
Abstract
As environmental change and mass extinctions underline an urgent need to establish more humane relationships with non-human beings, there is a creative opportunity to reimagine concepts of kinship to promote the collective well-being of all living kinds. Anthropology draws on culturally diverse interspecies relations: some locate human and other species within distinctive and hierarchical categories, while others have more fluid and egalitarian notions of personhood. Engagements with non-human species therefore range from objectifying and exploiting them, to their acceptance as kin, as persons, and as reciprocal co-creative partners in the composition of shared lifeworlds. Though the concept of kinship is conventionally used to illuminate inter-human relations, this article suggests that it has further potential to raise key questions about how societies engage with non-human beings, and our ethical responsibilities towards them. These questions might usefully inform contemporary debates about non-human rights, and how these might be upheld by state and/or international legislation.
Introduction
Extinction rates for non-human beings are now running at about 200 species a day and are still accelerating. As human populations expand and continue to intensify their use of material resources, local, regional and planetary ecosystems are being heavily impacted. Anthropogenically caused climate change is promising a storm that, to paraphrase James Hansen (2011), will be visited upon our grandchildren. There is a clear need for paradigmatic changes to human relationships with non-human beings. A key question for anthropologists, therefore, is whether we can employ ideas about kinship to challenge the anthropocentricity that permits societies to externalise the costs of unsustainable lifeways to other species. This article considers the approaches within anthropology and related disciplines that might be brought together to compose narratives of engagement that move beyond the confines of human kinship to a vision of organic and social connection that encompasses all living kinds and protects our collective future. It aims to conceptualise a more convivial and mutually reciprocal relationship with the non-human domain, which we might describe as ‘living kindness’.
Anthropology has a long-term interest in human–non-human relations. Challenging conventional categories, Lévi-Strauss (1964) suggested that speciesism and racism sat alongside each other, with notions of human exceptionality strengthening the internal speciesism of racism. Viveiros de Castro (1992) describes ways of conceptualising human–non-human relations in ways that eschew dualistic categories of culture/nature or human/other. Also in South America, Descola (2017) notes Achuar concepts in which subjectivity is seen as an experience shared by humans and non-humans alike. There are, in fact, multiple alternate ontological models in which living kinds inhabit indivisible lifeworlds and nature is not ‘othered’ (e.g. Deimel and Ruhnau, 2000; Fienup-Riordan, 2017; Kohn, 2015; Morphy, 1991; Strang, 2005a).
Thus the ‘ontological turn’ in anthropology and philosophy has moved beyond a comparison of cultural perspectives on an implicitly normative reality to recognising that diverse lifeworlds are created in relatively autonomous and dynamic processes of being and becoming (Charbonnier et al., 2017). While embracing this diversity, my own research on human engagements with water has been influenced by consistent evidence that the meanings encoded in water recur cross-culturally because of the shared essentiality of water in human lives; the commonalities of human sensory experience; and water’s consistent material properties (Strang, 2005b). A similar point could be made about cultural diversity and underlying commonality in societal concepts of kinship, following Sahlins’ (2013: 28) cross-cultural vision of kinship as ‘mutuality of being’, and Strathern’s (2020a) lead on a broader concept of ‘relations’ and its potential for comparison.
Thus acknowledging cross-cultural recurrences does not preclude cultural and ontological diversity in human–water relations any more than it does with ideas about who or what constitutes kin. It merely highlights the potential for balancing a comparative view of shared human experiences with an appreciation that every cultural (and individual) experience is unique, and formed by a particular location in time and place. This ‘have your cake and eat it’ approach is, I suppose, an unapologetic vote of confidence in the comparative advantages of anthropology, which enables cross-cultural consideration of concepts such as kinship, or of human engagements with a shared material world, while simultaneously valorising the cultural and historical diversity that permits comparison.
Totemic kinds
In challenging conventional speciesism, Lévi-Strauss’s writing about totemic ancestry (1964) encouraged further work on notions of kinship between human and non-human beings. But anthropology’s interest in totemic relations goes back further. The term ‘totem’ itself is based on an Algonquian/Ojibwa word for ‘brother-sister kin’ (Figure 1), and was foundationally defined by James Frazer as: an intimate relation which is supposed to exist between a group of kindred people on the one side and a species of natural or artificial objects on the other side, which objects are called the totems of the human group. (1910: 8) Totem pole, Stanley Park, Vancouver. Source: Photo Wikimedia Commons.
