
Abstract
SARS-CoV-2, the causative agent of COVID-19, caused respiratory disease outbreaks with increased mortality in 4 mink farms in the Netherlands. The most striking postmortem finding was an acute interstitial pneumonia, which was found in nearly all examined mink that died at the peak of the outbreaks. Acute alveolar damage was a consistent histopathological finding in mink that died with pneumonia. SARS-CoV-2 infections were confirmed by detection of viral RNA in throat swabs and by immunohistochemical detection of viral antigen in nasal conchae, trachea, and lung. Clinically, the outbreaks lasted for about 4 weeks but some animals were still polymerase chain reaction–positive for SARS-CoV-2 in throat swabs after clinical signs had disappeared. This is the first report of the clinical and pathological characteristics of SARS-CoV-2 outbreaks in mink farms.
Keywords
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) belongs to the Betacoronavirus genus of the Coronaviridae family and is currently causing a pandemic disease named coronavirus disease of 2019 (COVID-19). The virus rapidly spread among humans even in the face of a range of interventions. 6 Recently, we reported outbreaks of SARS-CoV-2 in farmed mink (Neovison vison) in the Netherlands with animal-to-animal spread, but so far all other natural cases in animals seem to be limited to incidental spillover from humans. 7 This includes cases in dogs, cats, and tigers that tested positive for SARS-CoV-2 RNA, and infection in the latter 2 species were associated with clinical respiratory disease. 4,10,12 Reports of infections with SARS-CoV-2 in other animal species are mostly limited to experimentally induced conditions with either mild to moderate disease in hamsters and juvenile cats, subclinical viral replication in ferrets (which are closely related to mink), and a few other species including cynomolgus macaques, cats, and Syrian hamsters. 2,4,5,8,9 This report describes the clinical presentation and the pathological findings of SARS-CoV-2 outbreaks on 4 mink farms (NB1, NB2, NB3, and NB4) in the province of Noord-Brabant in the Netherlands.
Mink on these 4 farms were housed in wire netting cages placed in halls (NB1, NB2, and NB4) or sheds (NB3). Each cage housed one adult mink, and after whelping (end of April to early May) it also housed their kits. The population before whelping consisted of 12 000 female and 1700 male mink on NB1, 7500 females on NB2, 1550 females on NB3, and 10 300 females and 242 males on NB4. The animal populations on all 4 farms had been positive for Aleutian disease virus for at least 10 years but clinical problems due to this virus were minimal since farmers were breeding for resistance against the disease.
Increased mortality and respiratory signs in a small number of adult mink were reported on 19 and 20 April 2020 in farms NB1 and NB2, respectively. Histological evaluation showed interstitial pneumonia and lung samples of these mink tested polymerase chain reaction (PCR)-positive for SARS-CoV-2 and PCR-negative for both influenza A virus and canine distemper virus, while routine aerobic cultures from these lungs yielded no growth on sheep blood agar plates. Thus, the diagnosis was interstitial pneumonia caused by SARS-CoV-2. On 6 May, the same diagnosis was reached on farms NB3 and NB4. In contrast to the other 3 outbreaks, clinical signs on NB4 were nearly absent at the time of detection and became apparent only at a later stage.
On all farms, clinical signs in adult mink included labored breathing and watery to mucoid nasal exudates, with severity ranging from barely noticeable to very severe. In individual animals, these clinical signs were recognized for 1 or 2 days after which the animal typically stopped eating and was found dead the next day (ie, 2–3 days of clinical illness was typical). Farmers noted that all animals with moderate to severe signs died. Females that became ill and stopped eating in the first weeks after whelping often lost a large number of their kits. No clinical signs were noted in these kits before they died, and it is not known how many died due to viral infection or indirectly due to a lack of maternal care.
