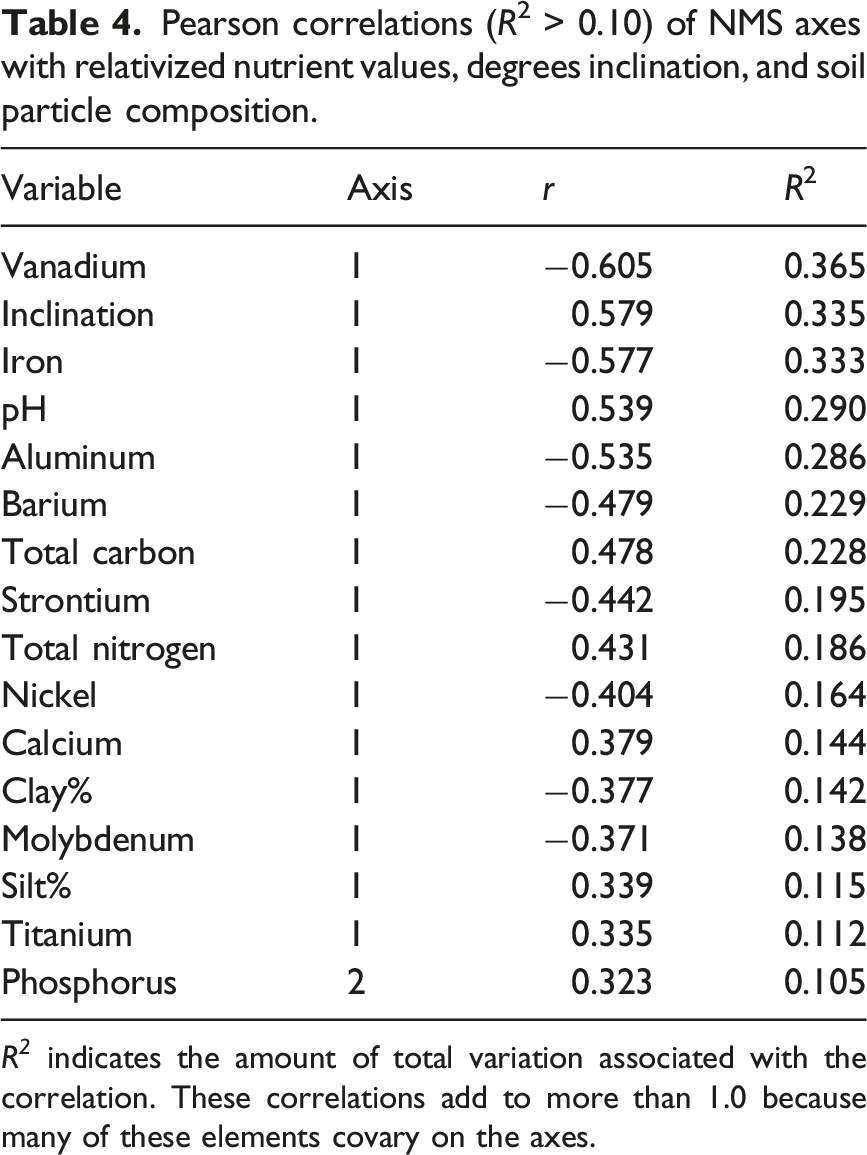
R 2 indicates the amount of total variation associated with the correlation. These correlations add to more than 1.0 because many of these elements covary on the axes.
