
Abstract
Feline infectious peritonitis (FIP) is one of the most important fatal infectious diseases of cats, the pathogenesis of which has not yet been fully revealed. The present review focuses on the biology of feline coronavirus (FCoV) infection and the pathogenesis and pathological features of FIP. Recent studies have revealed functions of many viral proteins, differing receptor specificity for type I and type II FCoV, and genomic differences between feline enteric coronaviruses (FECVs) and FIP viruses (FIPVs). FECV and FIP also exhibit functional differences, since FECVs replicate mainly in intestinal epithelium and are shed in feces, and FIPVs replicate efficiently in monocytes and induce systemic disease. Thus, key events in the pathogenesis of FIP are systemic infection with FIPV, effective and sustainable viral replication in monocytes, and activation of infected monocytes. The host’s genetics and immune system also play important roles. It is the activation of monocytes and macrophages that directly leads to the pathologic features of FIP, including vasculitis, body cavity effusions, and fibrinous and granulomatous inflammatory lesions. Advances have been made in the clinical diagnosis of FIP, based on the clinical pathologic findings, serologic testing, and detection of virus using molecular (polymerase chain reaction) or antibody-based methods. Nevertheless, the clinical diagnosis remains challenging in particular in the dry form of FIP, which is partly due to the incomplete understanding of infection biology and pathogenesis in FIP. So, while much progress has been made, many aspects of FIP pathogenesis still remain an enigma.
Keywords
History
Feline infectious peritonitis (FIP) was first thoroughly described and named in 1966, when experimental infections of healthy cats with organ material of diseased animals confirmed it as a specific, fatal infectious disease, and a viral etiology was suspected. 178 However, the disease syndrome had already been observed in the 1950s and 1960s in the United States, and even earlier, a very similar disease had been reported from cats in Naples. 17,86
In 1968, the viral etiology was demonstrated. 171 The virus morphology suggested a coronavirus (CoV), which was finally confirmed in 1976. 121,126,173 The virus was first grown in peritoneal cells of experimentally infected cats and, after propagation in cell culture, was shown to cause FIP in 100% of intraperitoneally infected animals. 128 Subsequently, the macrophage cell line Felis catus whole fetus-4 (Fcwf-4) has predominantly been used for virus propagation. 90,128
In his recent review article, based on more than 40 years of work in the field, Pedersen 136 speculated on the reason for the likely emergence of FIP in the 20th century. He considered as the most relevant potential factors the evolution of the feline CoV (FCoV) alongside CoV of pigs and dogs, the development of virulent FIP virus mutations from enteric FCoV that had only evolved at that stage, and the changes in keeping and especially breeding of cats due to their increasing popularity as pets.
At present, despite decades of research on its etiology, pathogenesis, transmission, and prevention, FIP is still one of the most frequent fatal and infectious feline disease for which there is so far no effective cure.