This usefully reminds us that totemism is not merely a feature of the place-based societies that have retained long-term cosmological belief systems, but also appears in the pre-history of many larger societies, whose religious ideas subsequently shifted towards the worship of more humanised deities (Strang, 2023). Thus Jane Harrison (2010: 119), in her seminal work about the totemic origins of Greek religion, points to the non-human nature of earlier divinities and the practice of omophagia, the eating of totemic beings, which both absorbed them into the human body and expressed sacramental communion with them. First and foremost, then, in totemism, is the idea of the unity of a group…. This relation between the human and non-human group is so close as to be best figured by kinship, by unity of blood, and is expressed in terms of actual identity. (Harrison, 2010: 120–1)
As she points out, such co-identification expresses an equitable relationship of mutual interdependency. Totemic ceremonies are ‘intensely sympathetic and cooperative; they are, as the Greeks would say, rather methektic than mimetic, the expression, the utterance of a common nature participated in, rather than the imitation of alien characteristics’ (2010: 125).
A vision of a ‘common nature’ appears in many stories of cosmogenesis. These necessarily explain how things and persons come into being, and in multiple cosmological schemes non-human ancestral beings feature as progenitors for human descendants. Given that all societies understand that water is essential to life, it is unsurprising to find that many origin stories describe a world-creating being ‘taking form’ out of fluid chaos, or emerging from primal waters. Critically, in kinship terms, such tales often involve parthenogenesis, in which material worlds and all living kinds are made out of the bodies of primal ancestors. My interest in homologues (Strang, 1999, 2017) leads me to agree with Lincoln (1986: 3), that origin stories employ scheme transfers in which living kinds, the world and the cosmos are alloforms of each other, and thus share a ‘fundamental consubstantiality’.
Thus ancient Mesopotamians described a great serpent arising from a watery primordial abyss, and the world being made from the two halves of its body (Dalley, 2008). In the Mayan cosmos, all living kinds emerge from an aquatic creative being, Itzam Na (Figure 2), and two celestial serpent beings dismember a third to form the world (Ferguson, 2000). In early Chinese origin stories the body of Pan Gu becomes each part of the world (Chen and Yang, 1995). Norse sagas describe how the creator Ymir begets progeny and forms the world out of its own body (Sigfusson 1866). In these ways, emerging from water (the element literally essential to all living organisms), creative beings establish the common ancestry of living kinds. Itzam Na, the Mayan primal ancestor being, Dresden Codex. Source: Photo Wikimedia Commons.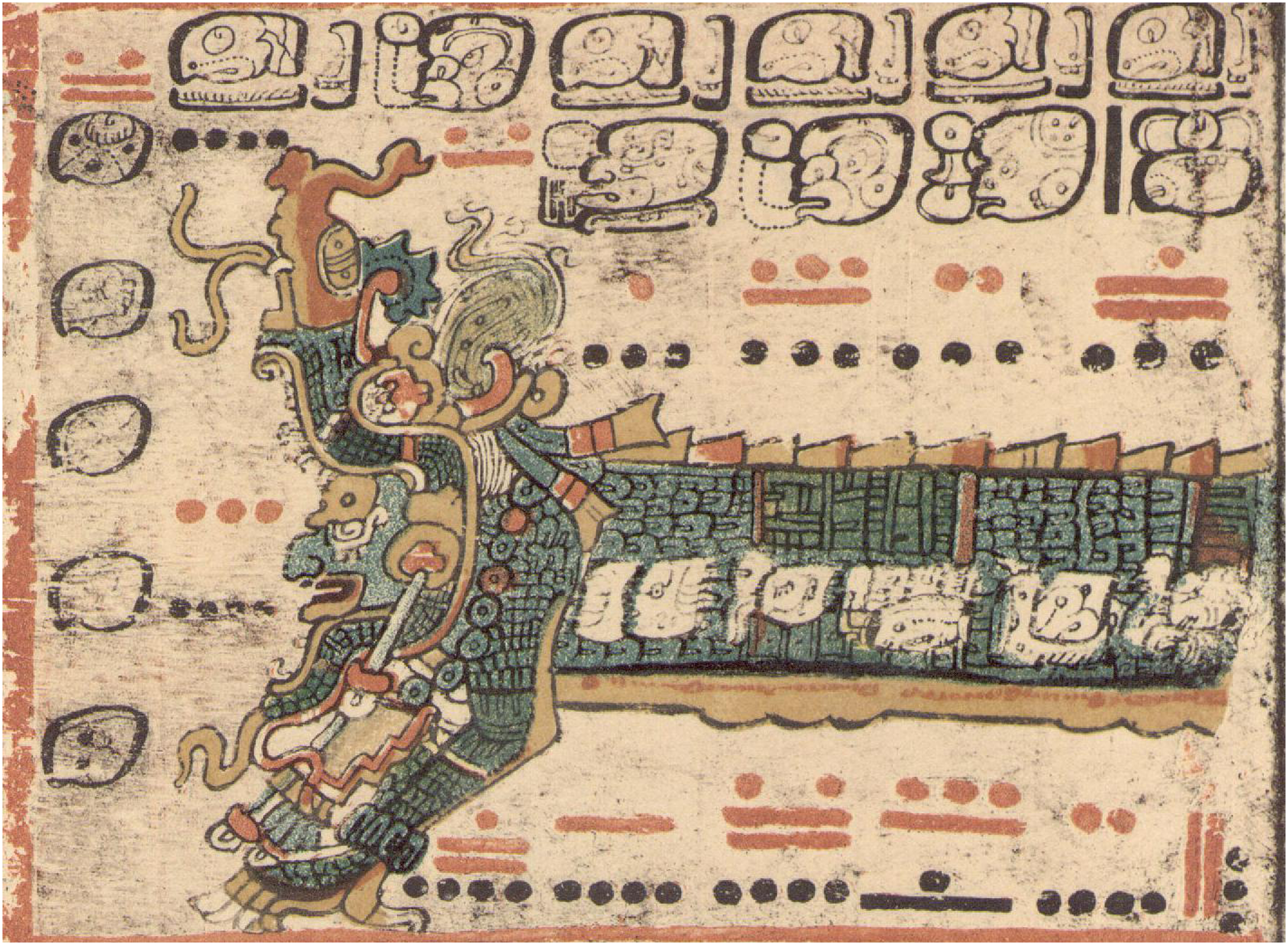