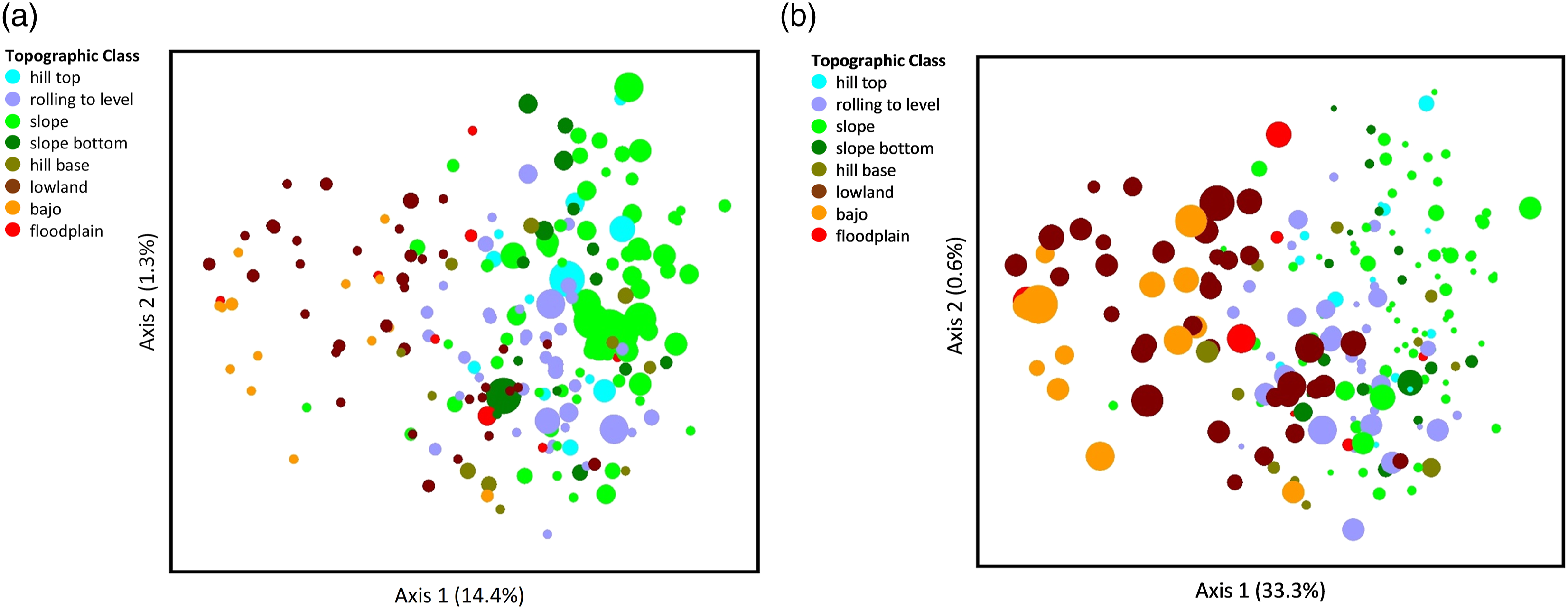
Ordination of soil elements: (a) calcium and (b) iron. The graphs depict the same ordination of plots arranged according to similarity in species composition as in Figure 2, with dots (study plots) colored according to Topographic Class. But the dots here are sized to indicate the relativized value of soil characteristics (larger dots = larger values). The amount of variation accounted for by each axis is shown as a percentage in parentheses on the axis label.
Species composition and categorical variables
MRPP results for association of Axes 1, 2, and 3 of the NMS ordination with categorial variables. (For explanation of the test statistic A, see Methods.)

Numbers in bold are significant at α ≤ 0.05.
Individual species
Twenty-one species whose abundances correlated (R2 > 10%) with one or more axes of NMS ordination.