Feline Coronaviruses
Virology
FCoVs are pleomorphic, enveloped, single-stranded positive-sense RNA viruses with an almost 30-kb nonsegmented genome and 11 putative open reading frames (ORFs). They belong to the family Coronaviridae, order Nidovirales, and, together with canine coronavirus (CCoV) and transmissible gastroenteritis virus (TEGV) of pigs, belong to the subfamily Coronavirinae, genus Alphacoronavirus, species Alphacoronavirus 1 (Fig. 1). 63
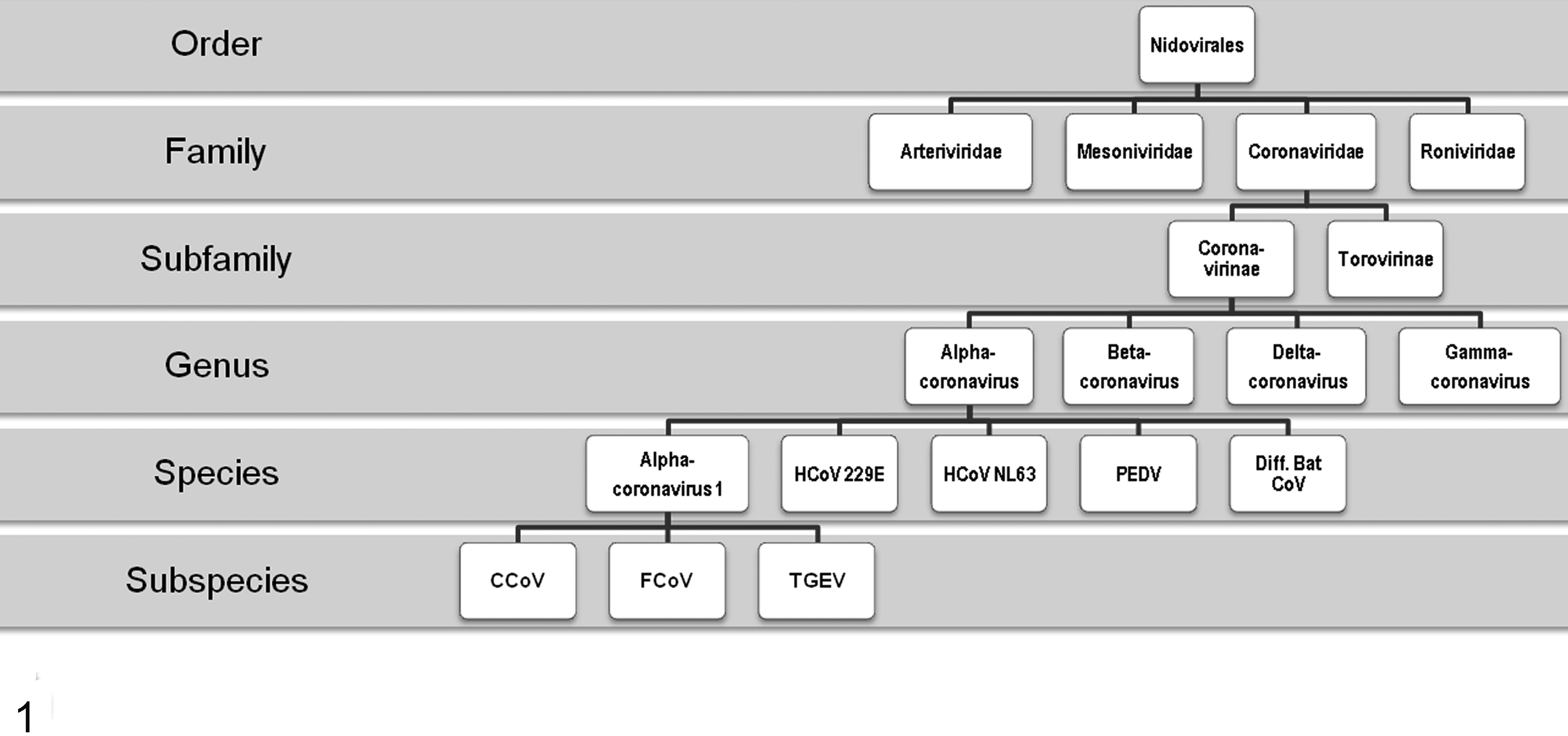
Feline coronavirus classification flowchart. CCoV, canine coronavirus; FCoV, feline coronavirus; HCoV, human coronavirus; PDEV, porcine epidemic diarrhea virus; TGEV, transmissible gastroenteritis virus.