A powerful vision of consubstantiality is provided by indigenous beliefs in Australia, passed down the generations through songs, stories, art and ritual. Central to Aboriginal visions of cosmogenesis is the Dreaming or Story Time, in which totemic ancestors emerged from the Rainbow Serpent, which manifests the generative powers of water (Figure 3). Living kinds were spewed or spat out of the serpent, to ‘take form’ in the visible, material world. Exemplifying the customary lifeways of hunter-gatherers, the ancestral beings engaged with and formed the land and waterscape. In Far North Queensland, for example, Kunjen elders in Kowanyama relate a flood story that (like other flood stories around Australia’s northern coast) may recall sea level rises that filled the Gulf of Carpentaria 10,000 years ago. The Two Brothers, Antujil, chopped down a giant tree. As they did so they sang patens (sacred songs) and the wood chips flew everywhere. Every time a chip flew, they named it, making all of the different trees in the world. They took the biggest branches from the tree and threw them to the west, forming the coastal sand ridges that prevent the sea from flowing across Cape York. They put two poles up to stop that water from coming, overflowing the country, so that land and the sea go one side and lagoon separate. When big rain came they still here. Sink it down, just like a big yam stick, you know – with a sharp end…. They stopped the sea coming this way – they put all them tree along that hill country…. Stopped that sea. (Lefty Yam, in Strang, 1994) Rock art in Chillagoe, far North Queensland. Source: Photo Veronica Strang.
But the Dreaming is more than a creative era. It is a non-material dimension from which life continues to flow in and out of the visible material world. The Rainbow Serpent remained in the land, and – especially when encouraged by increase rituals – continues to generate all forms of life. This includes humans: every individual has a totemic home, usually a water source within their clan estate, from which the Rainbow Serpent generates their spirit. This ‘jumps up’ into their mother’s womb, moving from the non-material domain into the visible world and a material form. When a person dies it is to this watery home that their spirit must be returned via songs and rituals, so that it can be reunited with its totemic ‘mate’, the ancestral being located in that home place. This totemic connection locates them spatially and socially in a clan and kin group, and provides them with rights and obligations in relation to clan country (Strang, 1997).
These cosmological beliefs therefore envision both material and social kinship. Humankind emerges from a primal ancestor who, as in many cultural contexts, continues to create not just human persons, but all living kinds. Underlining this consubstantiality, Aboriginal concepts of materiality observe that human beings are literally composed of the food and water they consume and of the places that ‘grow them up’. There is frequent recurrence, cross-culturally, in ideas about how relations are both socially and materially emergent. For example, observing that ‘identity and substance are mutable’, Carsten (1995: 223) offers a similarly processual view of kinship, as conceptualised in Malay households. And there is accord between the social and material sciences here: like all living organisms, the human body is a constantly developing micro-ecosystem reliant upon water, nutrients and other materials flowing in an orderly fashion between itself and its environment.