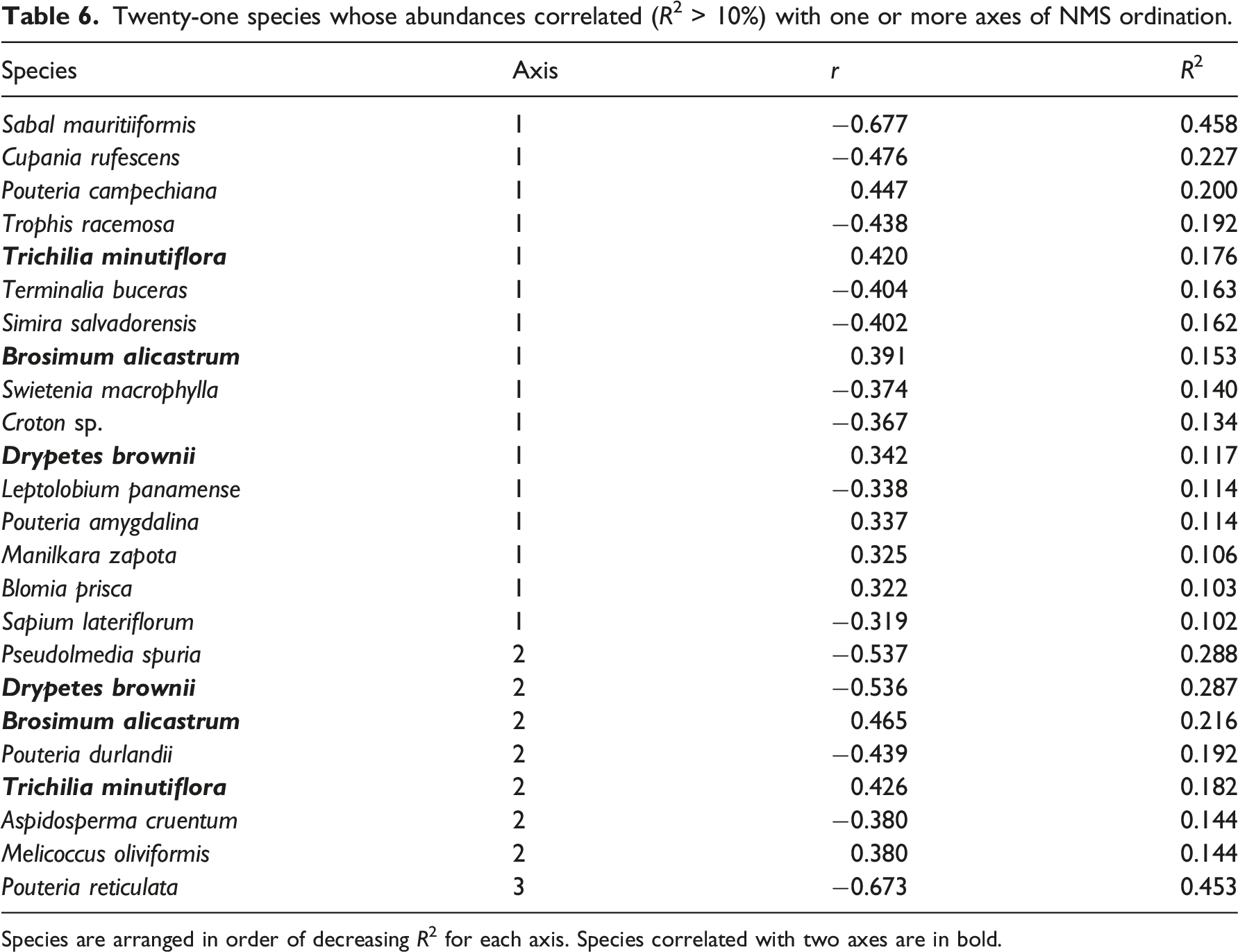
Species are arranged in order of decreasing R2 for each axis. Species correlated with two axes are in bold.
Indicator species: Tree species associated with a particular Topographic Class or with USDA Soil Texture.
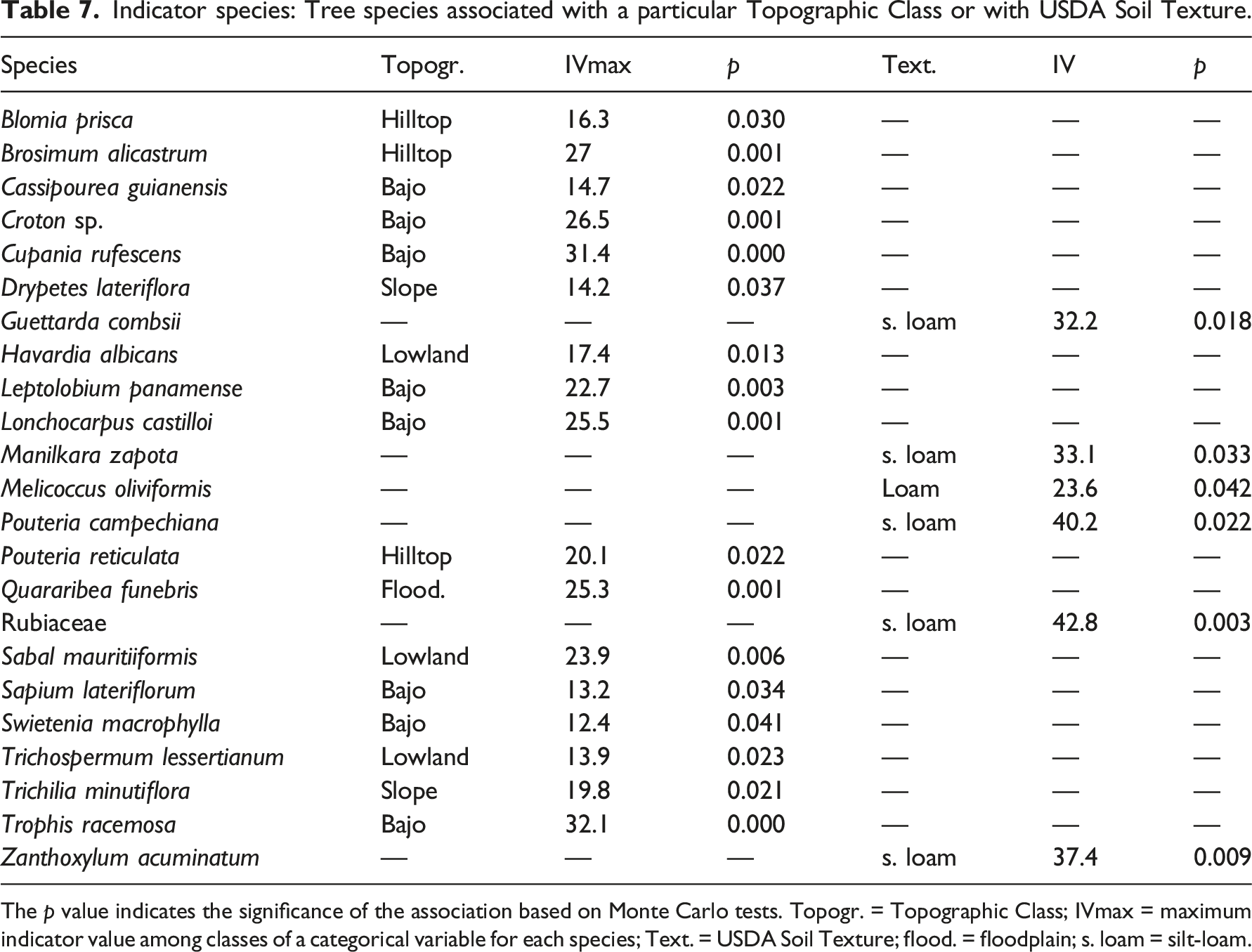
The p value indicates the significance of the association based on Monte Carlo tests. Topogr. = Topographic Class; IVmax = maximum indicator value among classes of a categorical variable for each species; Text. = USDA Soil Texture; flood. = floodplain; s. loam = silt-loam.
Three abundant species were chosen to portray contrasting patterns in the ordination (Figure 4). Brosimum alicastrum is an indicator most abundant in sites high on the topographic gradient (Figure 4(a); Sabal mauritiiformis, a palm, is an indicator more abundant lower on the gradient (Figure 4(b)); and Aspidosperma