At the 5′ end of the FCoV genome, approximately 20 kb comprise the 2 overlapping ORF 1a and 1b that encode for 2 polypeptides that are subsequently enzymatically cleaved into 16 nonstructural functional proteins mainly involved in the synthesis of the viral RNA (viral replicase). The remaining genome contains 9 ORFs that encode for 4 structural proteins (spike [S], nucleocapsid [N], membrane [M], and envelope [E]) and 5 group-specific, accessory proteins (3a–c, 7a, and b) (Fig. 2a). These are expressed individually from a nested set of subgenomic messenger RNAs (mRNAs) that each contains a leader RNA sequence derived from the 5′ end of the genome and is generated by discontinuous transcription from the 3′ end of the genome. 45,47
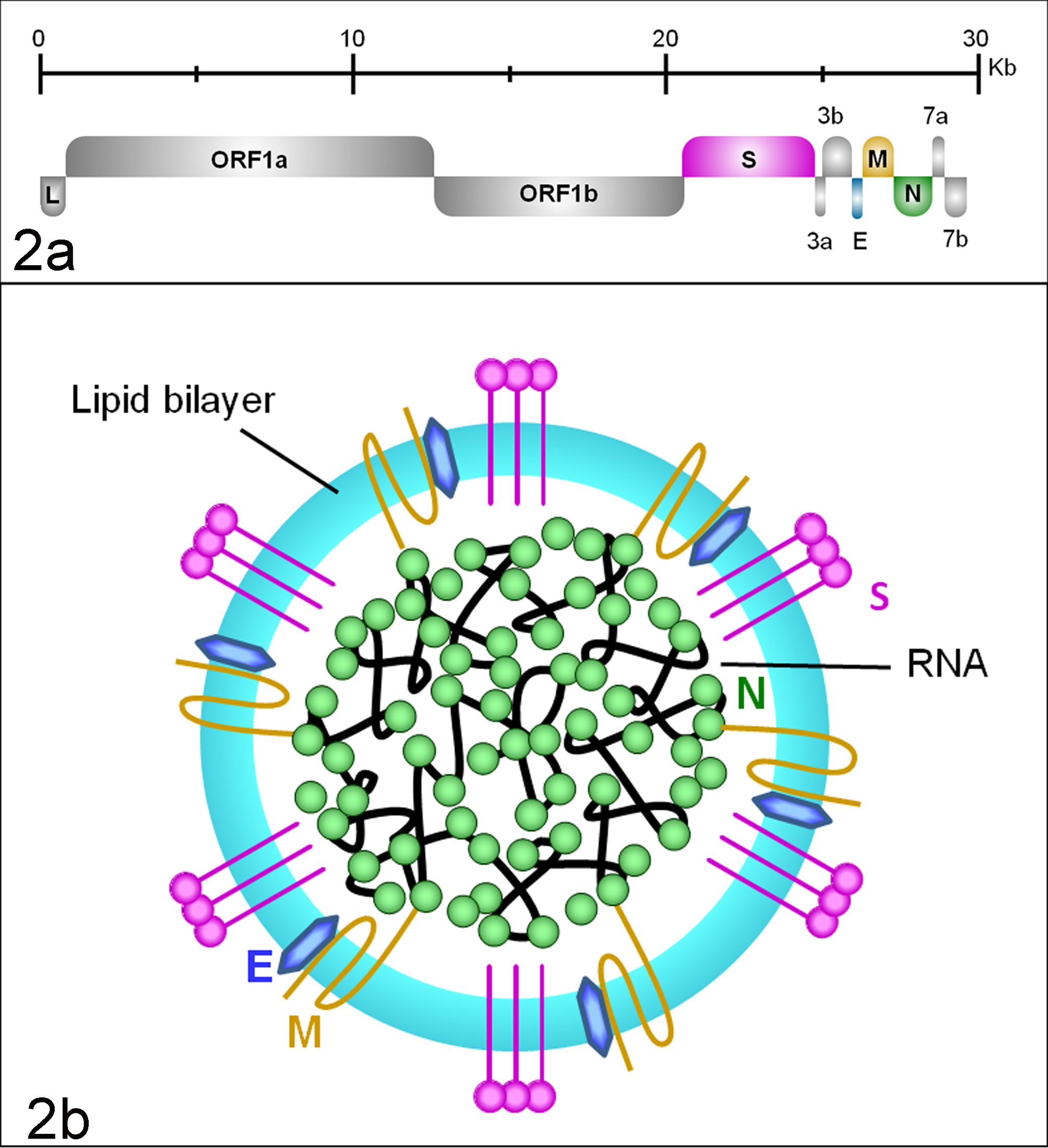
Feline coronavirus (FCoV) genome and structure. (a) Genomic organization of FCoV. E, envelope; Kb, kilobases; L, leader sequence (LTR); M, membrane; N, nucleocapsid; ORF, open reading frame; S, spike. (b) Schematic drawing of the viral structure. E, envelope protein; M, membrane protein; N, nucleocapsid; S, spike protein (peplomer).
