
Abstract
Chordomas are uncommon neoplasms arising from notochord remnants, most commonly occurring in the axial skeleton. Extraskeletal soft tissue chordomas are rare primary tumors, and primary alimentary tract chordomas have not been reported. Herein we report 24 cases of spontaneous primary intestinal chordomas in zebrafish, as well as 9 spontaneous vertebral chordomas. Both intestinal and vertebral tumors showed invasive behavior, although more commonly in the latter. In all cases of primary intestinal chordomas, there was no axial or peripheral skeletal or other nonvisceral involvement. Although uncommon, intestinal chordomas represent a unique background lesion in aged zebrafish.
Chordomas are uncommon primary malignant neoplasms of notochord origin that most frequently affect the axial skeleton in humans. The overall incidence is reported to be 8.4 per 10 million, with a higher incidence in males and individuals older than 40 years. 11,16 Tumors are reported to arise in the cranium (frequently clival, 32%), spine (33%), and sacrum (29%). 11,16 Chordomas are typically lobulated with fibrous trabeculae, and the presence of numerous vacuolated (physaliphorous) cells is characteristic. 12 Although they infrequently metastasize, tumors are typically locally invasive with frequent recurrence, often owing to the difficulty of complete resection. 16 Extra-axial skeletal chordomas are rare in people; extraskeletal soft-tissue chordomas are exceptionally rare, with less than 20 reported cases. 9
Chordomas are uncommon skeletal tumors in most animals. Chordomas are relatively more frequent in ferrets, occurring as axial tumors of the caudal vertebrae and skull base. 3 A number of tumors have also been reported in F344 rats, with a notably high rate of pulmonary metastases (75%). 19 No spontaneous extraskeletal chordomas have been reported in animals. Parachordoma, a variant of myoepithelioma with myxoid differentiation that resembles chordoma, has been reported only once in the veterinary literature, in the gluteal muscle of a dog. However, the distinction between parachordoma and chordoma in that case was largely informed by the extra-axial location rather than morphologic characteristics, and the authors acknowledged uncertainty in the diagnosis. 6 No spontaneous extraskeletal soft-tissue chordomas originating in the gastrointestinal tract have been reported to date in any species. Primary vertebral chordomas have been previously reported as a rare tumor in zebrafish (2% incidence), but no spontaneous chordomas in any location have been reported in any other species of fish. 17 Recently, a transgenic zebrafish model of vertebral chordoma was reported, via expression of the HRAS oncogene in the notochord. 1 Herein we describe spontaneous primary intestinal and vertebral chordomas in 33 laboratory zebrafish, the former representing a novel tumor type not previously described in any species.
Materials and Methods
Thirty-two of the cases came from the archives of the Zebrafish International Resource Center (ZIRC) or from the case files of Dr Jan Spitsbergen from her studies of spontaneous and carcinogen-induced neoplasia. One of the cases was identified prospectively in a mutant screen (Mullins and Fisher [UPenn] and Cheng [PSUHMC]).
Zebrafish were processed for histology by several different methods, depending on the source or study. For some carcinogenesis studies, fish were euthanatized in tricaine methanesulfonate (MS 222; Argent Laboratories, Redmond, WA), pH 7.4, in phosphate buffer; the tail was removed, and an incision was made through the ventral abdominal wall from the heart to the anus to promote internal fixation. Fish were fixed in Bouin’s fixative for 24 hours. Fish were dehydrated in a graded series of ethanol solutions, then embedded in paraffin. Sagittal step sections were cut from the fish’s left side. Three 4- to 6-micron sections were saved and placed onto a single glass slide: 1 section through the lens of the left eye, 1 just medial to the left eye, and 1 from midline. Sections were stained with hematoxylin and eosin (HE). For broodstock and more recent carcinogenesis bioassays, fish were fixed in buffered zinc formalin for 24 hours and decalcified for 48 hours in Cal X II (formic acid/formalin; Fisher Scientific, Waltham, MA), and both halves of the fish were sectioned for histology. Nine step sections were cut between the middle of the lens of the left eye and the middle of the lens of the right eye. Three sections were placed onto each of 3 slides (9 sections total) and stained routinely with HE. For diagnostic cases submitted to the ZIRC at the University of Oregon, fish were routinely fixed in Dietrich’s fixative and decalcified overnight in 5% trichloroacetic acid. Fish were bisected for embedding by cutting sagittally, just to the left of midline, using a razor blade. The 2 halves were placed into a single cassette. Detailed histology protocols are available on the ZIRC website (http://zebrafish.org/zirc/health/diseaseManual.php). Sections from 7 cases were stained with Masson’s trichrome or Alcian blue (pH 2.5) periodic acid–Schiff (PAS) with and without hyaluronidase digestion (H3884; Sigma-Aldrich, St Louis, MO). All images were obtained with an Olympus BX51 microscope and DP71 digital camera using cellSens Standard 1.6 imaging software (Olympus America, Center Valley, PA).