megalocarpon is not an indicator species and is more broadly distributed on the gradient (Figure 4(c)). The axes explain a substantial amount of the variation of the indicator species B. alicastrum and S. mauritiiformis but explain little of the generally distributed A. megalocarpon. Representation in ordination space of (a) Brosimum alicastrum, (b) Sabal mauritiiformis, and (c) Aspidosperma megalocarpon. The graphs depict the same ordination of plots arranged according to similarity in species composition as in Figure 2, with dots (study plots) colored according to Topographic Class. But the dots here are sized to indicate the relative abundance of three tree species in each study plot (B. alicastrum, smallest to largest dots = 0–7 individuals per plot; S. mauritiiformis, 0–9 individuals per plot; A. megalocarpon, 0–3 individuals per plot). The amount of variation accounted for by each axis is shown as a percentage in parentheses on the axis label.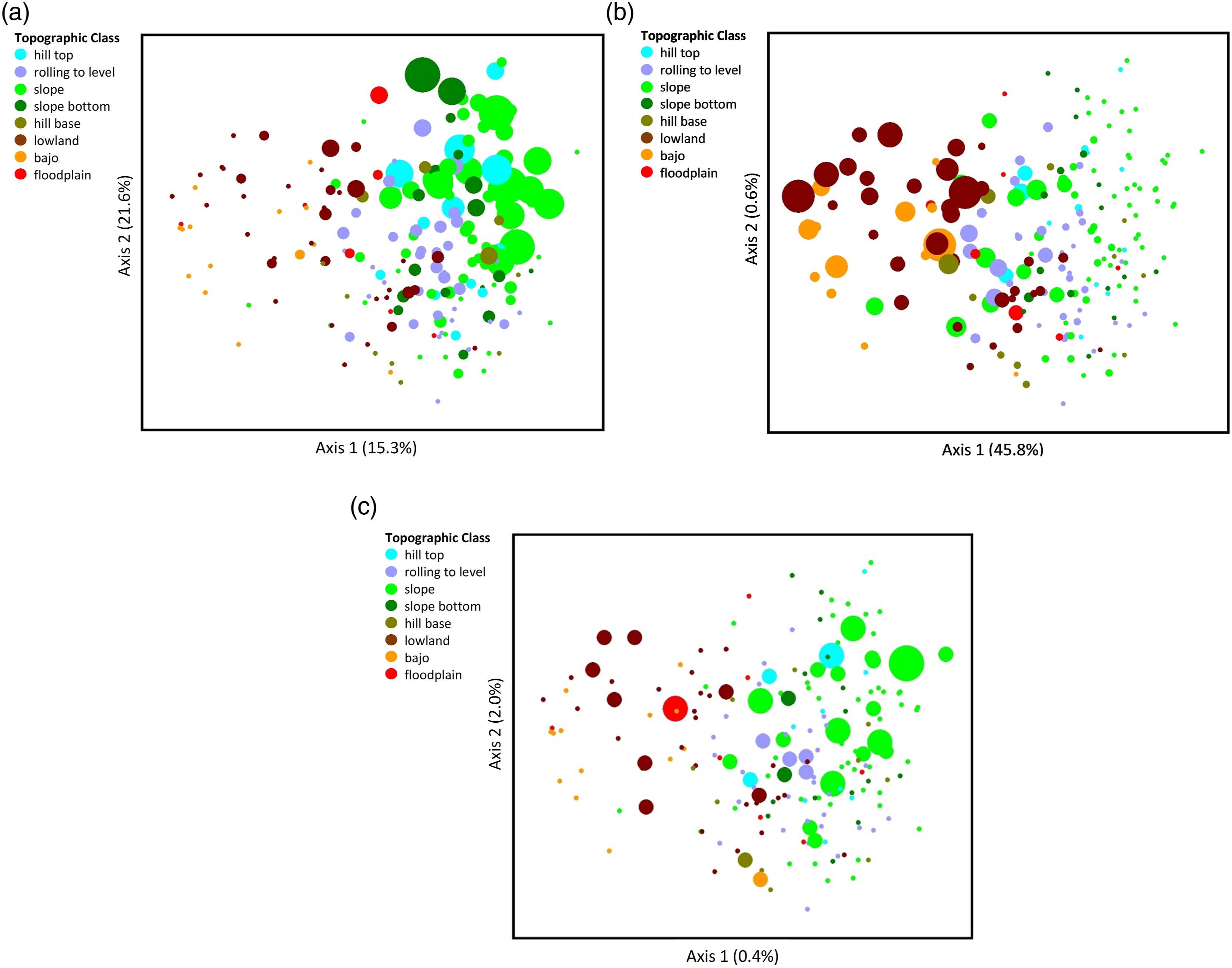
Discussion
More than 60 years ago Charles Wright described “… the remarkable correlation between the ecology of the natural vegetation on the one hand and the soil pattern on the other [in Belize]” (Wright et al., 1959). Our study supports this generalization. We found that tree species composition and the spatial distributions of many individual tree species were significantly associated with quantitative soil variables and Topographic Class in northwest Belize. Twenty-one tree species, representing 68% of all individual trees sampled in our 209 plots, exhibited consistent, non-random distributions along the three axes of the NMS ordination. Twenty-three Indicator Species were significantly associated with the categorical variables Topographic Class and USDA Soil Texture.
