
Abstract
C-terminal Src kinase (Csk) is one of the critical negative regulators of the Src family of kinases. The Src family of kinases are nonreceptor tyrosine kinases that regulate inflammation, cell proliferation, motility, and adhesion. To investigate potential histologic lesions associated with systemic loss of Csk gene activity in adult mice, conditional Csk-knockout mice were examined. Cre-mediated systemic excision of Csk induced by tamoxifen treatment resulted in multiorgan inflammation. Specifically, induction of Csk gene excision with three days of tamoxifen treatment resulted in greater than 90% gene excision. Strikingly, these mice developed enteritis that ranged from minimal and suppurative to severe, fibrinonecrosuppurative and hemorrhagic. Other inflammatory lesions included suppurative pneumonia, gastritis, and myocarditis, and increased numbers of inflammatory cells within the hepatic parenchyma. When tamoxifen treatment was reduced from three days to one day in an effort to lower the level of Csk gene excision and limit lesion development, the mice developed severe suppurative to pyogranulomatous pneumonia and minimal to mild suppurative enteritis. Lesions observed secondary to Csk gene excision suggest important roles for Csk in downregulating the proinflammatory activity of the Src family of kinases and limiting neutrophil-mediated inflammation.
Keywords
The Src family of kinases (SFKs) are a family of nonreceptor tyrosine kinases with critical roles in inflammation, 3,5,10 –12,14 –16,19,20 cell signal transduction, 5,10 –12,15,16,20 motility, 14,15,20 adhesion, 14,15,19,20,23 and proliferation. 1,14,15,20 The nine members of the SFKs are Blk, Fgr, Fyn, Hck, Lck, Lyn, Src, Yes, and Yrk. 1,10,14,15,20 SFKs have important roles in inflammation, including activation of neutrophils (including subsequent to integrin binding), monocytes, and macrophages; 3,5,10,11,15,19 adhesion of neutrophils to endothelial cells; 3,15 IgE-mediated stimulation of mast cells, 5,10,11 eosinophils, 5 and basophils; 5 and increasing vascular permeability. 15 Furthermore, SFKs play critical roles in transmitting the early signals in the signal transduction cascade resulting from T cell receptor 5,10 –12,15,16,20 and B cell receptor 5,10,20 engagement, which in turn regulates the activation state and development of both cell types. 4,16,17 Some SFKs have wide patterns of expression (eg, Src, Fyn, and Yes) 15,16,20 while the expression of others is limited to cells of certain lineages (eg, Lck is expressed in T and B cells; Hck is expressed in granulocytes, monocytes, and macrophages). 2,5,10,11,13,15,16,20
Regardless of the tissue distribution, the kinase activities of all SFKs are downregulated in a similar manner, namely phosphorylation of a tyrosine residue in a conserved COOH-terminal domain by C-terminal Src kinase (Csk) and to a lesser extent by CSK-homologous kinase (Chk). 1,2,5 –8,10,12 –16,19,20,23 Consistent with the number of important cellular processes regulated by SFKs as described above, systemic loss of Csk activity is embryonically lethal in mice, 1,6,8,13,14,17,19 with homozygous knockout embryos exhibiting growth retardation, 8,13,19 defective neural tube development, 1,8,13,14,19 and allantoic abnormalities. 8 These embryos concomitantly have increased Src, 6,8,13 Fyn, 6,8,13 and/or Lyn activity. 13 Csk-null embryos die between days E9.5 and E10.5. 8,13
Because Csk-null embryos are not viable, 1,6,8,13,14,17,19 a number of cell type-specific and tissue-specific models of Csk loss have been generated to investigate the effect of Csk loss in vivo. When Csk activity is lost from granulocytes via conditional mutagenesis, the resultant mice develop suppurative blepharitis, otitis externa, dermatitis, and pneumonia and have increased hematopoiesis in the bone marrow, spleen, and liver. 19 Neutrophils from these mice are hyper-reactive with increased adhesion and degranulation, and the mice are extremely susceptible to lipopolysaccharide. 1,19 Loss of Csk from thymocytes results in abnormal T-cell development and maturation with concomitant enhanced Fyn and Lck activity. 17,23 When loss of Csk activity is limited to keratinocytes, mice develop epidermal hyperplasia, 6,23 hyperkeratosis, 6 ulceration, 6 chronic mononuclear dermatitis, 6,23 and papillomas without malignant transformation. 6 Keratinocytes from these mice have elevated Fgr, Fyn, Src, and Yes activity 6 and increased phosphorylation of SFK targets. 23 When Csk is knocked out from intestinal epithelial cells, adult mice develop small intestinal and colonic epithelial hyperplasia. 7
In an effort to elucidate potential liabilities associated with the development of a systemic Csk inhibitor, work was undertaken to investigate the effect of systemic loss of Csk expression on the phenotype of adult mice. To avoid embryonic lethality, mice expressing Cre recombinase regulated by the estrogen receptor T2 (ERt2) domain 18,21 were crossed with mice homozygous for Csk flanked by LoxP sites. 17 The resultant Csk conditional knockout (cKO) mice were treated with tamoxifen for either three days or one day to induce Csk gene excision. Herein, we detail the lesions that adult Csk cKO mice develop following Csk gene excision.
