
Abstract
Feline primary cutaneous lymphomas (FPCLs) account for 0.2% to 3% of all lymphomas in cats and are more frequently dermal nonepitheliotropic small T-cell tumors. Emergence of FPCL seems unrelated to feline leukemia virus (FeLV) serological positivity or to skin inflammation. A total of 17 cutaneous lymphomas with a history of vaccine injection at the site of tumor development were selected from 47 FPCLs. Clinical presentation, histology, immunophenotype, FeLV p27 and gp70 expression, and clonality were assessed. A majority of male (12/17), domestic short-haired (13/17) cats with a mean age of 11.3 years was reported. Postinjection time of development ranged from 15 days to approximately 9 years in 5 cats. At diagnosis, 11 of 17 cats had no evidence of internal disease. Lymphomas developed in interscapular (8/17), thoracic (8/17), and flank (1/17) cutaneous regions; lacked epitheliotropism; and were characterized by necrosis (16/17), angiocentricity (13/17), angioinvasion (9/17), angiodestruction (8/17), and peripheral inflammation composed of lymphoid aggregates (14/17). FeLV gp70 and/or p27 proteins were expressed in 10 of 17 tumors. By means of World Health Organization classification, immunophenotype, and clonality, the lesions were categorized as large B-cell lymphoma (11/17), anaplastic large T-cell lymphoma (3/17), natural killer cell–like (1/17) lymphoma, or peripheral T-cell lymphoma (1/17). Lineage remained uncertain in 1 case. Cutaneous lymphomas at injection sites (CLIS) shared some clinical and pathological features with feline injection site sarcomas and with lymphomas developing in the setting of subacute to chronic inflammation reported in human beings. Persistent inflammation induced by the injection and by reactivation of FeLV expression may have contributed to emergence of CLIS.
Lymphomas represent more than 50% of all tumors in cats, with a prevalence of approximately 1.6% of the general feline population and 4.7% of hospitalized sick cats. 34 Most common sites include the intestine and mediastinum. 34 Primary cutaneous lymphomas account for 0.2% to 3% 25,29,54,71 of all feline lymphomas.
While epitheliotropic lymphomas of the skin account for up to 44% of primary cutaneous lymphomas in dogs and humans, 21,27,31 epitheliotropism is rarely observed in cats. Nonepitheliotropic lymphomas are more frequent in cats 27,54 and include indolent T-cell lymphoma, also referred to as cutaneous lymphocytosis; 23,24 diffuse T-cell lymphoma; T-cell-rich large B-cell lymphoma; 15,27 and lymphoplasmacytic lymphoma. 37 Similar to other species, cutaneous lymphomas mostly have a T-cell origin 21,27 while B-cell tumors are considered extremely rare. 14,27
In addition to viral infections, such as Epstein-Barr virus (EBV) and human T-lymphotropic virus type 1 (HTLV-I), 32,73 chronic inflammation is generally accepted as a risk factor for the development of hematopoietic malignancies, 20,26,59 specifically cutaneous T- and B-cell lymphomas in humans. 6,20,59
The overall incidence of feline leukemia virus (FeLV) in cats with lymphomas has decreased from 70% 33 to 15% in North America. 43 A similar drop in prevalence, from 59% to 13% between 2 consecutive 15-year periods, has been reported in Germany. 51 FeLV prevalence in the 1970s was estimated at less than 10% in cats with cutaneous lymphoma. In Italy, seroprevalence evaluated in high-risk (stray cats) and sick cats has decreased from 18% to 3.8%, 4,64 while no data are available regarding cats with lymphoma. Therefore, this excludes a major role of progressive FeLV infection in feline cutaneous lymphoma development. Progression of chronic inflammation to lymphoma is not yet confirmed in cats. However, the development of sarcomas at injections sites (eg, rabies vaccine, long-acting antibiotics or steroids, 38 or the benzoylurea pesticide lufenuron 18 ), at sites of implanted foreign material (nonabsorbable suture material, 5 microchip implants, 7,12 retained surgical sponges 28 ), or trauma 17,22,63 are well documented and thought to be attributed to the chronic inflammation elicited. 39 Moreover, cutaneous lymphomas have arisen in areas of feline injection site sarcomas subsequent to chemotherapy or radiation therapy of the primary tumor, and lymphomagenesis in these cases was hypothesized to be due to the mutagenic action of chemotherapy or radiation treatment. 44
In a subgroup of recently reported primary feline subcutaneous lymphomas, site of development (lateral thorax and interscapular region), clinical presentation as a single nodule, presence of necrosis leading to central cavitation, and peripheral inflammation were considered clinically highly suggestive of feline injection site sarcomas. 52 The aim of this report is to describe pathological features of a series of primary feline cutaneous lymphomas, which developed in sites of previous injections and were not preceded by sarcomas.
Materials and Methods
Tissue Samples and Clinical History
Forty-seven cases of feline primary cutaneous lymphomas were submitted to the diagnostic histopathology service at the School of Veterinary Medicine of Milan from 2001 to 2012. Samples were submitted by veterinary practitioners or as second opinion cases from other veterinary schools and private diagnostic laboratories throughout Italy. Punch or excisional biopsies of skin nodules or formalin-fixed, paraffin-embedded tissue blocks were submitted.
Clinical data, including signalment, tumor site, previous history of injection at the tumor site, clinical staging, therapy, and follow-up information, were received upon submission or collected by contacting the referring veterinarian after the case selection according to histomorphology suggestive of injection/vaccination or in cases where injection/vaccination was suspected by the referring veterinarian. Based on histopathological features and clinical information, 17 of 47 primary cutaneous lymphomas were included in this study.