Material ancestry
A leap across disciplinary divides to consider how ancestral connections are explicated in the material sciences offers a complementary evolutionary vision of life ‘taking form’ out of primal oceans. Life on Earth began in ancient seas 4 billion years ago, when water and light interacted to enable amino acids, carbon monoxide, carbon dioxide, nitrogen and other organic matter to form living cells with metabolic and reproductive capacities. Then, about 2 billion years ago, photosynthesis created an atmosphere supporting more sophisticated multicellular organisms.
All species remained aquatic in nature until relatively recently when – about 450 million years ago – they emerged onto terra firma. Since then, even as they diverged (and some evolved into bipedal primates), all species, without exception, remained dependent upon water flowing through and irrigating their bodies. Thus all of the organic beings composing terrestrial life remain united by a ‘hypersea’ of water which flows between them, connecting them to each other, and to the material world (McMenamin and McMenamin, 1994). Water is similarly central to more evanescent connections, being necessary to all forms of consciousness. Neurological activity relies on water molecules to carry the electrochemical transmissions that literally spark our thoughts and, we can reasonably suppose, similarly enables the varied forms of consciousness of other species living in the hypersea (Kohn, 2013; Tsing, 2017).
Water is also the major constituent of blood, which is seen in many cultural contexts as one of the most essential material ‘substances’ of kinship. In this sense, the hypersea of water circulating between living kinds might be conceptualised as a larger, albeit ‘watered down’, material element of kinship, in which differences between species are not degrees of separation but merely a matter of distance (or a distance of matter). Evolutionary science offers other interspecies connections: developmental trajectories leading to similar skeletal structures, circulatory systems and the composition of flesh. Crucial to both similarities and differences is the self-replicating DNA that passes on hereditary traits from one generation to the next. 1 , 2 Pan-species comparison, based on shared DNA, reveals the close and more distant relationships between species that have emerged over evolutionary time.
Like giant kinship diagrams, evolutionary relationships can be traced to common ancestors, with each generation dividing and diversifying (Figure 4). Humans contain about 3 billion genetic building blocks, and less than .01% of these genes differentiate us from other human beings. Our closest primate relatives, chimpanzees and bonobos, with a common ancestor a mere 4–5 million years in the past, share 98.7% of our DNA. Domestic cats share about 90%, and mice about 85%. With cattle it is about 80%; with chickens and insects about 61%. Our common organic heritage with plants, though more temporally distant, is still readily discernible, in that even now there is a 50% DNA overlap between humans and bananas. This allows us all to indulge in a comment that some of our relatives are bananas. Darwin’s evolutionary tree. Source: Photo Wikimedia Commons.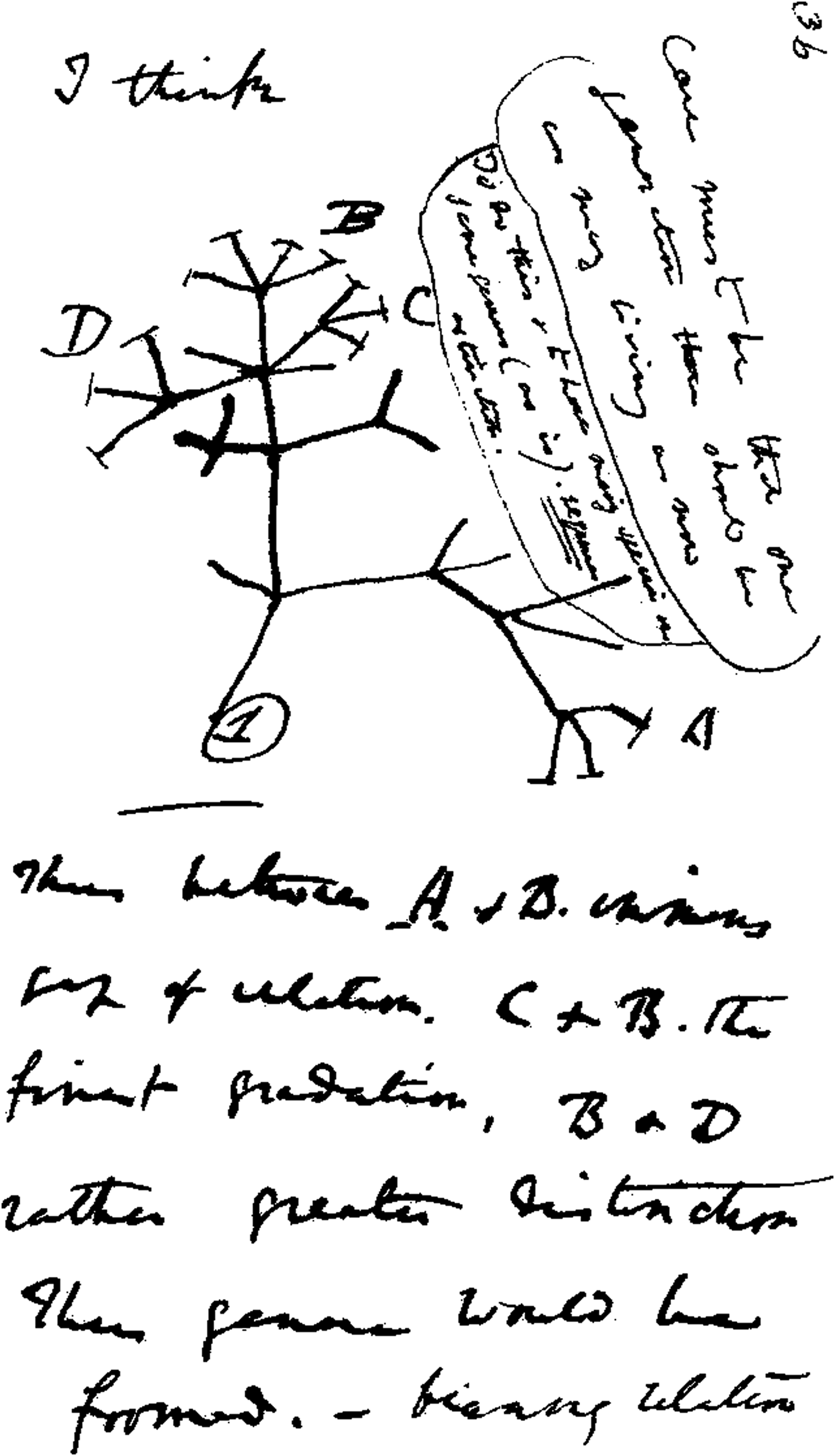