Materials and Methods
Mouse Genetic Construct
The Csk conditional knockout and control mice were on a mixed C57BL/6 J 129P2/OlaHsd background and were obtained from the Jackson Laboratory (Bar Harbor, ME). Csk conditional knockout (Csk cKO) mice were B6.129P2 - Csktm1Tara/J, 17 Gt(ROSA)26Sortm9(Cre/ESR1)ArteCreERT2. 18 B6.129P2 - Csktm1Tara/J 17 mice were commercially available from the Jackson laboratory and were on a mixed C57BL/6 J 129P2/OlaHsd background (Jackson Laboratory stock #018320). Gt(ROSA)26Sortm9(Cre/ESR1)ArteCreERT2 18 mice were on a C57BL/6J background and were originally obtained from Taconic Biosciences (Rensselaer, NY) and then subsequently bred at the Jackson Laboratory. Genotyping was performed at the Jackson Laboratory. The Csk cKO mice described herein were homozygous for the floxed B6.129P2 - Csktm1Tara/J 17 transgene and hemizygous for the Gt(ROSA)26Sortm9(Cre/ESR1)ArteCreERT2 18 transgene. The control animals were homozygous for a wild type Csk allele and hemizygous for the Gt(ROSA)26Sortm9(Cre/ESR1)ArteCreERT2 18 transgene. Although the Csk cKO and control mice used for the studies were age, sex, and strain matched, they were not necessarily littermates. Tamoxifen treatment was required to induce Cre-mediated excision of the floxed allele in the Csk cKO animals. Csk cKO and control mice were assigned to their respective experimental groups so that both genotypes were sex and age matched and the tamoxifen treatment regimen was identical for each mouse included in an experiment. Table 1 provides a summary of the mouse cohorts (including mouse ages) and endpoints evaluated. Sufficient age matched Csk cKO and control female mice were only available for one experiment (Table 1, column 2). The other three experiments, therefore, used only male Csk cKO and control mice.
Summary of the Experimental Design for Comparing Csk Conditional Knockout and Control Mice.

aEqual numbers of Csk conditional knockout mice and age- and sex-matched control mice were used in each study.
Animal Husbandry
All mice were maintained at Invivotek (Hamilton, NJ) and were kept in a specific pathogen free barrier facility on a light:dark / 12 hours:12 hours cycle. The mice were fed a regular chow diet (Purina rodent diet #5053, Fisher Feeds, Bound Brook, NJ) throughout the studies. Food and water were available ad libitum. The majority of the mice in this study were group housed with 3 to 5 mice per cage. Shepherd’s ALPHA-dri bedding (Shepherd Specialty Papers, Watertown, TN) was used as the cage bedding. All procedures were approved by the Institutional Animal Care and Use Committee at Invivotek, an Association for the Assessment and Accreditation of Laboratory Animal Care International accredited facility.
Prior to arriving at Invivotek from the Jackson Laboratory, mice were confirmed to be serologically negative for the following viruses: mouse hepatitis virus/coronavirus, mouse parvovirus, pneumonia virus of mice, Sendai virus, Theiler’s mouse encephalomyelitis virus, mouse adenovirus, lymphocytic choriomeningitis, ectromelia virus (mouse pox), K virus, polyoma virus, epizootic diarrhea of infant mice/rotavirus, reovirus 3, mouse cytomegalovirus, Hantaan virus (hantavirus), mouse thymic virus, murine norovirus, mouse minute virus, and lactic dehydrogenase elevating virus. The mice were confirmed to be culture negative for Bordetella bronchiseptica, Cornebacterium bovis, Pasteurella spp, Salmonella spp, Streptobacillus moniliformis, Citrobacter rodentium, and Corynebacterium kutcheri. The mice were confirmed to be serologically negative for CAR Bacillus, Clostridium piliforme, Mycoplasma pulmonis, and Encephalitozoon cuniculi. The mice were PCR negative for Helicobacter spp. The mice were determined to be negative for endo/ecto parasites (pinworms via fecal flotation, fur mites via scotch tape testing, and examining cecal contents for visible worms) and opportunistic protozoa (eg, Giardia, Spironucleus via microscopic examination of intestine).
Induction of Csk Gene Excision
Csk cKO and control mice received tamoxifen (Sigma-Aldrich, St. Louis, MO) in 10% ethanol/ 90% sunflower seed oil (Sigma-Aldrich) via intraperitoneal (IP) injection once daily for either one or three days in an effort to induce gene excision. Each mouse received either 1.74 mg (for mice smaller than 23 g) or 2.0 mg (for mice larger than 23 g) of tamoxifen (Sigma-Aldrich) per day.
The extent of Csk gene excision was determined by Invivotek using Southern blot analysis. Per Taconic’s website, gene excision efficiency was anticipated to be greater than 70% in most tissues (https://www.taconic.com/mouse-model/creert2, last accessed 07.23.2017). DNA from spleens, lymph nodes, bone marrow, and/or skin were digested with EcoR1 (New England Biolabs, Ipswich, MA), fractionated on 0.8% agarose gels, transferred to positively charged nylon membranes (Biodyne B, Pall Life Sciences, Port Washington, NY) under alkaline conditions and hybridized using a 362 bp probe (the probe sequence can be found in the Supplemental Material). Following high stringency washes, blots were exposed to phosphoimager cassettes and scanned on the PharosFX Plus (Biorad, Hercules, CA). The extent of excision was determined using Quantity One imaging software (Biorad) by dividing the counts of the knockout band (KO: 2.1 kb Csk allele after tamoxifen induced Cre-mediated gene excision) by the sum of the counts of cKO band (2.5 kb floxed Csk allele before gene excision) and KO bands within each individual lane ([KO/(cKO+KO)] * 100). The wild-type Csk allele (present in the control mice) is 8.1 kb.
Necropsy and Histopathology
Mice were euthanized via isoflurane anesthesia (Midwest Vet Supply, Norristown, PA) followed by cervical dislocation. Necropsies were performed at Invivotek following euthanasia. The following tissues were collected from the mice that were either treated with 3 daily IP injections of tamoxifen and euthanized on days 10 through 12 or 1 IP injection of tamoxifen and euthanized on day 4, 7, or 14: colon, heart, kidney, liver, lung, pancreas, small intestine, sternum with bone marrow, and stomach. Spleens were also collected from mice treated with 3 daily IP injections of tamoxifen. Sections of skin from the pinna were available from male Csk cKO and control mice that received 3 daily IP injections of tamoxifen and were euthanized on days 10 through 12 and from male Csk cKO and control mice that received 1 IP injection of tamoxifen and were euthanized on day 4, 7, or 14. All tissues were fixed in 10% neutral buffered formalin. Formalin fixed tissues were shipped from Invivotek to Bristol-Myers Squibb in Lawrenceville, NJ for processing. Tissues were routinely, processed, paraffin embedded, and stained with hematoxylin and eosin (HE). HE-stained slides were evaluated by a board certified veterinary anatomic pathologist (LDBB) in a nonblinded fashion using a Zeiss Axio Scope.A1 (Oberkochen, Germany) with Jenoptik camera and ProgRes® CapturePro 2.8.9.3 software (Jena, Germany).