Results
Most fish had no reported history of experimental manipulation, and many tumors were incidental findings in fish submitted as colony sentinels. Demographic characteristics of fish with intestinal tumors are reported in Supplemental Table S1. Average age was 454 days, and there were 10 females and 12 males (sex was not reported for 2 fish). One fish (I22) was only 74 days old. Only a single fish (I11) was exposed to a known carcinogen, diethylnitrosamine. Three fish had genetic manipulations of possible relevance. One fish had morpholino knockdown of the tumor suppressor brca2 (fish I9), but a tumor was also identified in a scrambled morpholino control (fish I10). Morpholinos are modified antisense oligonucleotides targeted to specific messenger RNA (mRNA) transcripts commonly used to silence gene expression in zebrafish (http://www.ncbi.nlm.nih.gov/genome/probe/doc/TechMorpholino.shtml). One fish (fish I17) had an inactivating M214K missense mutation in the DNA-binding domain of the tumor suppressor tp53 (ZFIN ID: ZDB-ALT-050428-2), and lecithin retinol acyltransferase b (lratb, fish I13) is a gene involved in retinol metabolism that is specifically expressed in the notochord (ZDB-GENE-060810-31).
Tumors were well differentiated and resembled normal notochord present in the spine of adult zebrafish (Fig. 1). Intestinal tumors formed sessile polypoid exophytic and expansile masses projecting into the intestinal lumen (Fig. 2). Subgrossly, tumors were distinctly multilobular, separated by fibrous bands, and expansile, consisting of multiple well-circumscribed and encapsulated nodules. Tumors were more commonly located in the cranial half of the intestine. In 4 fish (cases I7, I9, I19, and I24), multiple individual nodules were scattered widely throughout the intestine (Fig. 3). Within the lobules were 1 to several thin layers of plump spindloid cells arranged parallel to the capsule. The centers of the lobules contained numerous individual or clustered large round cells with compressed peripheral ovoid nuclei and a large single clear cytoplasmic vacuole with sharp margins (resembling physaliphorous cells; Fig. 4). Less commonly, these cells had abundant pale eosinophilic cytoplasm and central nuclei. Clusters of small round (glomoid) cells were absent. There were moderate to abundant amounts of glassy eosinophilic acellular matrix between the cells. Mitotic figures were absent. The overlying intestinal epithelium was often attenuated but only rarely ulcerated. Both normal vertebral and neoplastic intestinal matrix was weakly positive with Alcian blue and resistant to hyaluronidase digestion (Fig. 5). In case I10, multiple nodules were present in the exocrine pancreas (Fig. 6). In case I11, multiple nodules were present in the dorsal wall of the cranial intestine as well as within the adjacent liver.
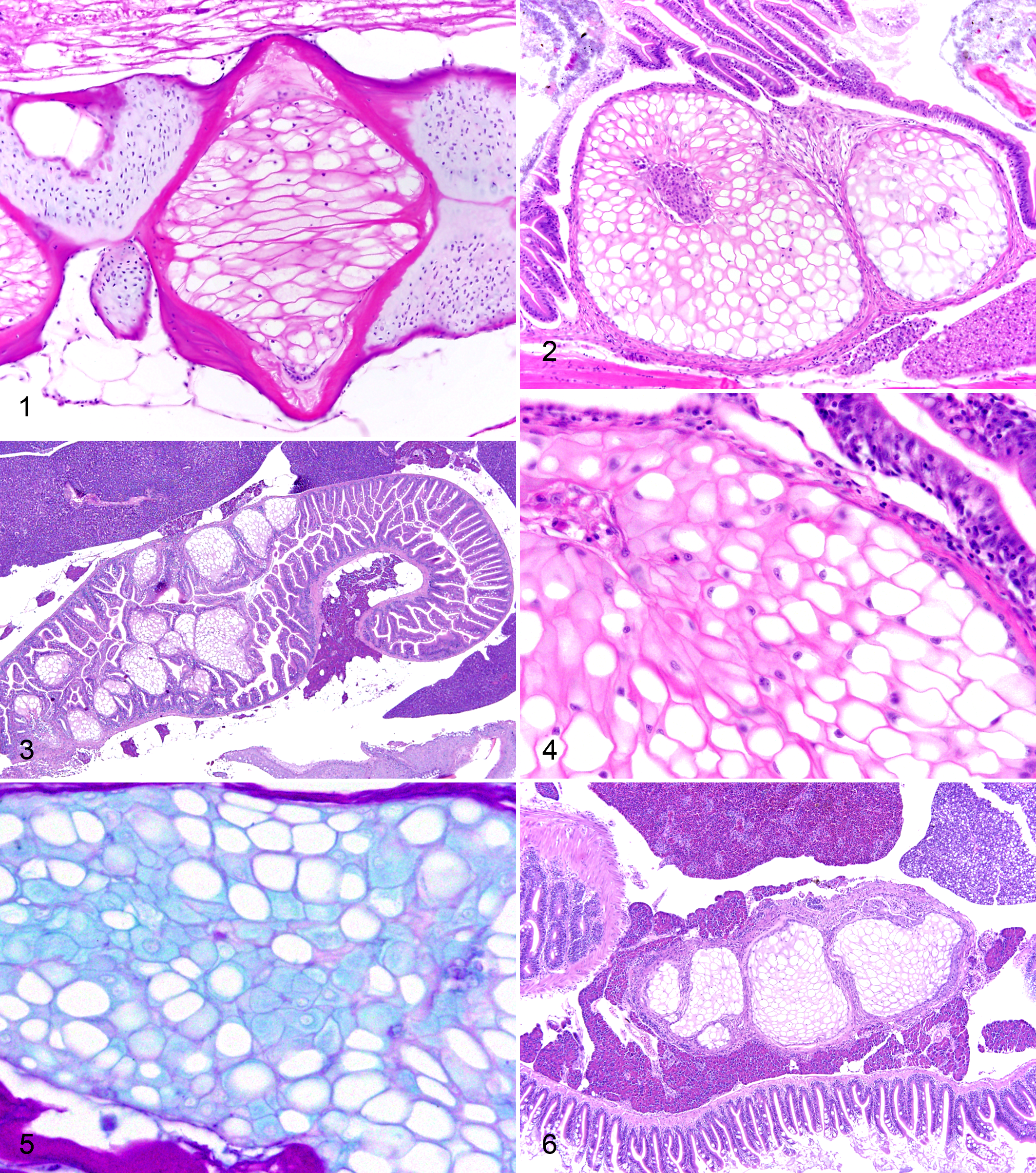
Demographic characteristics of fish with vertebral tumors are reported in Supplemental Table S2. Average age was 482 days, similar to intestinal tumors. There were 3 females and 2 males (sex was not reported for 4 fish). Two fish (V4 and V5) were exposed to known carcinogens, diethylnitrosamine and dibenzo[a, l]pyrene. A skeletal deformity was clinically identified in only 1 fish.