At the peak of the outbreaks, the morbidity (ie, the percentage of adult mink that showed moderate to severe respiratory signs or refused to eat that day) was approximately 2 and 3 times the daily mortality (ie, the percentage of adult mink that died that day). Despite notable variations in severity, the duration of clinical disease on each farm was approximately 4 weeks during which time the mortality was increased (Fig. 1). On farm NB1, mortality was logged in detail and was found to be unrelated to sex (3.7% in males and 3.8% in females) or color type, and mortality cases were not clustered but instead widespread within the farm.
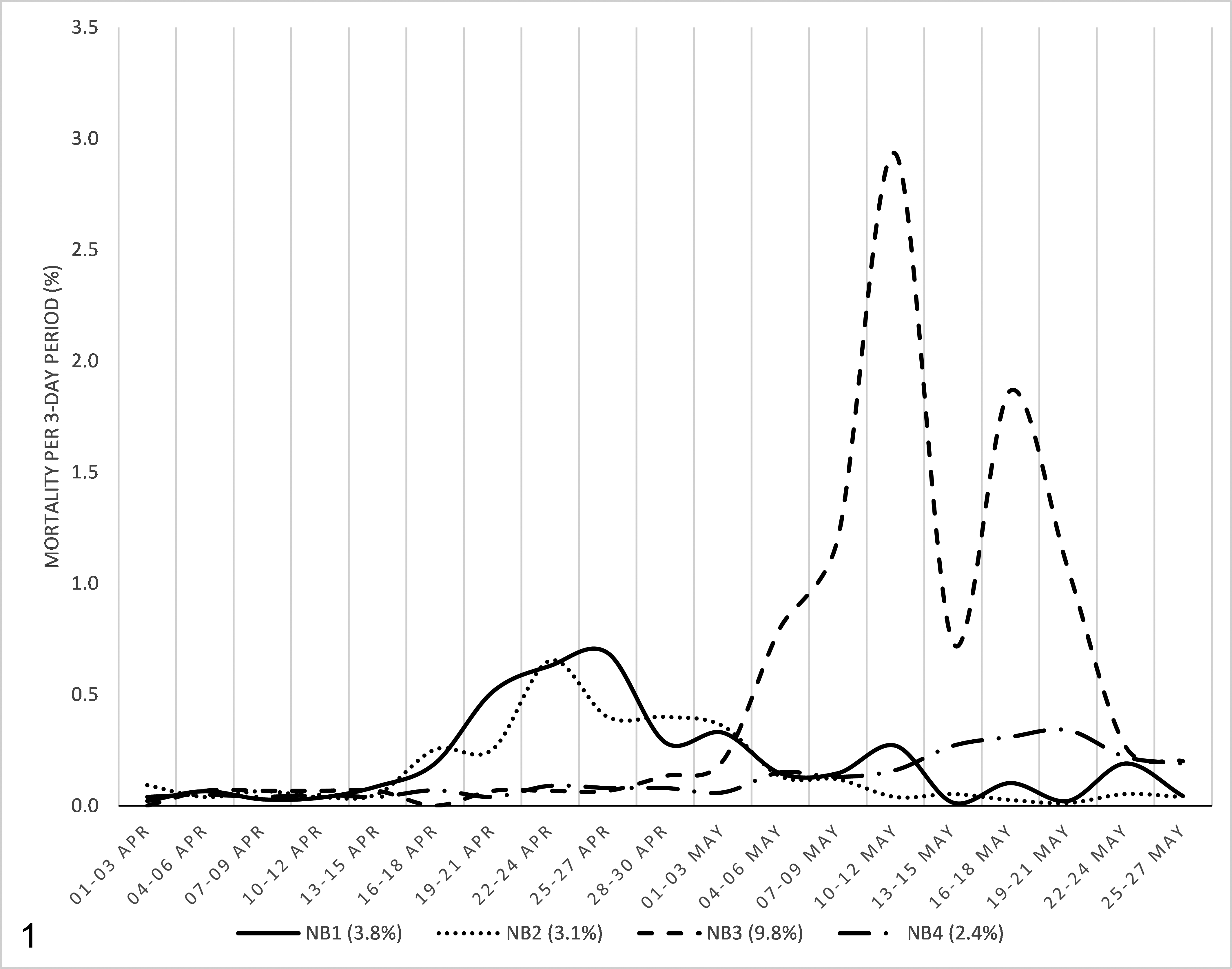
Incidence of mortality in adult mink at farms NB1, NB2, NB3, and NB4 from 1 April 2020 until 27 May 2020 in each 3-day period. Mortality was calculated as a percentage of the whole adult mink population at the start of April. The total mortality on each farm during this time period is shown in parentheses. All 4 farms were at baseline mortality in the first 2 weeks shown on the graph.