Serum Clinical Biochemistry
Blood was collected via cardiac puncture under isoflurane (Midwest Vet Supply) anesthesia during necropsy at Invivotek. For serum preparation, the blood was allowed to coagulate for 30 minutes at room temperature and then centrifuged at 1000 × g for 15 minutes at 4°C. The serum was analyzed on a clinical chemistry analyzer (Alfa Wassermann Alera, West Caldwell, NJ) using the manufacturer’s reagents according to the manual. Serum biochemical data were evaluated by a board certified veterinary clinical pathologist (DIB).
Hematology
Blood was collected via the retroorbital vein under isoflurane (Midwest Vet Supply) anesthesia at Invivotek. Hematological analysis of the peripheral blood was performed on the Hemavet HV950FS analyzer (Drew Scientific, Miami Lakes, FL) on the mouse setting. Hematology data was evaluated by a board certified veterinary clinical pathologist (DIB).
Results
Csk cKO Mice Treated With 3 IP Injections of Tamoxifen
Three-day tamoxifen induction: gene excision (data presented as average ± standard deviation)
Csk gene excision after three consecutive daily IP injections of tamoxifen in 10% ethanol/ 90% sunflower seed oil was evaluated in two cohorts of Csk conditional knockout (Csk cKO) mice (Table 1, columns 2 and 3). The first subset (Table 1, column 2) consisted of eight Csk cKO mice (4 male and 4 female) and eight control mice (4 male and 4 female) that were euthanized on days 10 and 11 (Csk cKO mice) or day 12 (control mice). There was nearly 100% Csk excision in the analyzed tissues from the Csk cKO mice, with average Csk gene excision of 99.1% ± 0.4%, 99.5% ± 0.61%, 98.9% ± 0.61%, and 91.2% ± 3.53% in the spleen, lymph nodes, bone marrow, and skin, respectively. Tissue differences in Csk gene excision efficiency may have been due to tissue differences in tamoxifen exposure 18 and/or chromatin structure affecting the accessibility of LoxP sites to Cre. 21
The second subset of mice (Table 1, column 3) were male Csk cKO and control mice (n = 5 mice/genotype) that were euthanized on day 7. The average Csk gene excision in the spleens of these Csk cKO mice was 93.1% ± 4.8%.
Three-day tamoxifen induction: body weight (data presented as average ± standard deviation)
Csk cKO (n = 8, 4 male and 4 female) and control mice (n = 8, 4 male and 4 female) were weighed daily and at necropsy 10 to 12 days after the initiation of the study (Table 1, column 2). Unexpectedly, male and female Csk cKO mice lost weight after the 3-day induction with tamoxifen (Supplemental Figure 1). Male Csk cKO mice lost an average of 13.2% ± 8.3% of their body weights, and female Csk cKO mice lost an average of 16.4% ± 2.8% of their body weights. In contrast, male control mice lost an average of 0.25% ± 1.6% of their body weight, and female control mice gained an average of 6% ± 2.9% of their body weight. Aside from the weight loss, no clinical signs or gross lesions were noted in the Csk cKO mice.
Three-day tamoxifen induction: histopathology
All 13- to 14-week-old Csk cKO and control mice that were injected with tamoxifen IP once a day for three days and euthanized on days 10–12 (Table 1, column 2, Csk cKO mice: days 10 and 11; control mice: day 12) exhibited varying degrees of peritonitis, mesenteric steatitis, and/or serositis that ranged from lymphohistiocytic to suppurative. This inflammation was presumed to be in reaction to the oil in the tamoxifen vehicle (10% ethanol/90% sunflower seed oil) since peritonitis secondary to usage of oil as a vehicle has been documented. 9,22
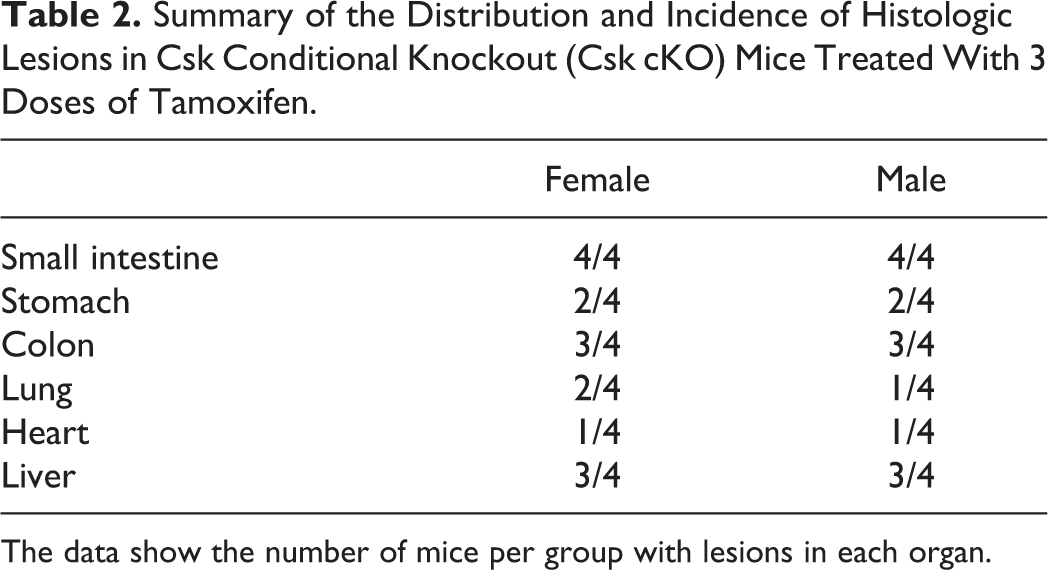
Female and male Csk cKO mice (n = 4/sex) exhibited a striking adverse histological phenotype characterized by suppurative inflammation in multiple organs. Affected organs included the small intestine, stomach, colon, lung, heart, and liver (Table 2). A summary of the histopathologic findings for each individual mouse can be found in Supplemental Table 1. Strikingly, all 8 Csk cKO mice exhibited varying degrees of suppurative enteritis. Five of the eight Csk cKO mice (62.5%) had enteritis that ranged from multifocal minimal to moderate and was composed of infiltrates of neutrophils with hemorrhage and varying numbers of associated lymphocytes, histiocytes, eosinophils, and rare multinucleated giant cells. Three of the eight Csk cKO mice (37.5%) had severe segmental acute fibrinonecrosuppurative and hemorrhagic enteritis with villar loss and intravascular fibrin thrombi (Figs. 1–4). Remaining crypts within and adjacent to the lesions of severe necrosuppurative enteritis were frequently hyperplastic. Two female Csk cKO mice and two male Csk cKO mice also had minimal to mild locally extensive suppurative gastritis in the squamous portion of their stomachs. The suppurative gastritis was associated with hemorrhage and edema of the mucosa, submucosa, and/or muscular layers of the stomach. Neutrophils were also present around blood vessels (Fig. 5). Three female Csk cKO mice and three male Csk cKO had scant to minimal multifocal suppurative colitis characterized by scant to minimal infiltration of neutrophils within the colonic lamina propria (with or without lymphocytes and histiocytes) and muscle layers and occasional crypt abscesses. The described gastrointestinal lesions and presumed associated decreased food intake may have been responsible for the observed body weight loss in these mice.