Histopathology
Tissue specimens received fixed in 10% neutral buffered formalin were routinely processed, and tissue blocks (from the diagnostic service or from other institutions) were used to obtain multiple 4- to 6-μm thick sections. Morphologic features were evaluated on tissue sections stained with hematoxylin and eosin stain. The following parameters were recorded: site and extension of the neoplastic lesion; presence of angiocentric, angioinvasive, and angiodestructive behavior; cell morphology and cell size; number of mitoses; necrosis; mineralization; and inflammation, including reactive lymphoid nodular aggregates. The mean mitotic index (MI) per high-power field (HPF; 400× magnification) was evaluated by dividing by 10 the mitotic count obtained in 10 consecutive HPFs in areas having the highest number of mitotic figures. The MI was calculated independently by 3 pathologists (P.R., G.A., and C.G.) using the same microscope, and a mean of the 3 readings was used to calculate the MI per field. According to the MI, a low grade was assigned to cases with a mean of 0 to 6 mitoses, medium grade to cases with a mean of 6 to 10 mitoses, and a high grade with a mean over ≥11. 70 Definitive diagnoses were obtained according to the veterinary World Health Organization (WHO) classification of lymphomas. 70,72
Immunohistochemistry
Detailed information about the panel of primary antibodies is listed in Supplemental Table S1. Cross-reactivity of anti–human CD3-∊, CD79a, CD45RA, and CD20 with feline tissues has been previously demonstrated. 1,13,23,36,48 Paraffin sections (4–6 μm) were mounted onto poly-L-lysine–coated slides, deparaffinized in xylene, and hydrated through graded ethanol solutions. Endogenous peroxidase was quenched with hydrogen peroxide (0.3%) and sodium azide (0.1%) in Tris buffer (0.1-M solution, pH 7.5) for 30 minutes and rinsed in 3 changes of Tris buffer for 5 minutes each. For heat-induced antigen retrieval, slides were immersed in 10 mM citrate buffer (pH 6.0) (DAKO, Carpinteria, CA) and microwaved at maximum power for 1 minute and twice at 750 watts for 3 minutes. After cooling the slides down to room temperature for 20 minutes, unspecific staining was blocked with 10% heat-inactivated horse serum in Tris buffer for 30 minutes. Primary antibodies were applied at the specific working concentrations (Suppl. Table S1) diluted in Tris buffer containing 10% inactivated serum and were incubated in a humidified chamber at 37°C for 1 hour. Secondary detection was performed with the avidin-biotin enzyme complex (ABC kit, Vectastain; Vector Laboratories, Burlingame, CA, USA) for 30 minutes. The reaction was developed with the peroxidase amino-9-ethyl-carbazole (AEC) substrate kit (Dako, Glostrup, Denmark). Smears were counterstained with Mayer’s hematoxylin for 3 minutes and cover-slipped with an aqueous mounting medium (Glycerol; Sigma-Aldrich, St Louis, MO). Negative controls consisted of substitution of specific antibodies with an isotype-matched, irrelevant monoclonal antibody or omission of the primary antibody. Sections of feline reactive peripheral submandibular lymph node were used as positive controls. For FeLV antibodies, formalin-fixed sections from a lymph nodal lymphoma with FeLV p27- and gp70-positive cells from a FeLV enzyme-linked immunosorbent assay (ELISA) p27 serologically positive cat were used as positive controls.
Clonality Assessment
For DNA extraction, four 25-μm paraffin sections were collected in an Eppendorf tube. Blades were changed and microtome was cleaned after each case to avoid DNA cross-contamination. The sections were deparaffinized in xylene and washed twice in 100% ethanol. Genomic DNA was extracted using the DNAeasy Blood and Tissue Kit (Qiagen, Valencia, CA) following the manufacturer’s instructions. The concentration of genomic DNA was measured using an Introspect 2100 pro spectrophotometer UV/Visible spectrophotometer (Amersham Pharmacia Biotech, Uppsala, Sweden). Polymerase chain reaction (PCR) analysis for T-cell and B-cell clonality assessment was performed as previously described. 55,74 A single primer pair was used for assessment of T-cell receptor gamma (TCRG), as previously designed and described. 55 An approximately 110–base pair (bp) segment (±20 bp) of the TCRG locus was amplified, using the forward primer in the variable region (5′–3′: GAA GAG CGA YGA GGG MGT GT, Y: C or T; M: A or C) and the reverse primer in the joining region (5′–3′: CTG AGC AGT GTG CCA GSA CC, S: C or G). Two segments of the immunoglobulin heavy chain (IgH) locus were amplified to assess B-cell clonality. 23,74 One upper (forward) primer (5′–3′: CCA GGC TCC AGG GAA GGG) was paired with 2 lower primers (5′–3′: TGA GGA CAC TGT GAC TAT GGT TCC and GGA CAC CGT CAC YAK GVY TC, Y: C or T; K: G or T; V: A, C or G) to amplify framework 2 (an approximately 250-bp segment). The same lower primers were paired with upper primer (5′–3′: CTC CGT GAA GGG CCG ATT) to amplify framework 3 (an approximately 180-bp segment). Each reaction was performed with 100 ng genomic DNA. A reaction mixture was used as previously described, with 2-step touch-down amplification conditions applied. 23,74 The polyclonal control was composed of genomic DNA extracted from feline peripheral blood mononuclear cells; DNA extracted from feline lymph nodes with confirmed T-cell lymphoma or B-cell lymphoma were used as clonal controls.