From all 4 farms, fresh adult mink carcasses as well as kit carcasses were submitted for necropsy to evaluate the cause of death. At the first observation of the outbreaks, most adult animals (in total 38 out of 45) had severe pneumonia (Table 1) and a good body condition (Fig. 2), and none of them had indications of other significant lesions. In the affected lungs, all lobes were swollen, dark red, and failed to collapse (Figs. 3, 4). These animals had no or very little contents in the intestinal tract. Other than a few animals with abundant free blood in the nasal conchae and blood-staining around the nostrils, the upper respiratory tract was macroscopically normal. Adult mink that died without pneumonia had a wide variety of other lesions including hepatic lipidosis, chronic nephritis, signs of sepsis, dystocia, and urolithiasis, and often had poor body condition. In mink kits, the macroscopic findings were more subtle. Apart from some diffuse red-brown discoloration and a wet appearance of the lungs in less than 10% of the kits (Fig. 5), no significant macroscopic changes were noticed.
Frequency of Gross Lung Lesions and Detection of SARS-CoV-2 by PCR in Mink From 4 Farms (NB1 to NB4)a.
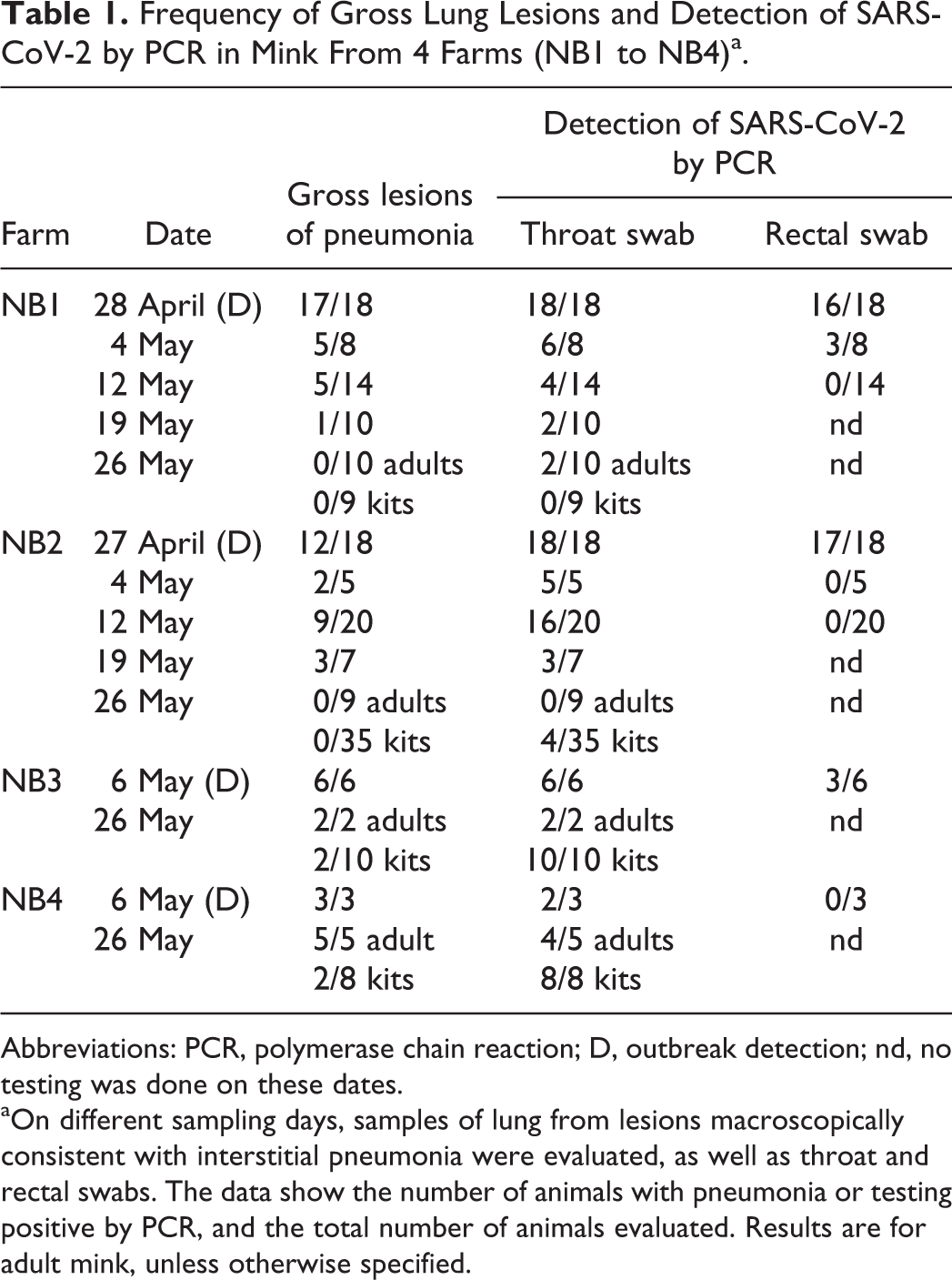
Abbreviations: PCR, polymerase chain reaction; D, outbreak detection; nd, no testing was done on these dates.
aOn different sampling days, samples of lung from lesions macroscopically consistent with interstitial pneumonia were evaluated, as well as throat and rectal swabs. The data show the number of animals with pneumonia or testing positive by PCR, and the total number of animals evaluated. Results are for adult mink, unless otherwise specified.
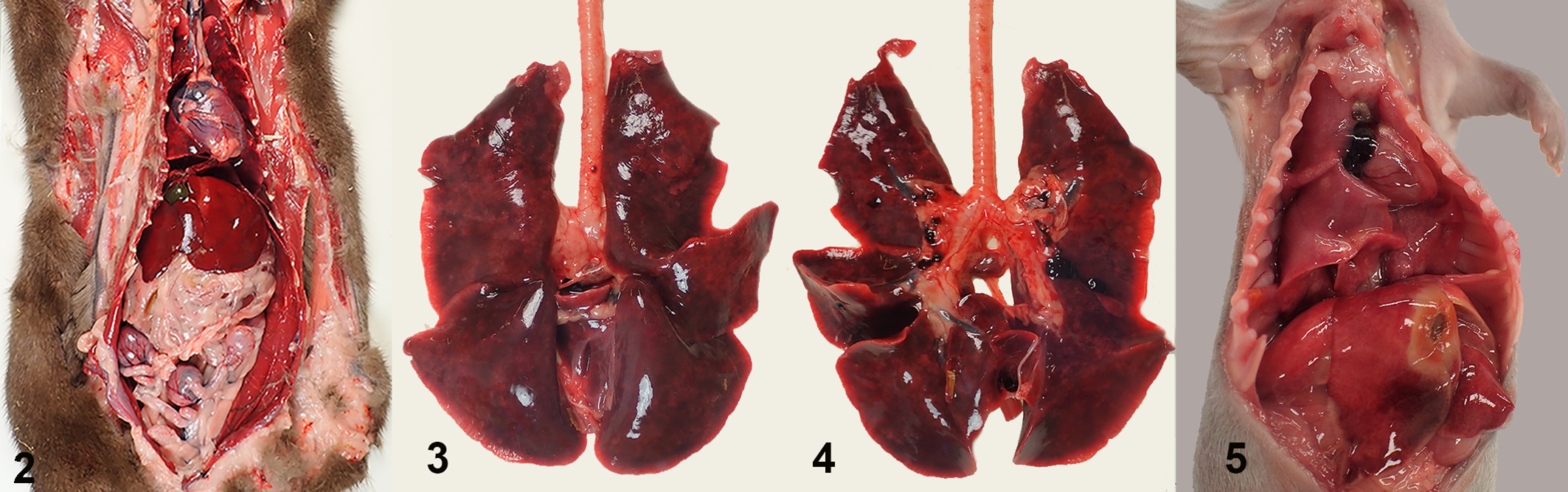
SARS-CoV-2 infection, lung, mink.