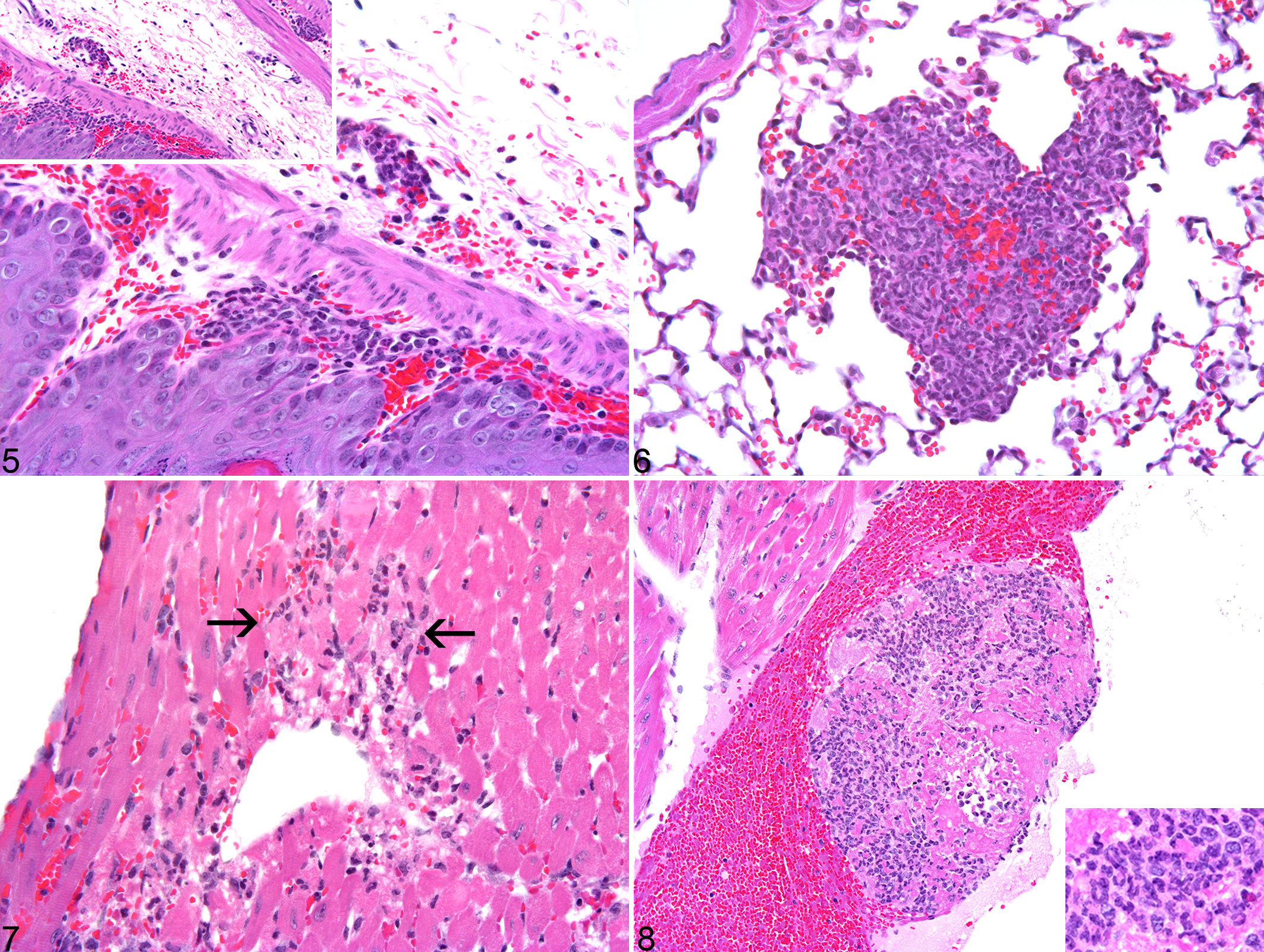
Summary of the Distribution and Incidence of Histologic Lesions in Csk Conditional Knockout (Csk cKO) Mice Treated With 3 Doses of Tamoxifen.
The data show the number of mice per group with lesions in each organ.