For 6 samples (cases Nos. 1–6), PCR reactions (prior to summer of 2007) were run in duplicates to confirm results. Native samples were used directly for gel electrophoresis. In addition, heteroduplex analysis, previously described to assist separation of true clonal from false-positive results, was performed for all samples. 23,74 Native PCR products (10 μl) were denatured at 95°C for 10 minutes, then allowed to reanneal at 4°C for 1 hour prior to gel electrophoresis. Duplicate PCR samples (10 μl) of each native and heteroduplex sample, including controls (negative, polyclonal, and clonal), were analyzed by gel electrophoresis using precast 10% nondenaturating polyacrylamide Tris-Borate-EDTA (TBE) gels (Criterion Pre-cast gels; Bio-Rad, Hercules, CA). Gels were subsequently stained with Gel Star nucleic acid stain (Cambrex Bioscience Rockland, Rockland, ME) as previously described. Bands and smears were visualized on a UV transilluminator and photographed.
For 11 samples (cases Nos. 7–17), PCR products (after summer 2007) were processed using the HDA-G12 Genetic Analyzer by eGene. Triplicate native samples (10 μl), including controls (negative, polyclonal, and clonal), were analyzed using BioCalculator software and eGene’s DNA Gel Cartridge Kit (eGene, Irvine, CA, USA). BioCalculator analysis method OL700 was used for 2- to 5-bp resolution between products of 100 and 500 bp.
Results
Clinical History and Signalment
Based on histopathological features and clinical information, 17 of 47 primary cutaneous lymphomas were included in this study. All 17 cases were confirmed to have arisen at sites of previous vaccine injection by the referring veterinarian and presented either as a single nodule or localized disease. Of the 30 cases not included, 24 developed at noninjection cutaneous sites, while in 6 cases, site and history were not retrieved; only one of these had microscopic features resembling those described in this caseload but was not included. All cats were vaccinated with a combination vaccine including panleukopenia virus, feline herpesvirus 1, and feline calicivirus (FHV-1/FCV). The vaccines came from 6 different manufacturers. FeLV vaccination was administered to 5 of 17 cats. No rabies vaccines were administered, since Italy is a country free of rabies. Exact time span between vaccination and development of lymphoma was available for 5 of 17 cats: lesions developed 15 days after last FeLV vaccination in cats Nos. 3 and 6, 1 year after the last annual routine vaccination in cat No. 17, approximately 5 years after the last vaccination in cat No. 14 (no additional injections were given), and approximately 9 years after the last vaccination in cat No. 16.