At the start of the outbreak and during the follow-up (until 4 weeks after initial detection of the disease), representative lungs from a total of 40 adult animals and 9 kits were sampled for histology. From 8 animals from NB1 and NB2, nasal conchae, trachea, liver, spleen, kidney, intestinal tract (duodenum, jejunum and colon), pancreas, and brain were sampled for histological evaluation on the day of detection. Histologic sections were prepared routinely and stained with hematoxylin and eosin. Consistent histologic findings in the macroscopically affected lungs of adult animals that were sampled for histology (35 of 40) were multifocal to coalescing areas with thickening and degeneration of alveolar septa, which were often lined by delicate hyaline membranes and showed moderate to severe type II pneumocyte proliferation (diffuse alveolar damage; Fig. 6). Thickening of the alveolar septa was due to fibrillar eosinophilic material and few mononuclear cells. Alveolar lumina were filled with mononuclear inflammatory cells, desquamated cells, and low numbers of neutrophils. These findings were most prominent in the area adjacent to bronchi, and the bronchi showed loss of cilia and some swelling and degeneration of epithelial cells. The epithelial cells of bronchioles in affected areas showed more severe changes with necrosis and formation of syncytial cells (Fig. 7). Other consistent findings included pulmonary alveolar edema with abundant foamy intra-alveolar macrophages, perivascular edema, and hyperemia of alveolar septa. There was variation in the severity of diffuse alveolar damage between individual animals, but there was no significant difference in severity between the individual farms.
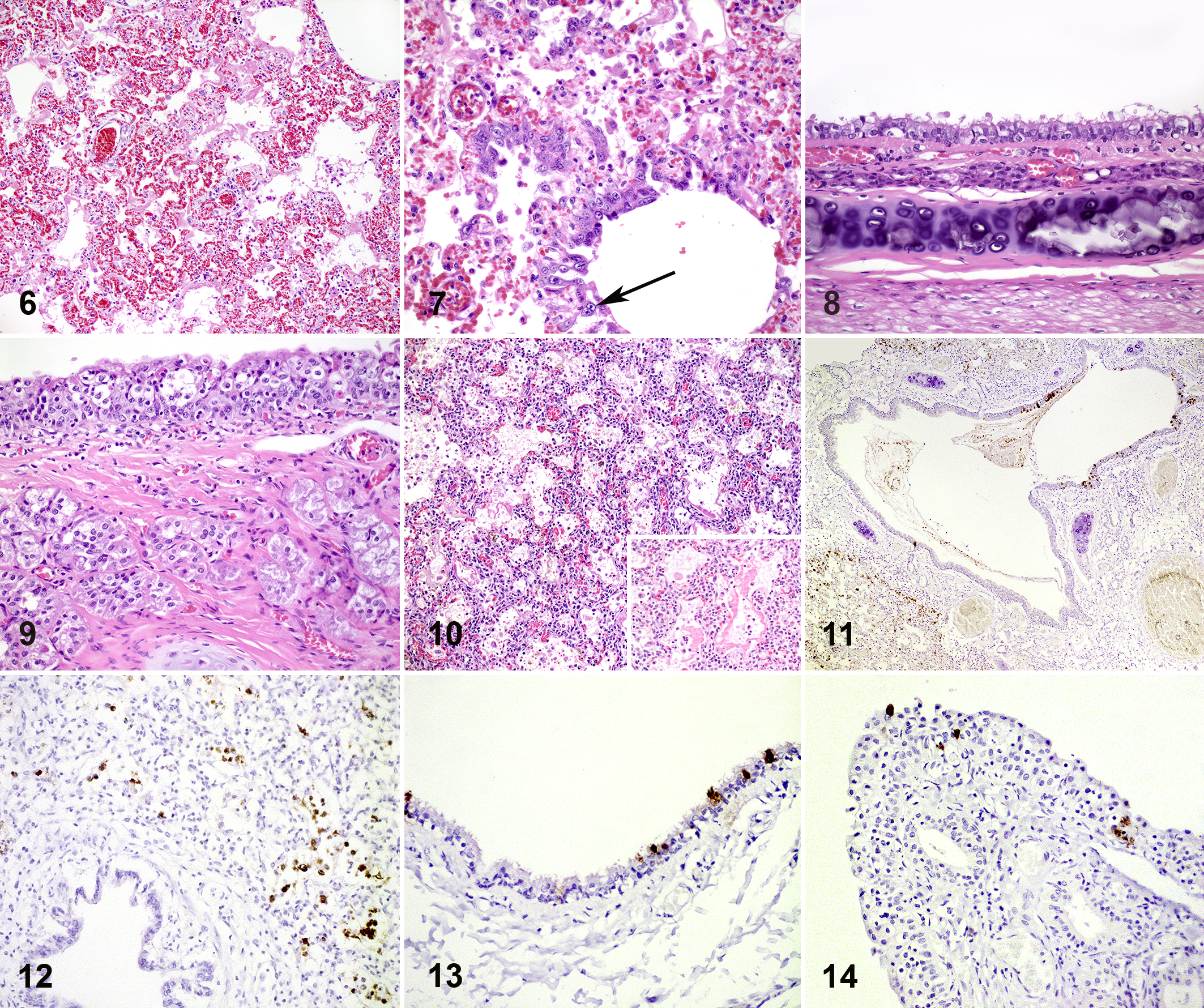
SARS-CoV-2 infection, mink.
In the upper respiratory tract, the trachea showed mild to moderate changes characterized by loss of cilia with some swollen and some flattened epithelial cells (Fig. 8), while the nasal conchae showed more severe changes characterized by multifocal swelling and degeneration of epithelial cells with diffuse loss of cilia (Fig. 9). Other investigated organs from adult animals had no significant histopathological lesions.
Lungs from 5 of 9 kits had lesions comparable to those in adult lungs, albeit without clear type II pneumocyte proliferation (Fig. 10). The kits without alveolar damage showed slightly thickened alveolar walls, with proteinaceous exudate and some foamy macrophages in alveoli.