Spinal tumors were morphologically similar to intestinal tumors. Four of the 9 vertebral tumors were invasive into the central nervous system, and 1 additional tumor was invasive into the pancreas. Invasion tended to be protrusive, expansile, and nodular rather than infiltrative or finger-like. Fish with vertebral tumors did not show any evidence of intestinal involvement, nor did those with intestinal tumors show any evidence of vertebral involvement.
In zebrafish that were 1.5 years or older, vertebrae often had multifocal mineralization of the vertebral notochord material, especially in and around the dorsal and ventral protrusions of the “hourglass-shaped” structures containing notochord material (Suppl. Fig. S1). In these areas of mineralization, hyperplasia of notochord cells often occurred, resulting in clusters of small round cells containing scant cytoplasm. Amorphous masses of mineralized material containing scattered notochord cells occasionally ruptured through the notochord sheath in the areas of dorsal and ventral protrusions of notochord in the spine. These degenerative changes were easily differentiated from chordoma because chordoma did not contain the mineralization seen in aging vertebrae.
Discussion
Based on histologic and histochemical characteristics, the tumors of the zebrafish intestine and vertebrae described are appropriately classified as chordomas. The major differential diagnosis for extraskeletal soft-tissue chordoma is parachordoma. Parachordomas are mixed tumors classified as variants of myoepithelioma but have vacuolated cells similar to chordoma. 8 Small round glomoid cells are present in parachordoma but absent in chordoma. 4,5 Parachordomas have a myxoid matrix that is Alcian blue positive but hyaluronidase sensitive. 5 Chordomas, in contrast, have an Alcian blue–positive, hyaluronidase-resistant matrix. Although some have used the axial vs extra-axial location to distinguish between chordoma and parachordoma, respectively, this is not an appropriate diagnostic criterion. 9 Extraskeletal soft-tissue chordoma does exist, is extremely rare, and is histologically, immunohistochemically, and behaviorally distinct from parachordoma, the former having metastatic potential that is absent in the latter. 9,15 The distinction between chordoma and parachordoma also takes on clinical significance in humans, as the former is invasive and likely to recur following surgical excision. It should be noted, however, that postresection local recurrence of parachordomas has been reported. 15 Gastrointestinal involvement by parachordoma is uncommon in humans, with only a single report of a primary tumor in the gastric serosa. 18
Expression of brachyury, a nuclear T-box transcription factor (T, brachyury homolog, HGNC:11515), is necessary and sufficient to distinguish chordoma from parachordoma in humans, as the tumors share a number of other markers, including cytokeratin (CK) 8/18, CK 7, S100, and epithelial membrane antigen. 7,9,15,21 Chordomas are also variably positive for CK20, CK 1/5/10/14, and carcinoembryonic antigen. 15 Brachyury is a regulator of embryonic notochord formation, and chordomas are positive for brachyury expression, whereas parachordomas are negative. 13 Attempts to detect brachyury in formalin-fixed, paraffin-embedded zebrafish were unsuccessful. Further complicating the matter, zebrafish, like many teleost fish, have 2 functional brachyury orthologues, bra and ntla. 10 Although ultrastructural studies could assist in the differentiation of chordoma and parachordoma, the nature and availability of specimens in this series exclude this possibility.