Signalment of cats and clinical information, including size, site, number, and microanatomical extension (evaluated histologically) of lesions (when available), are summarized in Supplemental Table S2. Most were domestic short-haired cats (13/17). Mean age was 11.3 years; 14 of 17 cats (81%) were 10 years or older. Male neutered cats were 12 of 17 (71%), and 5 of 17 cats (30%) were female spayed; the male to female ratio was 2.4. Serology for feline immunodeficiency virus was negative for 6 of 17 cats, and 8 of 17 cats tested negative for FeLV.
At time of diagnosis, none of the cats had systemic clinical signs. Most cats (14/17) had a solitary nodular skin lesion. The remaining 3 of 17 cats had either 2 nodules connected by a linear mass or regional disease characterized by multiple nodules limited to a 5-cm area or 2 adjacent nodules. Pruritus, ulceration, and pain were absent with the exception of 2 of 17 cats, which both presented with ulcerated lesions at first surgery.
Initial clinical staging by thoracic radiographs, abdominal ultrasound, or total-body computed tomography (CT) scan and blood analysis was available in 11 of 17 cats, all of which were confirmed negative for internal disease. In 3 of 17 cats (cat Nos. 2, 6, and 13), the presence of lymphoma limited to the skin was confirmed at necropsy.
Microscopical Features
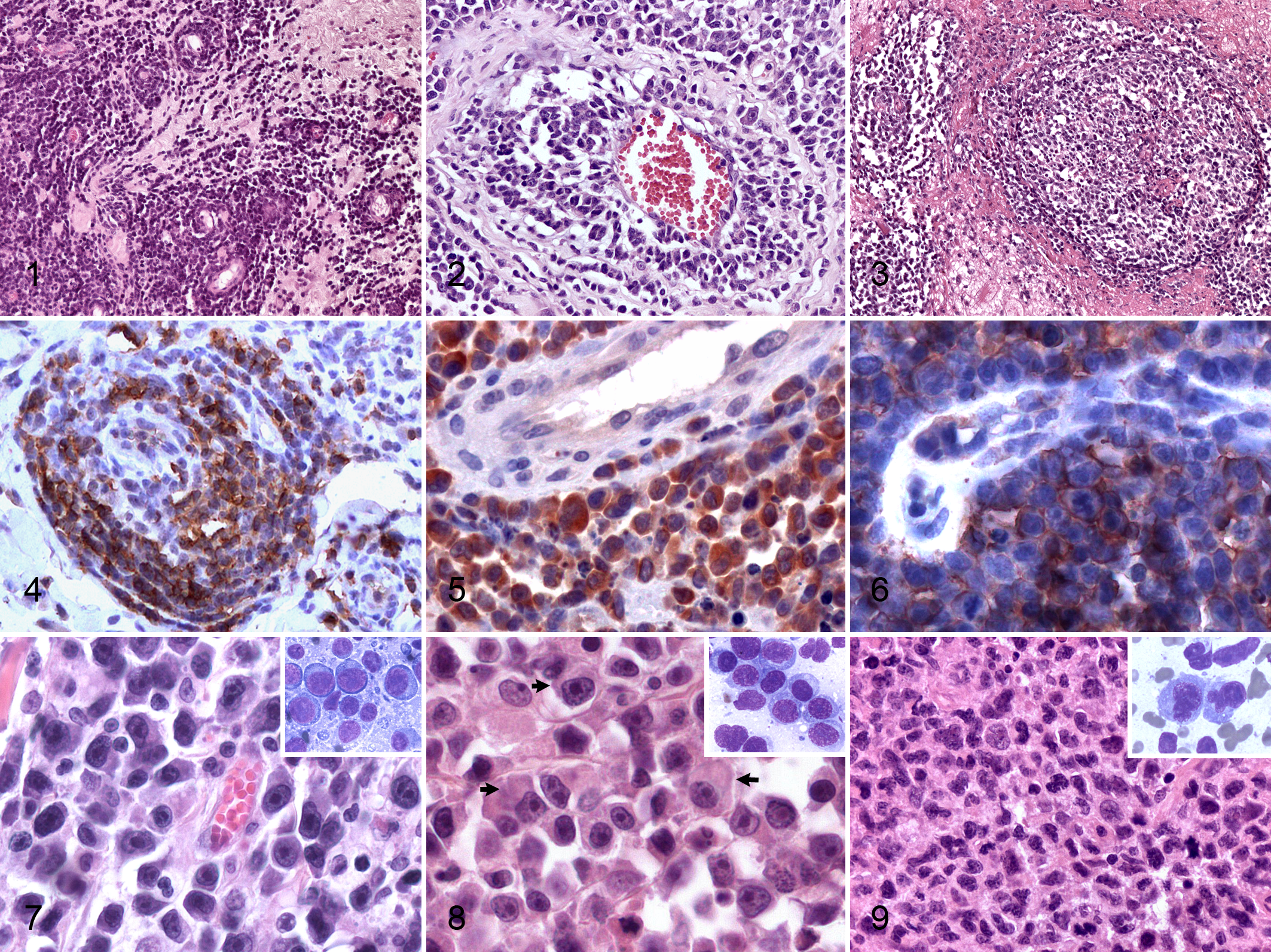
Incisional biopsies were available for all 17 cases included in the study. Main microscopic findings are listed in Table 1. In 4 of 17 cats, cytological specimens were also available and contributed to the diagnosis and classification of tumors. Compared with the other primary cutaneous lymphomas (not included in the study), morphology of the lesions was distinctive and consisted of a poorly circumscribed, nonencapsulated, infiltrative mass that variably extended from the superficial to the deep dermis to the subcutis (Suppl. Table S2). A Grenz zone was always present, and epidermal and adnexal tropism was never observed. Typically, cells were organized in dense sheets and in perivascular aggregates, commonly associated with extensive areas of necrosis (16/17 cats). Neoplastic lymphoid cells tightly surrounded vessel walls (angiocentrism) in 13 of 17 cases (Fig. 1). In 10 of 17 cats, angioinvasion was observed (Fig. 2). Angiodestruction, a feature associated with vascular wall disruption and variable mural hemorrhages, fibrin deposition and luminal occlusion were recorded in 8 of 17 cats (Fig. 3). Central microcavitation with fibrin accumulation and the presence of occasional reactive macrophages and nondegenerated neutrophils were present in most cases. Foreign material was not found in the necrotic cavities or in macrophages.
Histopathological Features of Feline Primary Cutaneous Lymphomas at Injection Sites.
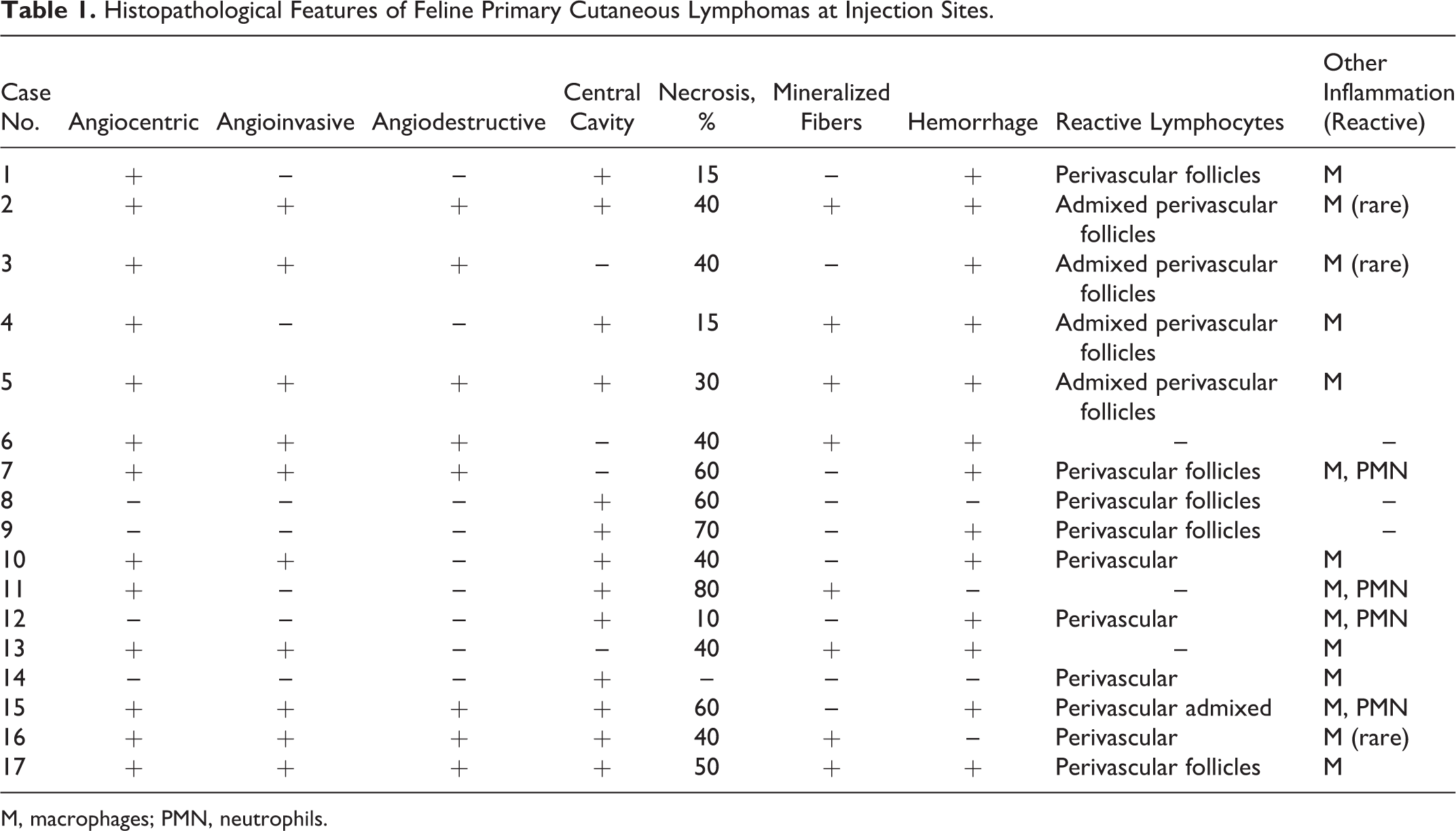
M, macrophages; PMN, neutrophils.