This multifocal to diffuse alveolar damage in the lungs of adult animals and kits is compatible with a viral pneumonia such as that caused by influenza virus infections in mink and also described in SARS-CoV-2 infections in humans. 1,11 Other possible causes for diffuse alveolar damage (eg, inhalation of toxic gases, ingestion of toxins, or sepsis) were considered unlikely based on the clinical history, the contagious nature of the outbreak, the macroscopic findings, and negative bacteriological cultures. With immunohistochemistry (see Supplemental Materials for methods) we confirmed the presence of SARS-CoV-2 antigen in 4 of 11 adult lungs (Fig. 11) and 1 of 5 kit lungs tested (Fig. 12). Immunolabeling was present within epithelial cells of the bronchi and bronchioles, alveolar epithelial cells, and in cells in alveolar lumina that were most likely desquamated pneumocytes or macrophages as also observed in human lung biopsies. 13 In adult mink, viral antigen was also present in the trachea (Fig. 13) and nasal conchae (Fig. 14) within epithelial cells, which were often flattened. Not all lungs had immunolabeling for viral antigen, possibly because the virus is only present in lung tissue during the acute phase (first week) of the disease and because the viral load is lower in the lung compared to throat swabs. 2,7
Throat and rectal swabs were collected from all mink submitted for necropsy (Table 1) and were tested for SARS-CoV-2 by E gene PCR as described previously. 3,7 While positive PCR tests were more common in adult mink that had macroscopic lesions of pneumonia (60/70, 86%), adult mink without gross lesions of pneumonia also had positive PCR results (28/65, 43%). Concerning the sampling for PCR testing, the addition of rectal swabs did not increase the sensitivity of detecting infection; all mink that tested positive in rectal swabs were also positive in the throat swabs, but not all mink that tested positive in throat swabs tested positive in rectal swabs. In each case where both the rectal swab and the throat swab were positive, the Ct value of the throat swab was lower (indicating a higher viral load in the throat swab). Oreshkova et al described the viral load in different organs of these SARS-CoV-2 positive mink and found that more virus was present in the throat swabs and the nasal conchae than any other sample site. 7 Using PCR on throat swabs, viral RNA was still detected on all farms at the end of the observation period (26 May). Even on NB1 and NB2, where clinical abnormalities were no longer noted and no gross lung lesions were found at necropsy at that time, a low proportion of the samples was still PCR positive.
Additionally, rectal swabs were taken from 226 healthy mink distributed over farms NB1 and NB2 on 27–28 April, 12–13 May, and 25–26 May and tested for presence of SARS-CoV-2 RNA with PCR. On NB1, 11/51 (22%) were positive on the first sampling date, 0 of 47 were positive 2 weeks later, and 0 of 51 were positive 2 weeks after that. On NB2, 9/30 (30%) were positive on the first sampling date, 14 of 27 were positive 2 weeks later, and 0 of 20 were positive 2 weeks after that. The Ct values of positive swabs were mostly between 36 and 38 (ie, there were very low amounts of viral RNA on the swabs), and none of the Ct values were below 30.
The clinical and gross findings together with immunohistochemistry and PCR results indicate that SARS-CoV-2 infection can induce acute severe interstitial pneumonia or diffuse alveolar damage in mink, and this can cause respiratory signs with increased mortality on mink farms. Other organ systems were not affected in the examined mink. SARS-CoV-2 appears to also cause subclinical infection in some mink, because on the first 2 sampling dates, 34 of 155 healthy mink were PCR-positive in the rectal swabs. The large variation in morbidity and mortality at each farm suggests that outbreaks can be so mild that they could be easily missed. For diagnosis of SARS-CoV-2 infection on farms where mink have clinical signs, throat swabs in combination with histopathology of the lungs are recommended. For surveillance of farms without clinical signs, PCR testing of throat swabs from mink that recently died can be considered, even in the absence of gross lesions.
Supplemental Material
Supplemental Material, Supplemental_materials-Molenaar_et_al - Clinical and Pathological Findings in SARS-CoV-2 Disease Outbreaks in Farmed Mink (Neovison vison)
Supplemental Material, Supplemental_materials-Molenaar_et_al for Clinical and Pathological Findings in SARS-CoV-2 Disease Outbreaks in Farmed Mink (Neovison vison) by Robert Jan Molenaar, Sandra Vreman, Renate W Hakze-van der Honing, Rob Zwart, Jan de Rond, Eefke Weesendorp, Lidwien A. M. Smit, Marion Koopmans, Ruth Bouwstra, Arjan Stegeman and Wim H. M. van der Poel in Veterinary Pathology
Footnotes
Acknowledgement
The authors gratefully acknowledge the help and cooperation of the owners of each farm.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was commissioned and funded by the Dutch Ministry of Agriculture, Nature and Food.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.