Mantel and MRPP tests indicated that plot species composition was strongly associated with patterns in quantitative and categorical environmental variables, both for the overall NMS analysis and for individual NMS axes. Of the variation in tree species abundances among plots, 39.9% was accounted for by Axis 1 from the NMS ordination. This axis was correlated at R 2 > 10% with 15 of the 34 quantitative variables we measured, the most important variables being pH, vanadium, aluminum, iron, and degrees inclination. Axis 2 accounted for 18.7% and was correlated only with phosphorus. But phosphorus is important; it limits productivity in many tropical forests (Hou et al., 2020), and its variation across the landscape could affect species composition and distribution. Axis 3 accounted for 14.5% of variation in tree abundances, and no environmental variable was correlated with it. Overall, 27% of the variation in species composition among plots was not accounted for by the NMS ordination. This remaining variation could be related to unmeasured environmental variables, small sample size of many uncommon species, stochastic effects, or unstudied biological traits and interactions.
The categorical variable, Topographic Class, was strongly associated with species composition on Axis 1. The strength of this variable reflects the fact that position along a topographic gradient is consistently associated with trends in soil moisture, depth, nutrient content, and other environmental variables (Furley and Newey, 1979). In our study, higher topographic positions had greater degrees inclination, pH, total carbon, total nitrogen, calcium, silt%, and titanium (most of these important for plant growth), and lower amounts of various elements and clay%, than did lower topographic positions (Figure 2, Table 4). Thus, the variable Topographic Class integrates different variables that are likely associated with species composition. In our study, the association of Topographic Class with species was stronger than the association of USDA Soil Texture with species. However, the associations of Topographic Class and USDA Soil Texture with quantitative soil variables were more similar. This may partly reflect the impact of soil particle composition on element or nutrient retention (Leinweber et al., 1995).
The distributions of 16 individual tree species were associated Axis 1, which in turn was associated with degrees inclination and 14 quantitative soil variables. Pouteria reticulata, the most common species on the plots, was the only species correlated with Axis 3 and had the strongest association of any species with an axis. Yet no measured environmental variable was correlated with Axis 3, suggesting that P. reticulata abundance reflects some unmeasured variable. Three species (Drypetes brownii, Brosimum alicastrum, and Trichilia minutiflora) were strongly correlated with both Axes 1 and 2, suggesting multiple environmental influences on their distributions.