Inflammatory cells (lymphocytes and reactive macrophages) were frequently located at the periphery of the lesions or admixed with neoplastic cells as well as within and surrounding the central cavity. Perivascular aggregates of small mature lymphocytes were observed at the margins of the main tumor in 14 of 17 cats.
Complete excision was observed in 7 cases. Of the 7 cases that were completely excised, 3 recurred (cat Nos. 1, 2, and 14), 1 did not recur (cat No. 15), and no information was available for 3 cases.
Cytological features of neoplastic cells, including cell size, nuclear shape, chromatin pattern, number and evidence of nucleoli, and mitotic index, are summarized in Supplemental Table S3. According to the MI, in 2 of 17 cats, large cell lymphomas were of intermediate grade. In the remaining 15 of 17 cats, lymphomas were low grade.
Immunohistochemistry
Detailed immunohistochemical results are listed in Table 2. Expression of CD18 confirmed leukocyte origin of all 17 lesions examined. B-cell lymphoma was considered in 10 of 17 cats based on expression of CD20 (Fig. 4) and CD79a. Strong CD3 expression (Fig. 5) by neoplastic cells in 5 of 17 cats was consistent with T-cell lymphoma. Neither CD3 nor CD79a and CD20 were expressed in 2 of 17 cats. Nodular and follicular lymphoid aggregates of small lymphocytes were mixed B cells (CD79a) and T cells (CD3), and perivascular reactive small mature lymphocytes were consistently CD3 positive, interpreted as reactive lymphoid infiltrates. The neoplastic cells expressed FeLV gp70 and p27 antigens (5 cats), FeLV gp70 only (4 cats), or FeLV p27 only (1 cat). No expression was observed in tumor cells of 7 cats (Table 2).
Immunohistochemistry, Clonality, and Definitive Diagnoses of Feline Primary Cutaneous Lymphomas at Injection Sites.
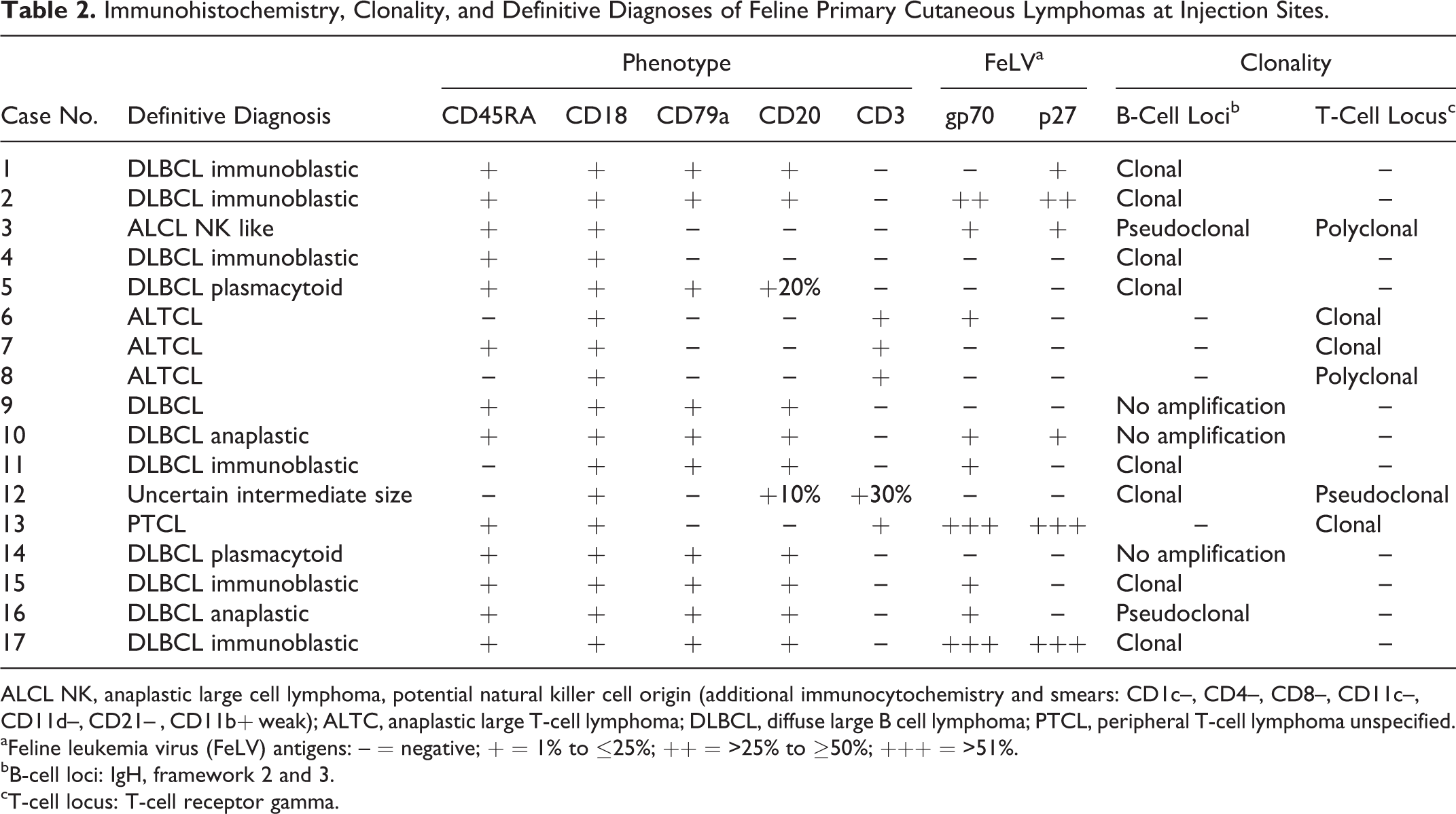
ALCL NK, anaplastic large cell lymphoma, potential natural killer cell origin (additional immunocytochemistry and smears: CD1c–, CD4–, CD8–, CD11c–, CD11d–, CD21– , CD11b+ weak); ALTC, anaplastic large T-cell lymphoma; DLBCL, diffuse large B cell lymphoma; PTCL, peripheral T-cell lymphoma unspecified.