DNA brings home the idea that the infinite range of potential evolutionary variations is nevertheless composed of, and generated within a shared material environment. These variations are readily comparable: for example, the evolution of vision reflects particular kinds of engagement with the world. Pollinating insects, who collect food from flowers, have highly developed colour vision. Cats, as nocturnal predators, have little colour vision but great capacity to see in dim conditions and to spot the slightest movement. Humans, omnivorous in depending primarily on fruits and berries, but also upon hunting, have vision that is somewhere in between, with a fairly good discernment of colour, as well as capacities to pick up movement.
An evolutionary perspective therefore casts a somewhat different light on anthropological understandings of totemic ancestry. Its internal logic turns out to be compatible with indigenous Australian thinking, that a human clan may have a non-human totemic ancestor, who emerged from a primal water being. No doubt similar resonances can be found between evolutionary ideas and visions of cosmogenesis in other cultural contexts. While radically different ontologically, all such views envision humankind ‘becoming’ out of co-creative organic and material systems. For example, Māori iwis describe their local rivers as ‘living ancestors’. Their ancestral progenitors, Papatūānuku the earth mother and Ranginui the sky father, beget other Atuas (divinities) representing different aspects of the material world, such as Tangaroa the god of the sea. These Atuas in turn produce human progeny (Barlow, 1991; Muru-Lanning, 2016). However, there is a key difference between beliefs in ancestral totemic beings and worldviews conforming to more Cartesian visions of evolution, and that is the extent to which the non-human domain is seen as an active and conscious partner in events.
Living kinds
There is a rich anthropological literature on diverse ways of thinking about what constitutes being alive, animate or sentient. Here too, there are ancestral connections between stories of cosmogenesis. Typically, primary ancestors are also the source of consciousness and enlightenment, endowing humankind with the wisdom encapsulated in concepts of religious morality and social ethics. Aboriginal Australians refer to this as ‘the Law’ and, like everything else, this is generated by the Rainbow Serpent, who (along with other totemic ancestors) remains immanent as a sentient, watching, nurturing and guiding presence in human lives.