The CoV envelope is formed by the S protein, a 180- to 200-kDa glycoprotein arranged in peplomers that is important for the induction of an antibody response and cell-mediated immunity in the host. The S peplomers are 12 to 24 nm long, dome shaped, and arranged like a crown (Fig. 2b); they are the key determinants of cell tropism. 13 The S protein is a type I transmembrane protein with a very short C-terminal cytoplasmic tail and a long N-terminal ectodomain that is divided into a N-terminal (S1) domain responsible for receptor binding and a C-terminal (S2) domain containing the fusion peptide, which mediates fusion with the target cell membrane. 18
The M and E proteins are smaller surface glycoproteins and important for virus maturation, assembly, budding, and interaction with the host cell. The M protein, with a mass of approximately 29 kDa, penetrates the envelope, connects it to the capsid, and participates in the RNA packaging. E proteins are type III membrane proteins of about 9 kDa that interact with the M protein in the budding compartment of the host cell. 45 In mouse hepatitis virus (MHV), they can induce apoptosis. 7 The N proteins have a molecular weight of approximately 50 kDa. Together with the viral RNA, they form the flexible, helical nucleocapsid and seem to be critical for viral transcription. 118 Vaccine studies based on the N protein indicate that it induces cell-mediated immunity and can play a protective role. 85
So far, no definite specific function could be ascribed to the accessory proteins. The 71 to 72 amino acid 3a and b proteins are well conserved among subspecies 1 alphacoronaviruses. Since they lack predicted hydrophobic segments, both are thought to be located and exert their function in the cytoplasm. ORF 3c is very well conserved among the alphacoronavirus genus, and its predicted sequence indicates that it is a class III triple-spanning membrane protein of 238 to 244 residues, with a topology similar to that of the M protein. 70,120 It has recently been shown that an intact 3c gene is necessary for the replication of enteric FCoV (Table 1). 26 ORF 7a encodes for a small membrane protein of approximately 10 kDa with an N-terminal cleavable signal sequence and a C-terminal transmembrane domain. It has just been shown that the FCoV 7a protein is a type I interferon (IFN) antagonist and protects the virus from the antiviral state induced by IFN in a ORF 3–dependent manner. 35 The ORF 7b is present only in FCoV, CCoV, and ferret CoV and encodes for a soluble glycoprotein of 207 residues (approximately 24 kDa) that has been shown to induce antibodies in naturally infected cats. 79,93 Deletions occur spontaneously in cell culture but appear not to have an in vivo effect. 79
Results of Recent In Vitro and In Vivo Research Into the Presence and Effect of Changes in Viral Proteins in Association With Viral Replication and Virulence.

BMDM, bone marrow–derived macrophages; FCoV, feline coronavirus; FIP, feline infectious peritonitis.
