
Abstract
A 4-year-old captive male central bearded dragon (Pogona vitticeps) was presented for recurrent episodic dyspnea and anorexia with occasional expulsion of oral mucoid discharge. Despite empirical antimicrobial therapy and supportive care, the animal died and was submitted for autopsy. Defining histologic features included heterophilic and lymphocytic interstitial pneumonia, with occasional amphophilic intranuclear inclusions and prominent type II pneumocyte hyperplasia. Transmission electron microscopy revealed intranuclear 80-nm, nonenveloped, hexagonal viral particles within pneumocytes. Helodermatid adenovirus 2 (HeAdV2) was determined as the etiologic agent through pan-adenoviral consensus polymerase (PCR) chain reaction and sequencing. Nucleic acid from a novel Mycoplasma sp. (provisionally called Mycoplasma pogonae) was identified by pan-generic PCR targeting the mycoplasma 16S ribosomal RNA gene with sequencing and phylogenetic analysis. As bacteria morphologically consistent with Mycoplasma sp. were not observed by special stains and transmission electron microscopy, the detection of M. pogonae nucleic acid is of indeterminate significance; however, M. pogonae and HeAdV2 coinfection may have exacerbated disease.
Keywords
A 4-year-old male captive central bearded dragon (Pogona vitticeps) was presented for episodic expiratory stridor and increased respiratory effort on exhalation of 1-week duration. The owner rarely took the animal outdoors and reported no contact with other reptiles during the 3 years of current ownership. Housing included a glass aquarium tank on sand substrate with ambient and basking temperatures and UVB exposure deemed appropriate by the clinical staff. The lizard’s diet included commercially available pellets formulated for adult bearded dragons, approximately 7 to 8 crickets offered every 2 to 3 days, daily mustard greens and canned pumpkin, and free choice tap water. On physical examination, the animal was bright, alert, and responsive and exhibited normal activity. Based on skin turgor and bilaterally sunken globes, the animal was estimated to be 5% to 10% dehydrated. The animal was in thin body condition, weighing 344 g. The animal displayed self-limiting episodes of mouth gaping and kyphosis lasting less than 1 minute, during which the animal’s beard turned black. The owner reported that in the days prior to presentation, these episodes had occasionally been accompanied by an expulsion of mucoid oral discharge.
Complete blood cell count interpretation revealed a lymphopenia (2.5 K/μL; reference range, 4.0–12.0) and mild monocytosis (1 K/μL; reference range, 0–0.5), along with a left-shift and toxic changes of the heterophil series (basophilic and foamy cytoplasm) consistent with a chronic-active inflammatory process. 18 Review of serum chemistry analysis identified a slight hypernatremia (175 mmol/L; reference range, 122–168) and hyperchloremia (137 mmol/L; reference range, 64–128) consistent with the dehydration observed clinically.
Evaluation of dorsoventral and left lateral whole-body radiographs revealed poor coelomic serosal detail attributed to poor body condition, distended gastrointestinal tract attributed to aerophagia, and hyperinflation of the lower respiratory tract (Suppl. Fig. S1). Instituted medical therapies included fluid therapy and administration of ponazuril (10 mg/kg per os [PO] q48 hours for 2 treatments), metronidazole (20 mg/kg PO q48 hours), and ceftazidime (23 mg/kg intramuscularly [IM] q72 hours) for presumptive coccidiosis and bacterial pneumonia. Six days into the course of treatment, the animal died and was submitted for postmortem examination.