aFeline leukemia virus (FeLV) antigens: – = negative; + = 1% to ≤25%; ++ = >25% to ≥50%; +++ = >51%.
bB-cell loci: IgH, framework 2 and 3.
cT-cell locus: T-cell receptor gamma.
The quantity and intensity of expression of both proteins varied. Expression was cytoplasmic in all but 1 cat in which cell membrane expression of gp70 (Fig. 6) was observed; cat No. 2 had predominately membranous expressions of gp70.
Of the 8 FeLV serologically negative cats, 5 had FeLV gp70 and/or p27-positive neoplastic cells.
Clonality
Results of clonality testing are listed in Table 2. Clonal rearrangement of IgH was consistent with a B- cell origin in 8 of 17 cats. Despite clonal IgH rearrangement, cat No. 4 lacked expression of CD79a and CD20. In 3 of 17 cats, no amplification of B-cell or T-cell loci was obtained, and 1 of 17 cats had a pseudoclonal IgH rearrangement; immunophenotype of all 4 cats was consistent with B-cell origin of the lesion.
Clonal rearrangement of TCRG along with strong expression of CD3 by tumor cells confirmed T-cell origin in 3 of 17 cats. Polyclonal TCRG arrangement was seen in 1 of 17 cats, although tumor cell morphology and immunophenotype were highly consistent with T-cell lymphoma.
Definitive Diagnosis
Definitive diagnoses based on histology, cytology, immunohistochemistry, and clonality are listed in Table 2. Most cases were diffuse large B-cell lymphoma (11 cases) with a predominance of immunoblastic morphology (Fig. 7) observed in 7 cases. Rare types of diffuse large B-cell lymphoma were plasmacytoid (Fig. 8) and anaplastic cell variants observed in 2 cases each.
Anaplastic large cell lymphomas were diagnosed in 4 cats when a T-cell (3 cases) or natural killer (NK)–like cell (1 case) phenotype was associated with large size, severe pleomorphism with presence of C-shaped nuclei (Hallmark cells), and frequent bi- to multi-nucleated atypical cells (Fig. 9). In cat No. 3, neither immunophenotyping on paraffin-embedded material nor clonality testing assisted in the identification of cell origin, as cells lacked expression of B- and T-cell markers and a pseudoclonal IgH and a polyclonal TCRG rearrangement were observed. Hence, lineage was considered uncertain. However, multiple fresh cytological specimens of the primary cutaneous tumor were available for cat No. 3. Extensive additional immunophenotyping (data not shown) revealed lack of expression of CD21 (B cells), CD3, CD4 and CD8 (T cells), myeloperoxidase, CD1, and the β2-integrin CD11d. Weak expression of CD11b was seen in approximately 20% of cells. Morphology and immunophenotype were consistent with an NK cell origin. Cat No. 12 had an unusual immunophenotype; only 30% of the cytologically atypical cells expressed CD3, and 10% of neoplastic cells were CD20 positive, while CD79 was negative in all cells. In this case, in addition to a clonal IgH rearrangement, pseudoclonality was observed for TCRG. Since CD20 is not considered a lineage-specific B-cell marker, whereas CD3∊-cytoplasmic expression can be found in T cells and in subpopulations of NK cells, immunohistochemistry did not provide a definitive result. In this case, discordance between clonality and marker expression may be explained by antigen loss or lack of primers covering all DNA segments used by the tumor population. Thus, a definitive lineage was not attributed to this case.
Clinical Follow-up
Clinical follow-up was available in 15 of 17 cats and is listed in Supplemental Table S4. Two cats were lost to follow-up. Date to death after diagnosis in 15 of 17 cats ranged from 15 to 761 days. Local recurrence was reported in 9 of 15 cats; 1 cat relapsed twice. Due to recurrence and progression of cutaneous disease, 6 of 16 cats were euthanized, while 9 of 16 died spontaneously; cause of death was not retrieved in 2 cats (cat Nos. 9, 11).
Two cats had cytologically confirmed progression of disease via fine-needle aspirate to the spleen and liver (cat No. 5) and to pleural effusion (cat No. 14). Three cats were submitted for necropsy (cat Nos. 2, 6, 13), and no internal disease was observed.