The concept of ‘presence’ is useful in revealing ideas about what is conscious or animate (Edwards, 2022). In an Aboriginal cultural landscape, ancestral beings respond to human engagement by increasing or withholding resources, offering healing or injury, or by expressing grief – for example causing waterholes to dry up when a clan ‘boss’ for that site dies. As is typical in such ‘nature religions’,2 supernatural beings live locally. But any notion of sacredness requires presence, and, even in the more distant abstractions of monotheistic religions, God is omniscient and all-seeing (Figure 5). The eye of God, the hand of God and the Sacred Heart. Source: Stipple engraving by C. Savini; photo Wellcome Collection.
Stewart Guthrie (1995) proposed that all such imaginaries arise from a cognitive predisposition to project human personhood into the world. Hegel (1979) described a process of projection and integration, in which human consciousness expands epigenetically through dialectical engagement with the world, and Varela (whose work acknowledged Buddhist teaching that conceptualises the mind as a mental activity of experiencing life) presented a view of an embodied mind integrating cognitive processes with phenomenological experience (Varela et al., 1991). Bloch (2012) has encouraged re-engagement with the cognitive sciences, and other more recent work in this area has articulated an extended mind thesis (Clark, 2008, 2017; Menary, 2010). This suggests that, rather than being contained within the brain, or even the body and its senses, the human mind extends into the environment in a variety of ways. These writers point to records – for example notes – as extensions of the mind, but there are many ways in which memory and meaning are located externally. In an indigenous Australian lifeworld, for example, the land and waterscape is a repository for the Law, with sacred sites acting as effective mnemonics for stories and events in which totemic ancestors actively respond to human action.
There are clearly many interesting questions about how beliefs about the sentient presence of ancestral or other supernatural beings intersect with cognitive processes that recursively extend the mind into the world. But with regard to kinship, we need to focus on how this extension affects societal engagement with the non-human domain. Diverse cosmological schemes may be united by a common human need to project the mind outwards, but there are major divergences between indigenous schemes envisioning a single, indivisible world inhabited by localised totemic ancestors with supernatural powers, and the dualism of monotheistic religions, in which omniscient powers sit with a more abstract, distant and fully humanised father figure, and the non-human domain is alienated from and subject to human dominion.
In a recent major study examining different trajectories in human–environmental relations, I have suggested that the latter bifurcation arose from some key developmental factors (Strang, 2023). Archaeological records suggest that early human societies worshipped ancestral deities manifesting aspects of the non-human world, who took multiple female, male (or androgynous) and often non-human forms. There is no space here to unpack the diverse and complex historical and cultural contexts in which specific changes occurred, and their many and fluid variations, but this large-scale comparative analysis illuminated an interesting recurrent pattern. It would appear that, repeatedly, as social and economic arrangements became more hierarchical and patriarchal, there was a broad tendency for deities to be humanised, becoming first semi-human, then human in form (with some slippage back and forth). Multiple non-human beings were thus superseded by deified rulers (monarchs and emperors), pantheons of humanised deities, and in the major monotheisms by a single ‘God the Father’. And it would appear that these religious and political changes were driven, in part, by technological changes enabling new and more intensive modes of production and greater material control over the non-human domain.
This suggests that Durkheim’s (1961) view of the relationship between religious and socio-political developments requires a ‘third pillar’, recognising the centrality of the physical power and authority conferred by instrumental technologies, and their capacity to transform inter-human and interspecies relations. This locates our discussion within societies’ long-term historical trajectories of development, from the changes initiated by early shifts into agriculture (Diamond, 1987) to the Industrial Revolution. In effect, processes of enclosure appropriated non-human habitats, reforming them and redirecting resources to support human activities. There was a notable intensification of these transformative processes, and further alienation between human and non-human beings, with the emergence of 19th-century capitalism (Berger, 2009). Federici (2018, 2019) similarly observes that pre-capitalist continuities between humans and animals were necessarily disrupted to create the disciplined workforces that capitalism required, and this discontinuity supported Cartesian dualism in its various forms, as well as the progressive enclosures of the commons that dispossessed many human groups, and especially women.
Feminist theorists have therefore pointed out that processes supporting patriarchal authority led, concurrently, to widening gender inequalities and to colonial visions of indigenous people as less than human, as well as introducing greater division in human–non-human relations (Mies, 1986; Spencer-Wood, 2016; see also Meillassoux, 1981). Although there is insufficient space here to discuss the gender and colonial issues that run parallel with this article, I do want to note that just as inter-human inequalities have emerged with the changes I have described, feminism has long been central to social and ecological countermovements seeking to re-establish more equitable interspecies relations (Gaard, 2017).