At autopsy, the cranial one-third of pulmonary parenchyma was bilaterally hyperemic, with dilation of central lumina by abundant white, frothy, mucoid exudate (Fig. 1). Representative specimens of pulmonary exudate from each lung lobe were collected via a sterile cotton-tip applicator. Specimens were submitted for routine aerobic culture with inoculation of sheep blood and MacConkey agar plates for 48 hours at 37°C. Major organs were fixed in 10% neutral buffered formalin, routinely processed, cut into 5-μm-thick sections, and stained with hematoxylin and eosin. In addition, Gram stains were performed on a section of affected lung and trachea. Formalin-fixed lung was trimmed, dehydrated, postfixed in 1% phosphotungstic acid (w/v), embedded in LR White, sectioned, and stained with uranyl acetate and lead citrate for transmission electron microscopy (TEM). Pan-adenoviral consensus polymerase chain reaction (PCR) and sequencing and pan-generic Mycoplasma 16S ribosomal RNA (rRNA) PCR were performed on fresh-frozen lung. 16,17,20 To obtain additional sequence, 2 specific primers were designed from the sequence obtained, BDMycR (5′-GTGAGTTCCCCACCGAGTAA-3′) and BDMycF (5′-TTACTCGGTGGGGAACTCAC-3′), and used in conjunction with the previously reported universal bacterial 16S rRNA gene primers 27F (5′-AGAGTTTGATC[A/C]TGGCTCAG-3′) and 1492R (5′-G[C/T]TACCTTGTTACGACTT-3′). Detailed phylogenetic analysis methods for the Mycoplasma results can be found in supplemental materials. PCR for herpesvirus, ranavirus, paramyxovirus, and reovirus was also performed on the same lung specimen, all of which were negative. 11,18,19,21
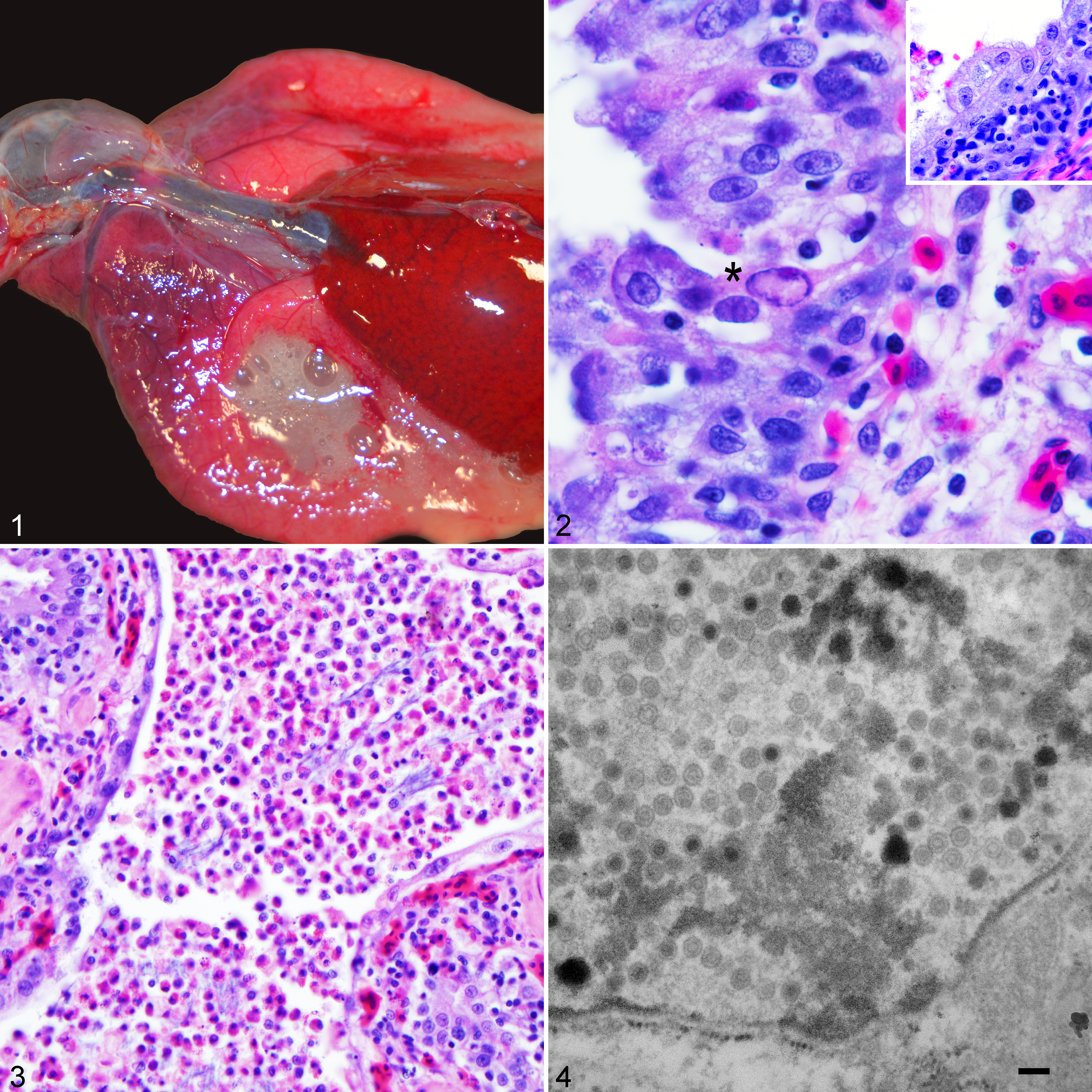
Viral pneumonia attributed to concurrent helodermatid adenovirus 2 and Mycoplasma pogonae infection, central bearded dragon.