Consistent with the strong association of species composition with Topographic Class, there were 17 Indicator Species associated with different Topographic Classes. Indicator Species tended to be located at the ends of the topographic gradient: in the hilltop class at the upper end (Brosimum alicastrum and Pouteria reticulata) and in bajo or lowland classes at the lower end (Havardia albicans and Sabal mauritiiformis) (Table 3). These species seem to specialize in, or are restricted by competition to, the presumably extreme conditions found at topographic end points. The six Indicator Species for USDA Soil Texture classes were not associated with any Topographic Class.
Studies of tree species composition in the Maya Lowlands using multivariate analysis of both tree and environmental data.
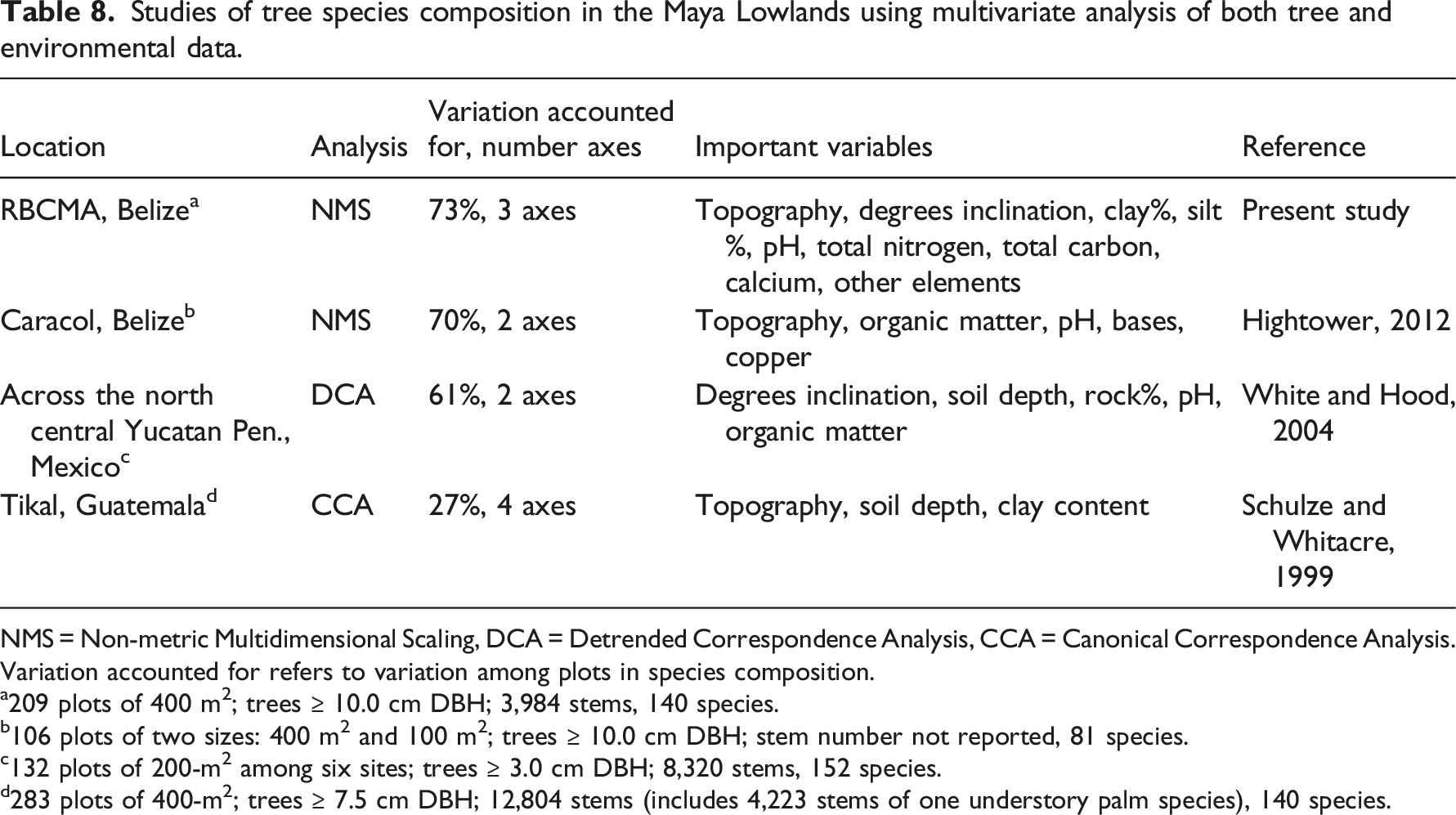
NMS = Non-metric Multidimensional Scaling, DCA = Detrended Correspondence Analysis, CCA = Canonical Correspondence Analysis. Variation accounted for refers to variation among plots in species composition.
a209 plots of 400 m2; trees ≥ 10.0 cm DBH; 3,984 stems, 140 species.
b106 plots of two sizes: 400 m2 and 100 m2; trees ≥ 10.0 cm DBH; stem number not reported, 81 species.
c132 plots of 200-m2 among six sites; trees ≥ 3.0 cm DBH; 8,320 stems, 152 species.
d283 plots of 400-m2; trees ≥ 7.5 cm DBH; 12,804 stems (includes 4,223 stems of one understory palm species), 140 species.
These studies (Table 8) are foundational for future research on factors that control tree species composition. So far, these studies suggest, but do not prove, that soil characteristics affect tree species distributions. To clearly determine causes of distribution, we need experiments. For example, in forests of Borneo (Born et al., 2014; Palmiotto et al., 2004) and Panama (Brenes-Arguedas et al., 2008; Kitajima, 1994), controlled field and growing-house experiments on tree seedlings revealed species-specific growth responses to variation in light, water, or nutrients that help explain species-specific distributions of adult trees in the forest. For other future studies, multivariate analysis of species and the environment can be coupled with measurements of species traits, such as “specific leaf area” and growth rate, to better understand the causes of species distributions and abundances (Meyer and Seifan, 2019).