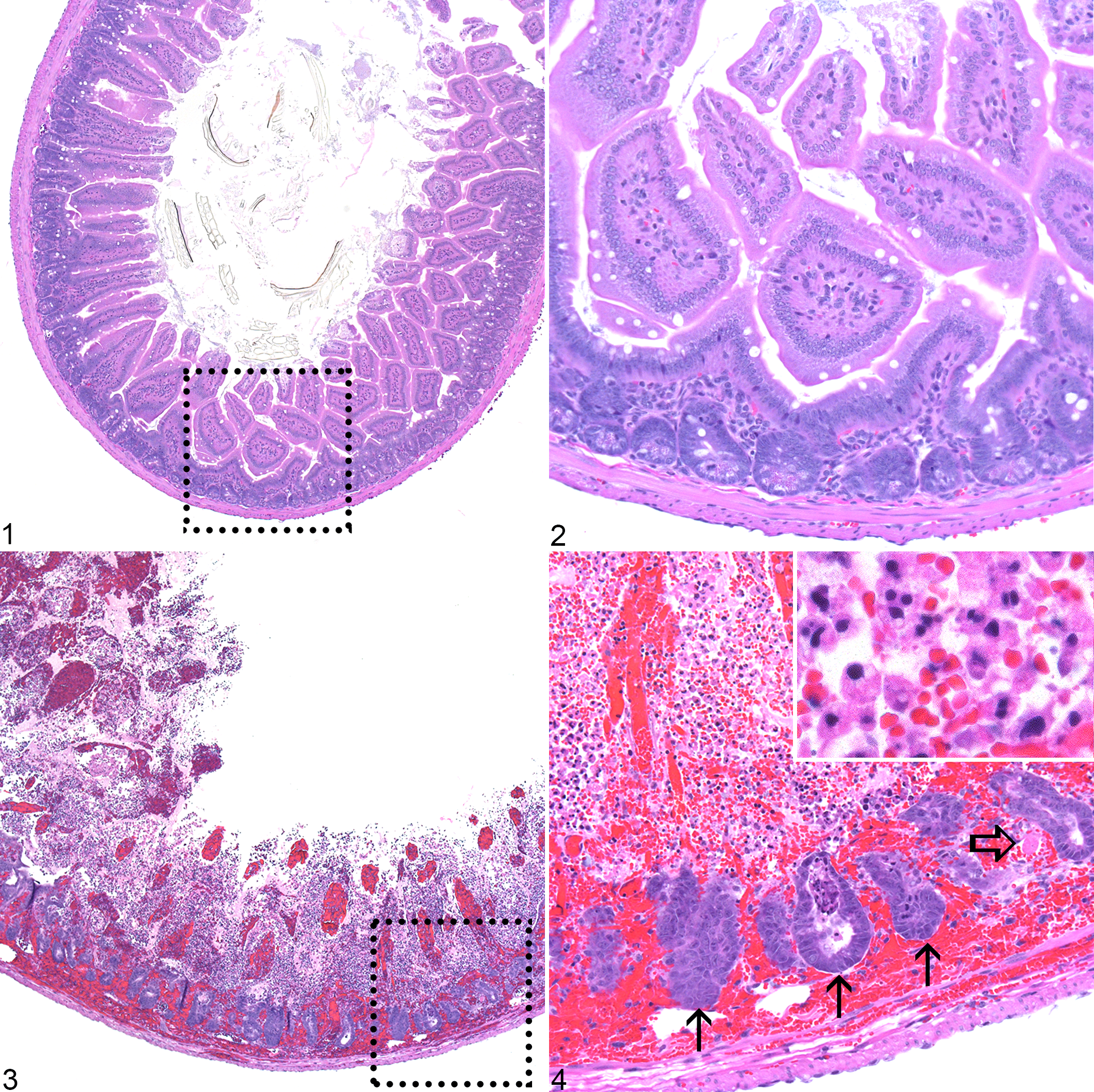
Two female Csk cKO mice and one male Csk cKO mouse had suppurative pneumonia with varying amounts of hemorrhage (Fig. 6). One male Csk cKO mouse had a mild locally extensive suppurative myocarditis with cardiomyocyte degeneration (Fig. 7). One female Csk cKO mouse had a thrombus within the lumen of the left ventricle. This thrombus was composed of strands of fibrin admixed with a myriad of neutrophils (Fig. 8). Three female and three male Csk cKO mice exhibited minimally increased numbers of inflammatory cells within their livers as well as an increase in extramedullary hematopoiesis. Hepatic inflammatory cell infiltrates were composed of varying numbers of neutrophils and histiocytes. There was no hepatocyte destruction associated with the presence of the inflammatory cells.
In addition to the above lesions, Csk cKO mice also exhibited increased medullary and extramedullary hematopoiesis (in addition to the hepatic extramedullary hematopoiesis), likely in response to the multiorgan inflammation. Three female and three male Csk cKO mice exhibited minimally increased splenic extramedullary hematopoiesis. Four female and three male Csk cKO mice had minimal to mild myeloid and megakaryocytic hyperplasia in the sternal bone marrow. One male Csk cKO mouse had minimal to mild megakaryocytic hyperplasia in the sternal bone marrow.
Three-day tamoxifen induction: clinical pathology
Blood was collected from the 8 Csk cKO and 8 control mice at necropsy (Table 1, column 2), and serum biochemical analyses were performed. Male and female Csk cKO mice had mildly elevated serum urea nitrogen compared to the age- and sex-matched control mice (2.1× and 1.4× fold increases, respectively, for the males and females) (Supplemental Table 2). This may have been secondary to bleeding into the gastrointestinal tract, since all mice with enteritis had varying amounts of associated intralesional hemorrhage. Triglycerides were also mildly elevated in the male and female Csk cKO mice compared to the control (2.3× and 3.6× fold increases, respectively, for the males and females) (Supplemental Table 2). Given the observed weight loss in both the male and female Csk cKO mice, the mild elevation in triglycerides could have been secondary to inappetance, although food intake was not monitored. Total protein and albumin were lower in the Csk cKO female mice and were 0.75× and 0.70× of the control means, respectively (Supplemental Table 2). This may have been due to a combination of presumed decreased food intake and gastrointestinal protein loss due to the enteritis.
Blood was collected from a cohort of male Csk cKO and control mice for hematology (Table 1, column 3) (n = 5 mice/genotype). Csk cKO male mice had mildly lower white blood cell counts (0.76× control mean) compared to the control mice, primarily due to decreased neutrophils (0.59× control mean) (Supplemental Table 3). Platelet counts were moderately decreased in the Csk cKO male mice (0.49× control mean) (Supplemental Table 3). These decreases were likely secondary to margination, sequestration, and utilization in the areas of tissues inflammation and hemorrhage and not due to myelosuppression, since the bone marrow demonstrated myeloid and megakaryocytic hyperplasia.