The most significant histologic changes were present within the sharply demarcated hyperemic cranial aspects of lung. Piling up of pneumocytes was observed within the pulmonary parenchyma both centrally and peripherally, measuring up to 6 cells thick, indicative of type II pneumocyte hyperplasia (Fig. 2). Pneumocyte nuclei occasionally contained large, glassy, amphophilic to lightly eosinophilic, 10- to 12-μm inclusions, with peripheral displacement of chromatin (Fig. 2). Scattered pneumocyte degeneration and necrosis were present among areas of pneumocyte hyperplasia, represented by cytoplasmic vacuolation and karyorrhectic debris, respectively. Faveolar spaces were multifocally flooded by a mixture of mucus, edema, viable and degenerate heterophils, and necrotic cellular debris (Fig. 3). Squamous metaplasia of respiratory epithelium was observed segmentally adjacent to areas of faveolar flooding by heterophils (Fig. 3). Pulmonary interstitium was expanded by edema and a moderate number of heterophils, lymphocytes, plasma cells, and rare histiocytes. Occasionally, the aforementioned inflammatory cells were observed transmigrating the respiratory epithelium. Blood vessel hyperemia was prominent and diffuse. No bacteria were isolated from routine aerobic bacterial culture performed on pulmonary exudate collected from each lung lobe. However, low numbers of Gram-positive rods and cocci, as well as Gram-negative rods, were observed within mucoheterophilic exudate in both the tracheal lumen and faveolar spaces. Negative bacteriology results were attributed to antibiotic therapy preceding death. No intracytoplasmic bacteria were observed in sections of trachea or lung by Gram stain. The trachea displayed mild, multifocal hyperplasia of tracheal epithelium, with inflammation of the mucosa and submucosa by the aforementioned inflammatory cells. No intranuclear inclusions were observed in tracheal epithelium. No other significant findings were observed in other routinely evaluated organs. Evaluation of TEM revealed intranuclear viral particles within pneumocytes measuring approximately 80 nm in diameter that were nonenveloped and had hexagonal outlines with either electron-dense or electron-lucent cores (Fig. 4). PCR amplified a 272–base pair (bp) segment of adenoviral DNA (GenBank accession KY552649) that had 100% nucleotide homology to helodermatid adenovirus 2 (HeAdV2) (GenBank accession #KJ156523). In addition, PCR using pan-generic Mycoplasma 16S rRNA primers identified a 1425-bp amplicon (GenBank accession MG563382) that had 95% nucleotide homology to Mycoplasma iguanae (GenBank accession EU859973), herein provisionally referred to as Mycoplasma pogonae. The Bayesian tree and maximum likelihood bootstrap values are shown in Supplemental Figure S2.
Adenoviruses are nonenveloped, double-stranded DNA viruses that range from 60 to 100 nm in diameter. In most instances, adenoviruses appear to have coevolved with their host species and have high host specificity. 20 Documentation of host jumping in adenoviruses is uncommon and may have significant consequences. Titi monkey adenovirus 1 caused severe respiratory disease in titi monkeys (Callicebus cupreus) in a research facility, with interprimate monkey-to-human transmission observed. 3 In squamates, eublepharid adenovirus 1, apparently endemic in leopard geckos (Eublepharis macularius), has been seen in association with more significant disease in African fat-tailed geckos (Hemitheconyx caudicinctus). 20 Clinical disease associated with adenoviral infection is highly variable among animal species and is strain specific, with hepatitis, gastroenteritis, pneumonia, nephritis, and encephalitis having been described. 4,6 –9,13,15 Type II pneumocyte hyperplasia has been found with several different experimental models of adenovirus infection. 1,10,12 The 2 adenoviruses identified in bearded dragons, HeAdV2 and Agamid adenovirus 1 (AgAdV1), both belong to the genus Atadenovirus; the latter is the better known and first detected virus. 7
The benefit of consensus nested PCR targeting the conserved region of the DNA-dependent DNA polymerase gene of adenoviruses is the capability to detect novel atadenoviruses in squamates. 20 This is exemplified by the discovery of HeAdV2 from pooled tissue homogenates of intestine, liver, and heart from captive Mexican beaded lizards (Heloderma horridus). 14 The initial rationale for testing these animals arose following a mortality event in Gila monsters (Heloderma suspectum) and Mexican beaded lizards housed together in a Danish zoo. 14 Following the mortality event, oral swabs collected from both naive animals and survivors of the mortality event detected HeAdV2 in both species. 14 As the associated lesions have not been described, it has not been determined if HeAdV2 is a subclinical infection in a reservoir host or if the virus caused these events. Subsequently, HeAdV2 has been detected from the livers of a juvenile western bearded dragon (Pogona minor) with “ill thrift” and an adult central bearded dragon that “was not doing well.” 5 Of these 2 cases, histologic evaluation was only described from the liver of the western bearded dragon, with no significant findings observed. More recently, HeAdV2 has also been detected from cloacal swabs of apparently healthy wild Gila monsters in North America and a death adder (Acanthophis antarcticus) (J. F. X. Wellehan, personal communication, 2017). Taken together, these findings illustrate the capacity of HeAdV2 to infect multiple reptilian genera, including Heloderma, Pogona, and Acanthophis; however, the clinical significance of identifying HeAdV2 in these species remains enigmatic and requires further investigation, including experimental infection studies.