The long view suggests a broad process of change in which growing instrumentality has tended to support beliefs and practices that widen inter-human and human–non-human inequalities. It is only in place-based societies able to retain long-term sustainable lifeways that more egalitarian relations have persisted, and it is these groups that provide not only vital alternate perspectives but also dogged resistance to globally dominant – and globally unsustainable – ideologies and practices.
In the major monotheisms, the division between humankind and others was explicated in religious texts that disassociated humans from ‘the beasts of the field and forest’ and asserted that only humans have ‘the breath of God’, that is, souls. Hence they were located ‘nearer to the angels’ and given dominion over the world and its non-human inhabitants. Even as evolutionary science traced the elaborate lineages and material connections between all species, strenuous religious efforts maintained this differentiation on the basis that spiritual being and reflexive consciousness placed humans in a separate and superior category.
Some anthropologists have been critical of this speciesist divide. Marilyn Strathern (2017, 2020), in considering the potential expansions and contractions of notions of relations, observes that: ‘When anthropologists talk about relations, it is persons who most often come first to mind, that is, beings inevitably enmeshed in a relational world, although these days persons may be other than human’ (2020: 11). Donna Haraway (2008, 2016) has elucidated interspecies (and material) connectivity, and promotes ‘making kin’ with other living kinds, with a view to encouraging greater ‘response-ability’. Anna Tsing (2014, 2017) has explored ‘more than human nature’ by treating non-human others – for example fungal spores – as ethnographic subjects. Indeed, a florescence of work on multispecies ethnography has made imaginative leaps into non-human lifeworlds to consider other-than-human perspectives (Kirksey and Helmriech, 2010; Lainé and Morand, 2020; Paerregaard, 2020; Simon, 2020).
A relational model has also been encouraged by Actor Network Theory (Latour, 2005), Systems Theory, and New Materialism (Bennett, 2009; Coole and Frost, 2010). Foregrounding the dynamic relations between all living kinds and the environments and processes in which they coexist highlights the co-creative role of the non-human domain in composing shared lifeworlds. This also raises questions about the dualism inherent in categories of ‘cultural’ and ‘biological’ heritage, and it may be that a single category of biocultural heritage would prove more useful in acknowledging the co-creative contribution of the non-human domain to all forms of legacy (Strang, 2022). Anthropology, therefore, has a range of theoretical approaches that allow us to consider diverse ideas about social and material relations between species, and it is here that I think that concepts of kinship may have useful practical applications in supporting more sustainable modes of human–environmental relations.
There are infinite ways in which human–non-human relationships may be constituted, but they always contain relations of power, even when – as in many place-based societies – these are framed in terms of partnership and reciprocal exchange (Fienup-Riordan, 2017). Although archaeology demonstrates that plants and animals have long been domesticated, in recent centuries human material control over the non-human world has radically expanded, invariably at the expense of ‘wild’ habitat and undomesticated species (Figure 6). Deforestation in Madagascar. Source: Photo Wikimedia Commons.
While concepts of kinship and partnership with non-human beings might persist in small-scale societies, in most larger societies, although kin relations may be composed with family pets, the industrialised farming that is now central to food production depends upon relationships of enslavement, dispatch and consumption that require a much greater degree of alienation. This ‘othering’ or ‘unkindness’ is supported by ideas about human exceptionalism and a dualistic vision of culture and nature. Casting non-human beings as being fundamentally unlike and inferior to ‘us’ enables levels of anthropocentricity in which it becomes permissible not only to consume them, but also to destroy their habitats, to pollute their ecosystems, and to override their needs and interests by externalising all of the costs of human activities to them. While domesticated species may be protected, and household pets accepted as kin, this is often at the cost of other species and their habitats. This is demonstrated, for example, by the detrimental environmental impacts of intensive farming, and by the ecological costs of feeding meat to the vast numbers of dogs and cats housed as domestic pets (the latter also being avid predators of local wildlife). And even in cultural contexts where some species are especially valorised or regarded as sacred, for example in Hindu traditions, this does not mean that the groups concerned eschew unsustainable economic practices with detrimental impacts on other species and environments.
Expanding kindness
In considering how to challenge an unsustainable status quo, there is much to be learned from theories critiquing anthropocentricity in human–non-human relations, and from indigenous perspectives that provide exemplars of undifferentiated worlds in which humans engage collaboratively with non-human beings and environments. As noted at the outset, there is a need to promote a paradigmatic shift towards a recognition of the literal ‘kind-ness’ shared by human and non-human beings, and the interspecies and material relations that co-create shared and indivisible lifeworlds. But such abstractions, while offering new positionalities, do not directly address the inequalities in power that characterise exploitative human–non-human relationships.