Csk cKO Mice Treated With 1 IP Injection of Tamoxifen
One-day tamoxifen induction: gene excision
Gene excision was evaluated from two cohorts of Csk cKO mice that received a single IP injection of tamoxifen (Table 1, columns 4 and 5). The Csk gene excision levels from these two cohorts of male Csk cKO mice were, as anticipated, less than those observed in the cohorts that had received three sequential doses of tamoxifen. The first cohort (Table 1, column 4) consisted of 15 male Csk cKO mice and 15 male control mice that received a single IP injection of tamoxifen in 10% ethanol/ 90% sunflower seed oil and were euthanized on day 4, 7, or 14 (n = 5 mice/genotype/time point). Spleen, lymph nodes, bone marrow, and skin were collected for gene excision analysis via Southern blot (Table 3). On day 4, gene excision ranged from 64% in the skin to 93% in the bone marrow (ranked by decreasing order of gene excision level: bone marrow > spleen > lymph nodes > skin). On day 7, gene excision ranged from 46% in the skin to 91% in the bone marrow (bone marrow > spleen > lymph nodes > skin). On day 14, gene excision ranged from 38% in the skin to 66% in the spleen (spleen > bone marrow ≈ lymph nodes > skin).
Csk Gene Excision Level After a Single Dose of Tamoxifen (n = 5 Mice/Day)a.
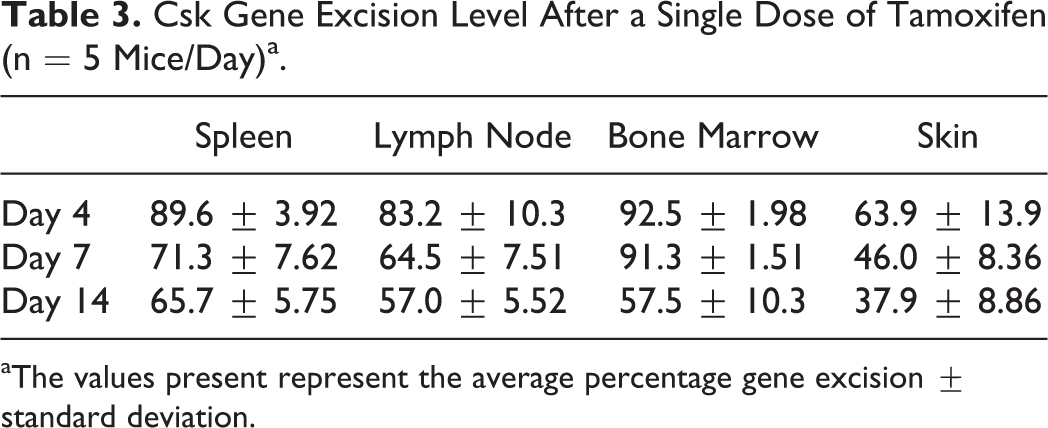
aThe values present represent the average percentage gene excision ± standard deviation.
The second cohort consisted of 5 male Csk cKO mice and 5 male control mice that received a single IP injection of tamoxifen in 10% ethanol/ 90% sunflower seed oil and were weighed daily for the following 15 days (Table 1, column 5). Gene excision on day 16 was confirmed via southern blot analysis of the spleen, lymph nodes, bone marrow, and skin with average Csk gene excision of 56.7% ± 10.5%, 46% ± 9%, 65.1% ± 7.7%, and 35% ± 8.4% in the spleen, lymph nodes, bone marrow, and skin, respectively (data presented as average ± standard deviation). Tissue differences in Csk gene excision efficiency in both of these cohorts of mice (with the lowest level of Csk gene excision observed in the skin) may have been due to tissue differences in tamoxifen exposure 18 and/or differences in chromatin structure which could affect the accessibility of LoxP sites to Cre. 21
One-day tamoxifen induction: body weight
Five male Csk cKO mice and five control mice received a single IP injection of tamoxifen in 10% ethanol/ 90% sunflower seed oil and were weighed daily for the following 15 days (Table 1, column 5). Mouse weights remained steady over the 16 days evaluated, and Csk gene excision was not associated with significant weight loss or gain, any clinical signs, or gross lesions (data not shown).
One-day tamoxifen induction: histopathology
Fifteen male Csk cKO mice and fifteen control male mice received a single IP injection of tamoxifen and were euthanized and necropsied on day 4, 7, or 14 (Table 1, column 4, n = 5 mice/genotype/time point). At all time points examined, the Csk cKO and control mice exhibited varying degrees of peritonitis, mesenteric steatitis, and or serositis that ranged from lymphohistiocytic to suppurative. This inflammation was presumed to be secondary to the oil in the tamoxifen vehicle (10% ethanol/ 90% sunflower seed oil) since peritonitis secondary to usage of oil has been documented. 9,22
Significant histologic lesions were identified in the day 14 Csk cKO mice. All (5/5) Csk cKO mice had widespread interstitial suppurative to pyogranulomatous pneumonia (affecting the interstitium, terminal airways, and blood vessels) consisting of coalescent areas and individual nodules of neutrophils and macrophages with fibrin, hemorrhage, and reactive hyperplasia of type II pneumocytes and fibroblasts (Figs. 9–11). In addition to the interstitial pneumonia, 5/5 of the Csk cKO mice (100%) had minimal to mild suppurative enteritis characterized by a neutrophil-rich infiltrate in the lamina propria of their small intestines with rare multinucleated giant cells. All five of the Csk cKO mice (100%) had minimal to mild myeloid hyperplasia in the bone marrow and 3/5 of the Csk cKO mice (60%) had minimally to mildly increased numbers of neutrophils, lymphocytes, and histiocytes within their livers compared to the controls. Mild multifocal suppurative dermatitis with subcorneal pustule formation was identified in the haired skin from the pinna of 1/5 Csk cKO mice.