Another significant differential for vertebral chordoma is hyperplasia secondary to degeneration of the notochord, a common age-related finding in laboratory zebrafish. Although both processes include proliferation of notochord cells, notochord degeneration is characterized by prominent mineralization of the notochord material. Alternatively, while some would perhaps classify the lesions described in these fish as nonneoplastic ectopic notochord rests (choristomas), the size, multiplicity, and invasive behavior demonstrated by these nodules are consistent with a neoplasm.
Although generally considered uncommon tumors, chordomas do occur as spontaneous background lesions in zebrafish. Primary intestinal chordoma, a condition not previously reported in any species, appears to be more common in zebrafish than chordoma of the axial skeleton. Overall, the tumor frequency is low. Chordomas were identified in only 25 of the 11 902 zebrafish examined between 2003 and the end of 2013. Over 20 000 fish were evaluated for carcinogenesis studies, but fewer than 75 fish have been evaluated to date in the UPenn study. Lesions disproportionately affected older fish (average age >400 days), consistent with observations in humans with chordoma. Both sexes were approximately equally represented in this data set. There were no significant commonalities in the experimental or clinical histories, with most fish presenting as unmanipulated wild types. Fish with mutations in connexin 41.8 and longfin (cx41.8t1; lof t2 ) appear to be overrepresented in our data set, with 2 spinal chordomas and 1 intestinal chordoma. The reasons for this are not clear but may be due to sampling bias related to the lines of fish submitted for evaluation. Interestingly, 1 case (fish I3) had a null allele for the ntla orthologue, although chordoma has not been described as a phenotype in those mutants. 14 However, genetic mutations affecting the brachyury T gene in human chordomas are characterized by amplification events, rather than silencing or loss of function. 22 Silencing of brachyury expression by small hairpin RNA has inhibited chordoma growth in vitro, consistent with the proposed central role of this transcription factor in driving tumorigenesis. 13
Because intra-abdominal metastases of chordomas have been reported in humans, 9,20 we felt it important to determine whether there were primary vertebral chordomas in the cases with intestinal tumors. In the fish examined, no involvement of vertebrae, other bones, or soft tissue was observed, with the exception of local invasion into adjacent viscera. The reason for the occurrence of primary intestinal chordomas is potentially explained by Burger et al 1 in their transgenic zebrafish model. In that model, the sonic hedgehog b promoter (shhb [ZDB-GENE-980526-41], previously tiggy winkle hedgehog [twhh]) was used to drive HRAS expression in the intestine. Because shhb expression also occurs in the notochord, abnormalities in that organ were also present. It is interesting to speculate that spontaneous shhb mutations may drive tumorigenesis in both sites, as alteration of the sonic hedgehog signaling pathway in human chordoma formation has been previously suggested. 2
Herein we describe the demographic and morphologic characteristics of primary intestinal and vertebral chordomas in laboratory zebrafish. Uniquely, intestinal chordomas exist as a distinct clinical entity in this species.
Footnotes
Acknowledgements
We thank Amy Kugath and Drs Shannon Fisher, Mary Mullins, and Keith Cheng for contributing case I13, identified in a histologic phenotypic screen (NIH R01HD069321-01). Qing Zhong provided immunohistochemistry expertise and Ellen Mullady performed histochemistry. Dr Elizabeth E. Frauenhoffer provided invaluable expert consultation.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: Parts of this work were funded by the US Public Health Service, National Institutes of Environmental Health Sciences (grants R01ES011587, R21ES013124, P30ESO3850, and P30ES00210); the National Center for Research Resources (grant 3P40RR12546 and its supplement 03S1); and the US Army (contract DAMD 17-91Z1043). The Zebrafish International Resource Center is supported by the National Institutes of Health Office of Research Infrastructure Programs.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.