These are more directly addressed by recent debates about non-human rights. Having written about these in detail elsewhere (Strang, 2020a, 2021), I will simply note a few key points here. Internationally, there is growing pressure on the United Nations to adopt a declaration of non-human rights to complement the Declaration of Human Rights made in 1948. There have been similar national efforts: for example, in Ecuador the rights of Pachamama (Mother Earth) have been enshrined in the constitution. Progress is being made towards ‘ecocide’ being registered as an international crime under the aegis of the International Court of Criminal Justice, and there have been myriad attempts to establish ‘Earth Law’: national and international regulations protecting the rights of non-human species and ecosystems. There is some coalescence between indigenous people’s critiques of the capitalist modes of production that entail exploiting and destroying their homelands and activist groups seeking social and ecological justice; alternate religious movements sympathetic to non-human interests and aiming to reconnect with ‘nature’; and efforts to promote de-growth or circular economic systems.
Although the focus of this article is on interspecies relations, it is worth noting that many of the arguments in it could be as readily applied to inequalities in inter-human relations, in particular situations in which certain human groups (based on differences in culture, gender, sexuality, caste, etc.) are also ‘othered’, and regarded as lesser human beings, or even as less than human, and therefore excluded from those who might be regarded as kin. Such othering can be similarly challenged by conceptual inclusion, as demonstrated for example by the ‘adoption’ of refugees in Australia, described by Maila Stivens (2023).
Of particular relevance here are campaigns in Colombia, India and New Zealand to define rivers as persons, with concomitant legal rights. As mentioned above, in 2017 the New Zealand government accepted a Māori cultural view of rivers as living ancestors, or Te Awa Tupua, and assigned legal personhood to the Whanganui River as a ‘living entity’ with rights equivalent to those of corporate ‘legal persons’ (Muru-Lanning, 2016; New Zealand Government, 2017a). In other cases a forest attained similar rights and status (New Zealand Government, 2014; see also Stone, 1972), as did Mount Taranaki/Taranaki Maunga (New Zealand Government, 2017b). These efforts to assert the kinship of the non-human domain cohere with indigenous activism similarly seeking to underline pan-human kinship and the social and legal obligations of the state (as pursued in this volume).
These decisions rest, fundamentally, on the notion of non-human beings and ecosystems as ancestral kin. Forests and rivers, and the taniwha (serpentine water beings) that manifest their generative powers, are present, sentient ‘living ancestors’. So in this sense ideas about kinship are already being used to challenge human–non-human inequalities and the unsustainable practices that have led to the current environmental crisis (Strang, in press). How might ideas about kinship promote more sustainable practices in other cultural contexts?
Notions of kinship, in whatever form, are fundamentally categorisations of beings ‘akin’ to ourselves, to whom we must offer care and protection, and those ‘unlike’ ourselves towards whom we feel no such obligation. For humans to be humane, there is a need to bring into public discourses inclusive ways of imagining non-human worlds that encourage greater co-identification or ‘kindness’. As I have suggested elsewhere, if societies are to achieve more sustainable lifeways they need to ‘re-imagine communities’ to encompass not only less powerful human groups, but also the non-human co-inhabitants of ecosystems (Strang, 2017, 2020b). This provides a basis for establishing non-human rights and forms of pan-species democracy that would uphold these. Such inclusiveness would enable new forms of conviviality – living together – in which, with the aid of people to ‘speak for’ them in decision-making fora, the needs and interests of non-human beings and living systems could be articulated and given ethical weight. To be effective, such reconceptualisations and their related representational practices would need to be multi-scalar: enacted within river catchments, carried into regional and national policy-making, and incorporated into the international debates that will define human and non-human futures.
The concept of kinship provides anthropology with a very effective way of articulating diverse and fluid ways of thinking about identity, social inclusion and access to the commons. Conventionally, it has been employed to elucidate social and material relations between humans, and more specifically to critique the enclosures and appropriations leading to widening inequalities between them. However, broadened to be more inclusive of non-human beings, kinship can contribute imaginatively to these debates. It has a central role to play in encouraging the recognition of our shared evolutionary and cultural heritage with non-human species. It raises key questions about how we conceptualise and therefore enact our relations with the non-human domain, and it provides a strong foundation from which to promote the need to extend ‘kind-ness’ to all living beings.
Footnotes
Note
Author Biography
![]()