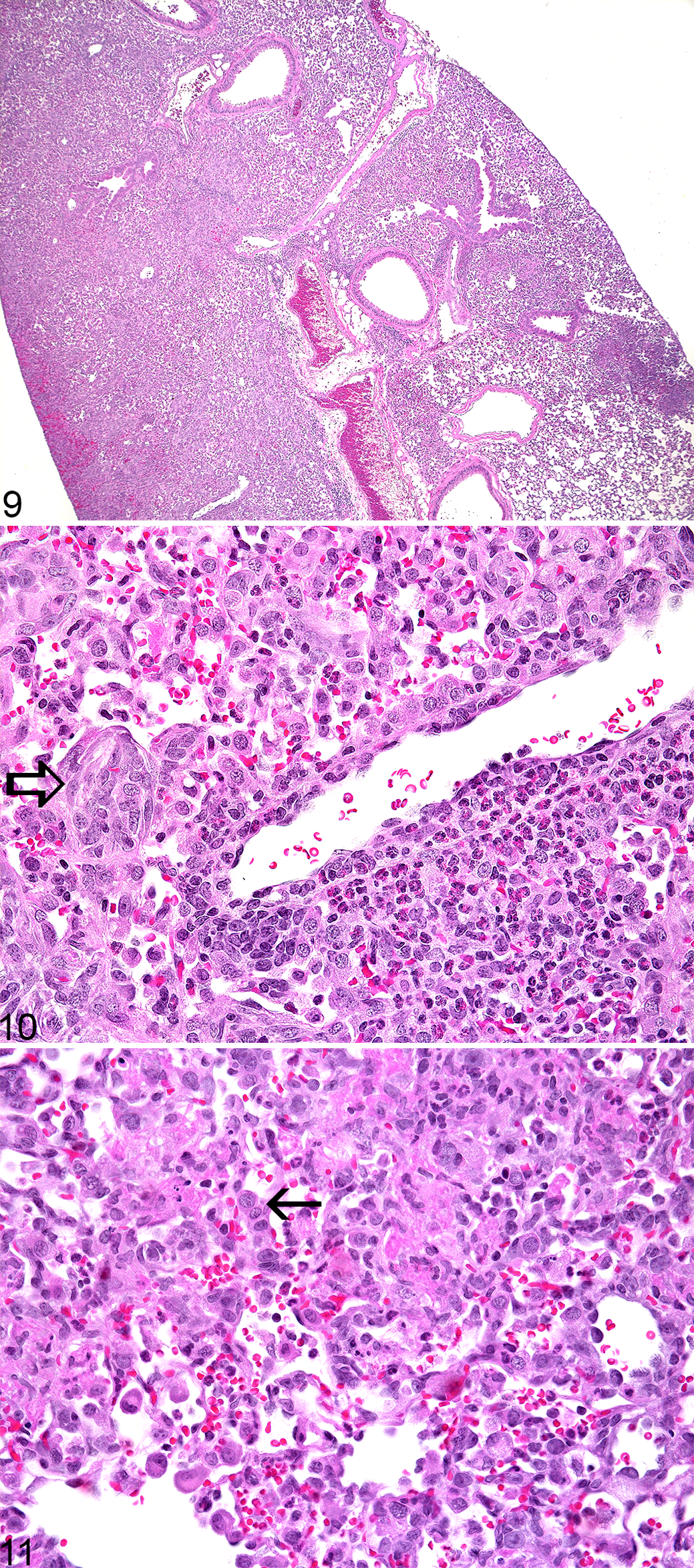
Lung lesions that were similar to but less severe and less extensive than those identified in the day 14 mice were found in 2/5 of the Csk cKO mice (40%) on day 7. There was no histologic evidence of enteritis at the day 7 time point. All 5 Csk cKO mice necropsied on day 7 had minimally to mildly increased lymphohistiocytic (with or without neutrophils) infiltration in their livers compared to the control mice. None of the Csk cKO mice had skin lesions, but intracorneal pustule formation was identified in the skin from the pinna of 2/5 of the control mice. Aside from the peritonitis described above, there were no significant histologic lesions identified in the Csk cKO or control mice euthanized on day 4, with the exception of minimally increased numbers of lymphocytes, neutrophils, and histiocytes in the liver of one mouse and mild, focal periportal to midzonal suppurative hepatitis with hepatocellular necrosis in one mouse. As with the day 7 time point, none of the Csk cKO mice had skin lesions. However, intracorneal pustule formation was observed in the skin from the pinna of 2/5 of the control mice.
Discussion
In this paper we showed that systemic loss of Csk activity, via use of Cre/LoxP gene excision technology, resulted in suppurative inflammation in multiple organs. Notable affected organs included the gastrointestinal tract, lung, and, to a lesser extent, heart. Minimally increased hematopoiesis, likely as a response to systemic recruitment and utilization of neutrophils, was present within the bone marrow, liver, and spleen. Importantly, the results from the two induction studies (3 days versus 1 day induction with tamoxifen) were similar to each other, supporting the robustness and reproducibility of these observations. Although reducing the tamoxifen dosing interval from three days to one day reduced the degree of Csk gene excision and eliminated the weight loss observed in the former group, the two groups of mice exhibited comparable histopathologic lesions.
The results of the current study were largely in agreement with previous work in mice engineered to have granulocyte-specific loss of Csk activity, with those mice developing suppurative pneumonia. 19 However, the Csk cKO mice in the present study appeared to be the first model of Csk loss in which inflammatory lesions were found in the gastrointestinal tract and heart. It is unknown if hearts from the granulocyte-specific Csk-knockout mice were evaluated histologically, but the authors state that there were no significant lesions in the gastrointestinal tracts. 19 Interestingly, the lung, which was a target of inflammation after Csk gene excision induced by either one day or three days of tamoxifen treatment, appeared to be a common site for inflammation associated with aberrant SFK activation 19 and is likely due to loss of inhibition mediated by Csk. In addition to the lung lesions present in mice with granulocyte-specific loss of Csk, 19 mice engineered to have a constitutively active mutant Hck (a SFK found in granulocytes, monocytes, and macrophages, which is resistant to inhibitory phosphorylation by Csk), develop pneumonia that is granulocyte- and macrophage-rich. 2 Although suppurative dermatitis, including suppurative otitis externa and blepharitis, have been described in mice engineered to have granulocyte-specific loss of Csk, 19 this finding was not recapitulated in the Csk cKO mice in the present study. Mild multifocal suppurative dermatitis was identified in the haired skin of the pinna of one male Csk cKO mouse treated with a single IP injection of tamoxifen and euthanized on day 14. This lesion, however, was unlikely to be related to Csk gene excision, since inflammation, as manifested by intracorneal pustules, was identified in the pinnae from four control mice (two euthanized on day 4 and two euthanized on day 7), and, therefore, was likely incidental. Histological examination of skin from more locations on the body would have been useful to discern whether systemic conditional Csk excision was associated with dermal inflammation.