Discussion
Primary feline cutaneous injection site lymphomas (CLIS) described in this study were extracted from a caseload of 47 cutaneous lymphomas on the basis of their development at sites of previous injection. Relevant traits included lesion development at injection sites, high recurrence rate and microscopic features of necrosis, central cavitation, and inflammation at the tumor periphery. These microscopic features have been consistently described in feline injection site sarcomas (FIS). 11,41,47 A direct connection between injection and the development of a primary cutaneous lymphoma has not been previously confirmed. Madewell et al 44 reported cutaneous lymphoma development following FIS. These lymphomas, which emerged at the same location as FIS following an interval of 3 to 45 months after treatment, were interpreted as the consequence of the mutagenic action of locally applied chemotherapy or radiation therapy. 44 A subset of cats with indolent cutaneous lymphomas, also referred to as cutaneous lymphocytosis, developed lesions on the lateral thorax, shoulder, dorsal neck, flank, and rear legs, all common injection sites in cats. Hence, vaccines were discussed as a possible causation for these lesions. 24 As an additional support to our findings, Meichner and von Bomhard 52 have voiced the suspicion of injection site reactions inducing lymphoma for a subgroup of recently described feline subcutaneous lymphomas. 52
Sites of chronic inflammation, surgery, trauma, metallic implants, and viral infections have been well documented as a potential nidus for B-cell lymphomas in humans. 8 –10,56,57 The WHO classification lists diffuse large B-cell lymphoma (DLBCL) associated with chronic inflammation (DLBCL-ACI) as a specific entity developing in the context of longstanding inflammation frequently in association with EBV. 8,10,56 DLBLC-ACI is often angiocentric 2 and occurs more frequently in middle-aged to old male patients. It develops from terminally differentiated B cells after a long latency period of over 10 years. 8,56 Paralleling DLBCL-ACI, angiocentrism was common in CLIS and was variably associated with angioinvasion, angiodestruction, and tissue necrosis. The latter is considered rare in dermal lymphomas 27,69,71 but has been recently reported as a common feature of feline subcutaneous lymphomas. 52 Likewise, the majority of CLIS of this study were DLBCLs. This is in contrast to previous reports for feline primary cutaneous lymphomas, which are predominantly of T-cell origin with a small cell morphology. 14,21,24,27,71 However, our data parallel recent findings of a predominance of a diffuse large cell morphology and a prevalent B-cell phenotype for feline subcutaneous lymphomas. 52 According to the MI, only 2 DLBCLs were medium grade, while 15 were low grade. This was an unexpected finding, especially since DLBCL, peripheral T-cell lymphoma (PTCL), and anaplastic large cell lymphoma (ALCL) tend to be intermediate to high grade. Grade did not correlate with clinical progression since in most cases where clinical history was available, tumors were aggressive. Thus, for this type of cutaneous lymphoma, grading according to the MI may not be prognostically useful. Unfortunately, due to the low number of cases and the fragmentary clinical information retrieved, a statistical analysis could not be performed, and a higher number of cases should be evaluated to substantiate this hypothesis.
It is noteworthy that primary cutaneous human DLBCLs have a relatively poor prognosis compared with other B-cell cutaneous lymphomas, with a 5-year survival rate of 20% to 55%. 42,65 Similar to our cases, primary human cutaneous DLBCLs manifest as a solitary nodule or as multiple tumors restricted to one anatomic area (regional disease). The most common morphological variants of human DLBCLs are centroblastic, immunoblastic, and anaplastic. 16,65 In this study, a predominance of immunoblastic lymphomas followed by anaplastic and plasmacytoid forms was documented. Immunoblastic morphology has been correlated with a poorer prognosis in humans, 16,35 although this correlation is still considered debatable. 65 Cutaneous immunoblastic lymphomas of this caseload demonstrated highly variable survival times ranging from 15 to 124 days; 2 were lost to follow-up, and 1 was still alive. Thus, a conclusion on their biological behavior could not be clearly drawn. In the human WHO classification, B-cell ALCLs have been excluded from the “true ALCL category” 50 and are now grouped with DLBCLs, 65 while the term ALCL is used for cutaneous or systemic T/NK lymphomas expressing CD30 and differentiated prognostically by anaplastic lymphoma kinase (ALK) expression. 50 Unfortunately, specific markers for ALCL characterization and prognostication are not available in veterinary medicine, and the diagnosis is made only on morphological grounds.
Results from extensive clonality testing in this work emphasize the importance of interpreting data in association with morphologic features as well as immunophenotypic characteristics of the lesions. Assessment of clonal rearrangement of IgH for B cells and TCRG for T cells supported the immunohistochemistry findings in 11 of 17 cats. These included cutaneous B-cell lymphoma in 8 of 17 cats and cutaneous T-cell lymphoma in 3 of 17 cats. However, 1 lesion with T-cell immunophenotype and 1 lesion with B-cell immunophenotype revealed a pseudoclonal and a polyclonal result, respectively. A robust reactive lymphoid population may not allow detection of a neoplastic clone within a polyclonal background, the latter originating from the reactive lymphocytes.
Also, DNA amplification failed in 3 lesions with a B-cell immunophenotype. This may occur if rearrangement is not accounted for by the primers used for the amplification or if somatic hypermutation of primer-binding sequences occurs and impedes the binding of oligonucleotide primers, thus preventing the detection of the rearranged sequences. Impaired sensitivity of B-cell clonality analysis has been attributed to somatic hypermutation in feline intestinal lymphoma. 53 False-negative results have also been attributed to extended time of tissue fixation prior to embedding, as well as inhibition of DNA amplification due to the presence of substances such as formalin, hemoglobin, or residues of the extraction process contaminating the nucleic acid extract. 45 Despite the fact that clonality was not able to support the diagnosis in some cases, morphologic features of the lesions as well as the tumor cell immunophenotype were consistent with a neoplastic lymphoid process of B-cell origin in the 4 cats and T-cell origin in 1 cat.