Although we think our study sites have been free of significant human impacts for c. 1000 years, the ancient Maya were present in our area (Scarborough et al., 2003), and researchers have shown local effects of ancient Maya land use on modern tree species composition at comparable sites (Hightower, 2012; Ross, 2011; see Supplemental Material regarding a study on the Maya legacy in the RBCMA but lacking adequate design). Such an effect of ancient Maya land use would reinforce our results showing predictable tree composition, if we assume the ancient Maya farmed consistently according to topography and soil and this had persistent effects on modern tree distributions. For instance, ancient farming was closely adapted to local soil types in the Puuc region of Mexico and the Pasión region of Guatemala (Dunning, 1996), and the modern Maya of Motul de San José in Guatemala apply a detailed soil classification to their agriculture (Jensen et al., 2007). Not inconsistent with local effects of ancient land use, two studies at Tikal, both covering large areas, concluded that modern spatial distributions of tree species were largely explained by current environmental conditions (Schulze and Whitacre, 1999; Thompson et al., 2015). Either way, our study shows how predictable the effects of topography and soil are for vegetation, whether it be modern forest or ancient crops. It reveals a basis for the informed land use—a “managed mosaic” of local soils and plants—attributed to the ancient Maya (Fedick, 1996; Ford and Nigh, 2015) and which some suggest strongly contributed to their success (Demarest, 2004). Also, the predictable variation in the distributions of numerous tree species versus micro-environmental variables indicates habitat specialization, which can partly explain the richness of species in this region (e.g., Paoli et al., 2006).
While our study of the modern forest helps us visualize the environment of the ancient Maya, our quantitative results can also help us visualize a future forest shaped by climate change. “Species distribution modeling” (Franklin, 2009) can use known correlations between species distributions and environmental variables to predict future distributions under environmental change. For example, a drier climate is predicted for Central America (Hidalgo et al., 2013). This could exaggerate the comparative lack of soil water on upper slopes (García-Gamero et al., 2021), which, in turn, can affect nutrient availability (Marschner and Rengel, 2012; Viets, 1967). In particular, soil water is known to affect the availability of the key element phosphorus (O’Connell et al., 2018), and therefore possibly the distributions of species sensitive to phosphorus, along the topographic gradient. Modeling future species distributions, based on results like ours that reveal the strength of environmental associations with species distributions, can be a basis for planning agriculture and conservation.
Supplemental Material
Supplemental Material - Tree species on environmental gradients in subtropical forest of northwest Belize, in the Maya Lowlands
Supplemental Material for Tree species on environmental gradients in subtropical forest of northwest Belize, in the Maya Lowlands by Sheila E Ward, Nicholas Brokaw, Stanley Walling, and Marisol Cortes-Rincon in Progress in Physical Geography: Earth and Environment
Footnotes
Acknowledgements
Fieldwork in Belize was undertaken under a permit issued to Fred Valdez for the Programme for Belize Archaeological Project (PfBAP) by the Institute of Archaeology of the Belize National Institute of Culture and History. The Rio Bravo Conservation and Management Area (RBCMA) is managed by the Programme for Belize (PfB), a Belizean organization dedicated to tropical forest conservation, sustainable management, and environmental education. We thank PfB for permission to do research in the RBCMA and for logistical support. The PfBAP (director Fred Valdez) has been the base for our field work. Many people have helped us in the field. The authors would like to acknowledge the field directors, student interns, and volunteers of the Rio Bravo Archaeological Survey under the direction of Stanley Walling, and of the Dos Hombres-Gran Cacao Transect project directed by Marisol Cortes-Rincon, and student interns from the University of Puerto Rico and the College of St. Benedict, all of whom have contributed to this research over a number of research seasons. Thanks also go to the able field workers from the community of San Felipe, Belize, who have assisted in excavation, survey and clearing operations. We thank Elizabeth Mallory for the base map in Figure 1 and unnamed reviewers for suggesting substantial improvements to the manuscript.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: the United States National Science Foundation (DEB-1114947), United States Department of Agriculture (2017-38422-27131), and a University of Puerto Rico-Río Piedras Small Summer Grant. The research was also partly funded by a Google Arts and Heritage Grant, National Geographic Society Grant 8266-07, the Montclair State University Foundation, The Community College of Philadelphia, and the Belize Archaeology Annual Field School of Humboldt State University.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.