Phylogenetic analyses show that the Mycoplasma sp. identified in this study clusters in the Mycoplasma hyopneumoniae clade (Bayesian posterior probability 100%, maximum likelihood [ML] bootstrap value 100%) and is most closely related to M. iguanae (GenBank accession EU859973; Bayesian posterior probability 100%, ML bootstrap value 100%). M. iguanae was isolated from green iguanas (Iguana iguana), and experimental infection did not induce lesions. 2 The role of this Mycoplasma sp. in the disease seen in this bearded dragon is unknown. Proliferative tracheitis and pneumonia have been attributed to Mycoplasma sp. in other squamates. 15 It is possible that coinfection with both agents may have exacerbated disease; however, coinfection studies are necessary to further investigate this possibility.
Herein we present the first report of disease associated with HeAdV2 infection, affecting a central bearded dragon, with molecular evidence to suggest concurrent mycoplasma infection. We propose the name Mycoplasma pogonae for the novel mycoplasma based on the PCR and sequencing data. Defining histologic features included interstitial pneumonia with intranuclear inclusions and prominent type II pneumocyte hyperplasia. Histologic findings also support concurrent secondary bacterial bronchopneumonia. TEM conclusively revealed virus particles morphologically consistent with those of the Adenoviridae family. Virus particles were located primarily within the nuclei and were rarely present within membrane-bound cytoplasmic vesicles. Due to a paucity of literature describing viral pneumonia in bearded dragons, other viruses causing pneumonia in reptiles (including herpesviruses, ranaviruses, paramyxoviruses, and orthoreoviruses) were tested for by PCR and not found. HeAdV2 was recognized through consensus nested PCR targeting of the conserved region of the DNA-dependent DNA polymerase gene followed by sequencing. The source of HeAdV2 in this case remains unknown. Reactivation of persistent adenoviruses has been associated with disease in other species. 9 With this in mind, initial exposure may have occurred via contact to another reptile preceding current ownership. Overt immunosuppression was not evident; however, the animal was lymphopenic. It is possible that there are clinically significant differences among HeAdV2 strains; while strain diversity has not yet been investigated in HeAdV2, it will likely require examination of genes beyond the polymerase. The apparent respiratory tropism seen in this case contrasts significantly with the primarily enterohepatic disease previously reported with AgAdV1 in bearded dragons. Knowledge gained from this report will help both clinicians and diagnosticians alike when considering differential diagnoses for both nonspecific (eg, lethargy and inappetence) and more organ-specific (eg, dyspnea and open mouth breathing) clinical signs observed in bearded dragons.
Supplemental Material
Supplemental Material, DS1_VET_10.1177_0300985818780451 - Pneumonia in a Captive Central Bearded Dragon With Concurrent Detection of Helodermatid Adenovirus 2 and a Novel Mycoplasma Species
Supplemental Material, DS1_VET_10.1177_0300985818780451 for Pneumonia in a Captive Central Bearded Dragon With Concurrent Detection of Helodermatid Adenovirus 2 and a Novel Mycoplasma Species by Nicholas A. Crossland, Peter M. DiGeronimo, Yulia Sokolova, April L. Childress, James F. X. Wellehan, Javier Nevarez, and Daniel Paulsen in Veterinary Pathology
Footnotes
Acknowledgements
We thank Alex Ross and Imani Brown for their technical assistance in figure preparation and the Core Microscopy Center of Louisiana State University School of Veterinary Medicine, Baton Rouge, LA.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplementary material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.