Initially, the cell of origin remained undetermined in 2 cases, as immunophenotype and clonality were contradictory or inconclusive. The possibility of an NK cell proliferation was considered for cat No. 3. The tumor cells were of leukocytic origin with a morphology consistent with large lymphocytes. However, neoplastic cells lacked expression of B-cell or T-cell markers, and rearrangement of IgH or TCRG was not clonal. Immunocytochemistry on fresh cytological samples further supported an NK-like origin that was finally attributed to this tumor. However, at this point, specific markers for feline NK cells are not available to definitely confirm this hypothesis. In cat No. 12, only a very small subpopulation of tumor cells expressed CD20 (but were CD79 negative), while a slightly larger portion expressed CD3; clonality testing revealed a clonal IgH rearrangement and a pseudoclonal TCRG rearrangement. Based on marker expression and clonality, a clear origin was not attributed since the tumor might have undergone antigen loss, a common event in B- and T-cell lymphomas, or primers might not have covered the gene segments used by tumor cells. However, a T-cell origin was suspected since the case closely resembled previous lymphomas described in humans. 3,46 Indeed, CD20-positive T-cell lymphomas are frequently reported in humans. 3,49,58 These results further heighten that CD20 should not be considered a lineage-specific B-cell marker and should not be used alone to identify neoplastic B cells.
It is important to emphasize that regardless of the variable clonal and phenotypic results, in all these cases, a rapid relapse and the aggressive clinical course confirmed the diagnosis of lymphoma.
As an adjunctive unexpected immunohistochemical finding, at least one of the FeLV antigens investigated in this study was expressed by neoplastic cells in 5 FeLV serologically negative cats. Also, in 5 cases (3 serologically negative and 2 unknown), discordance of FeLV p27 and gp70 staining was documented. Similar results have been previously reported. 33,40,62 FeLV-positive cats developing FeLV enteritis with gp70 strong expression and concurrent p27 negativity have been reported. 40 Interestingly, FeLV-positive nondiseased cats show intense p27 expression in normal intestinal crypts with concurrent negativity to gp70. Moreover, in cats with FeLV focal infection, p27 expression may be variable and discordant, and positive and negative results may alternate. 30 Thus, p27 capsid protein cell positivity indicates that viral infection has occurred at a certain point but does not imply productive and pathogenic viral assembly and, hence, does not indicate infection or disease in progress. 30,66
Kipar et al 40 document a correlation between gp70 intensity and the presence and severity of intestinal lesions, supporting the main pathogenic role of envelope proteins. Thus, FeLV gp70 envelope protein expression denotes viral assembly, confirming viral replication and productive infection. 30,66 Previously, FeLV-positive serology has been reported as low as 10% in feline cutaneous lymphomas. 34 However, a negative FeLV serology does not necessarily contradict the involvement of FeLV in cats with CLIS. Noteworthy, FeLV provirus integration has been demonstrated in 2 FeLV serologically negative cats with primary cutaneous epitheliotropic 67 and nonepitheliotropic lymphomas. 19 During regressive and focal FeLV infection, cats may be seronegative, but FeLV provirus can be demonstrated in peripheral blood and bone marrow cells by PCR. 30,68 Therefore, seronegative cats may bear the virus insert, but transcription and reactivation may occur only upon neoplastic transformation of infected cells. 19
To further complicate the correlation between serology and FeLV-specific protein expression, disorderly and abnormal nonfunctional translation and assembly of FeLV have been described. 30 Moreover, most FeLV tests evaluate for the presence of p27 antibodies or antigens, and FeLV negative serology may derive from the lack of translation of p27, while gp70 may still be produced. Last but not least, differences in glycosylation and abnormal protein synthesis with structural changes have been described, especially for gp70. 40 These changes may alter the monoclonal antibody binding with immunohistochemical false-negative results.
Chronic inflammation following the local delivery of a persistent immunogenic stimulus may have contributed to reactivation of FeLV transcription and development of CLIS in selected cases. This hypothesis is supported by FeLV protein expression by neoplastic cells in cats with CLIS, with some cases expressing both FeLV p27 and gp70. To support this theory, quantitative reverse transcriptase PCR for FeLV will be necessary. Similarly, inflammation has been implicated in the reactivation and proliferation of EBV-transformed B cells and seems to be the most accredited pathogenesis for DLBCL-ACI in humans. 8 Chronic inflammation enables virally transformed B cells to escape from host immune surveillance through production of interleukin (IL)–10 and providing autocrine and paracrine cell growth stimuli via IL-6 production. Furthermore, the finding of angiocentrism may derive from antigen persistence or immune complex deposition in vascular walls secondary to antigen challenge, followed by the expression of specific homing and tethering molecules by endothelial cells in the setting of the vaccine immune stimulation.
Conclusions
In summary, cutaneous lymphomas arising in areas commonly used for injection resembled previously reported cases in cats 52 and that of human DLBLC-ACI 60,61 in which persistent chronic inflammation and immune stimulation in association with a favorable cytokine milieu lead to neoplastic transformation of lesional lymphocytes, facilitated in some instances by viral transformation. In conclusion, CLIS are angiocentric, angioinvasive, primary cutaneous that are predominantly large cell lymphomas with aggressive clinical behavior. They represent a distinct clinicopathological entity among feline cutaneous lymphomas that arise in areas commonly used for injections. Samples from severely necrotizing panniculitis in areas of injections need to be carefully screened in view of an underlying angiocentric lymphoma.
Footnotes
Acknowledgements
We are grateful to Dr V. E. O. Valli for assisting in the diagnosis of DLBLC subtypes.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.