Our findings, described for the first time in an inducible Csk knock-out model, are consistent with previous reports highlighting the importance of Csk in regulating neutrophil activation, adhesion, and degranulation in vivo. 2,19 Specifically, the similarities between the inflammatory lung lesions in the present study to those described in granulocyte-specific Csk-knockout mice 19 and in mice with mutated Hck 2 suggest that the inflammatory lesions in Csk cKO mice may be due to exuberant neutrophil activation and dysregulation likely due to loss of Csk’s inhibitory activity on proinflammatory SFKs.
As previously discussed, the lung, gastrointestinal tract, and to a lesser extent, the heart were identified as target organs for loss of Csk activity in the present study, and pneumonia is a common sequela of loss of Csk regulation of SFKs. 2,19 It has been posited that the pulmonary inflammation in mice with granulocyte-specific loss of Csk was in response to the normal bacterial flora in the lungs, since knockout mice derived under specific pathogen free conditions or given antibiotics did not develop pneumonia. 19 Although the Csk cKO mice in the current study were specific pathogen free, they were not germ-free. Therefore, an exuberant and inappropriate reaction to normal resident microflora could explain why the inflammatory response affected the lung and gastrointestinal tract after Csk gene excision (although neither Gram stains nor blood cultures were performed in the current studies). The minimal increase in the number of inflammatory cells in the livers of the Csk cKO mice may have been in reaction to translocation of bacteria from the gastrointestinal tracts to the liver. The left ventricular thrombus (composed of fibrin and neutrophils) in one female Csk cKO mouse may have been secondary to hypercoagulability due to systemic inflammation as a result of aberrant neutrophil activation or due to intestinal protein loss (Supplemental Table 1).
Interestingly, although the histopathologic findings in the two different tamoxifen induction studies were qualitatively similar, the severity and extensiveness of the gastrointestinal tract and lung lesions varied depending on whether Csk excision was induced with either 3 or 1 days of IP tamoxifen: mice induced with 3 days of tamoxifen had more severe gastrointestinal lesions and less severe lung lesions than mice that received a single dose of tamoxifen. The gastrointestinal tract being more severely affected on days 10 and 11 (7–8 days after 3 consecutive IP doses of tamoxifen) compared to after a single IP injection of tamoxifen may be explained by a difference in sustained gene excision levels. Notably, the Csk cKO mice that received 3 doses of tamoxifen had higher gene excision on days 10 and 11 than mice that had received 1 dose of tamoxifen and were evaluated histologically on days 4, 7, or 14. The greatest differences in Csk excision levels were evident when the 3-day induction mice were compared to the mice that had received a single dose of tamoxifen and were euthanized on day 7 or 14.
Regarding lung lesion severity, Csk cKO mice that had received a single IP dose of tamoxifen and were examined on day 14 had the most severe lung lesions despite the fact that gene excision levels were lowest at this time point. Lung lesions in the Csk cKO mice that received a single dose of tamoxifen and were euthanized on day 7 (2/5 mice) were more similar in severity to the lung lesions present in the Csk cKO mice that received three IP injections of tamoxifen and were euthanized on day 10 or 11 (3/8 mice). Possibly, with time, the pneumonia in the Csk cKO mice induced with 3 doses of tamoxifen could have progressed to the severity of the pneumonia present in Csk cKO mice 14 days after a single injection of tamoxifen. It would have been informative to evaluate the lungs of Csk cKO mice induced with three doses of tamoxifen at later time points, however, that experiment was not possible given the progressive weight loss.
The usefulness of doing time course studies with induced knockouts, such as the Csk cKO mice, was evident from the current study. Lung lesion progression was clearly apparent when the lungs of Csk cKO mice that had received one IP injection of tamoxifen and euthanized on day 7 were compared to the lungs of the mice euthanized on day 14. Of note, the evaluation of a single early time point could have missed the lesions, since Csk cKO mice euthanized on day four had no significant lung or gastrointestinal tract lesions. The delay in histologically apparent inflammatory lesions may have been due to the fact that cellular turnover of functional Csk was necessary before the proinflammatory effects of Csk gene excision could become apparent and inflammatory lesions develop.
Although hyperplasia has been previously described in mice with either skin-specific 6,23 or enterocyte-specific 7 loss of Csk, the only hyperplasia present in the Csk cKO mice was regenerative in response to local inflammation (ie, type II pneumocyte and fibroblast hyperplasia in the lungs with severe pneumonia and crypt epithelial hyperplasia in the mice with fibrinonecrosuppurative and hemorrhagic enteritis). Hyperplasia was not identified in any organs without inflammation. The question of whether the Csk cKO mice would develop hyperplasia unassociated with significant tissue injury if they were given more time with continuous gene excision remains to be addressed.
In conclusion, systemic loss of Csk in adult mice, when induced with either one or three doses of tamoxifen IP, resulted in suppurative inflammation in multiple organs, likely as a result of aberrantly increased neutrophil activation. The gastrointestinal tract and lung were the organs most consistently and severely affected by loss of Csk activity, although lesions were also present in the heart and there was an increased number of inflammatory cells in the liver. The lesions that Csk cKO mice develop further illustrate the importance of Csk in limiting SFK-mediated inflammation and highlight the potential hazards of systemic inhibition of Csk.
Footnotes
Acknowledgements
We thank Karen Granaldi and Christopher Hasson of the Discovery Toxicology Histology Laboratory at Bristol-Myers Squibb for their invaluable histology support. We thank Lois Lehman-McKeeman for her helpful comments and review of the manuscript. We thank Jian Chen and Doreen Hewston for in vivo knockout coordination and John Megill for his help with the figure layout.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
These studies were supported by Bristol-Myers Squibb.
Supplementary material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.