CoVs occur in many mammalian species, including humans, and in birds. They lead to acute or chronic infections and, depending on their cell tropism, induce highly variable diseases in their hosts. The host and tissue specificity is dependent on sequence variations of the S gene as well as receptor usage and distribution. 47
RNA viruses have high error rates in their replication and therefore occur as quasispecies (ie, groups of related genotypes). 37 With every RNA replication of CoV, several point mutations occur. Genetic diversity within a quasispecies has been suggested to contribute to pathogenesis by cooperative interactions among variant viruses within a population. 169 On the other hand, proofreading or repair mechanisms, mediated by the exoribonuclease in the replicase complex, allow RNA viruses to evolve while keeping a balance between adaptation and viral fitness. 37 Also, homologous RNA recombination during mixed infections of closely related CoV strains in the same group promotes cross-species transmission and pathogenesis; the cat might represent a “mixing vessel” as in vitro studies showed that feline aminopeptidase N can be used as a functional receptor by closely related alphacoronaviruses, such as FCoV, CCoV, TGEV, and human coronavirus HCV-229E. 163
Genome sequences and subsequent phylogenetic analysis showed that FCoV isolates form geographical clusters. 10 –12,42,105,137 FCoVs from cats of the same household exhibit more than 95% genetic identity, suggesting infection from a common virus. 5,105,167 Focusing on the S gene, 1 study specifically examined the evolution of virus strains in cohorts of naturally infected cats over several years. It demonstrated very high conservation of the virus in persistently infected and (recurrently) shedding animals but also showed that cats can become transiently infected and subsequently reinfected with the same or a different strain. There was also evidence of super- or coinfection of persistently infected cats with other strains. 5
Serotypes
As shown by virus-neutralizing antibody reaction and amino acid sequences of the S protein, FCoVs form 2 antigenically distinct serotypes: type I FCoVs, which are difficult to grow in cell culture, and type II FCoVs, which are the consequence of a double recombination between type I FCoV and CCoV. 82,116,132,148 In vitro, the growth kinetics of both serotypes appear to be solely related to the S protein, as determined using a recombinant type I FCoV encoding a type II S protein. 162 For type II FCoV, as for several other alphacoronaviruses, the cell receptor is aminopeptidase N (APN, CD13), which, upon binding to the S protein, mediates the internalization of virus into the target cells. 84,162 Antibody blockage of APN has been shown to severely reduce the infection of bone marrow–derived macrophages (BMDMs) with the type II FIP strain 79–1146. 145 However, it has so far not been confirmed that APN is also the receptor for FCoV II in infected animals.
Furthermore, the receptor for serotype I FCoV is not known. Interestingly, however, isolated feline monocytes rapidly internalize both serotype I and II FIP strains and accumulate the virus particles in endosomes, followed by particle disassembly. 165 Both serotypes can use “dendritic cell (DC)–specific intercellular adhesion molecule (ICAM) grabbing nonintegrin” (DC-SIGN, CD209), a C-type lectin, which recognizes high-mannose oligosaccharides as ligands, to infect monocyte-derived dendritic cells. 143 Co-localization and binding inhibition studies confirmed that DC-SIGN and not APN is involved in the entry process of serotype I FCoV in monocytes, whereas for serotype II FCoV, both APN and DC-SIGN play a role in the infection of monocytes. Specifically, for serotype II, binding is mediated by APN, but DC-SIGN is important for either internalization or a subsequent step. 166 In both models, a role of an unknown co-receptor cannot be excluded.
Both FCoV serotypes can cause FIP, but serological and, more recently, molecular studies confirmed that type I FCoVs dominate by far in the cat population worldwide, with the prevalence of seropositive animals reaching 98%. 5,83,108,111,132,148 Type I FCoVs were shown to induce higher antibody titers than type II FCoVs and were more frequently associated with clinical signs and/or FIP. 108 Nevertheless, other studies report a higher type II prevalence, ranging from 10% to more than 30% in cats with FIP (the latter in an older study in Japan), and sometimes in mixed infection with type I FCoVs. 15,42,111,148
Feline Enteric Coronavirus vs Feline Infectious Peritonitis Virus
FCoVs occur as 2 pathotypes: feline enteric coronavirus (FECV), defined as the “ubiquitous enteric biotype,” and feline infectious peritonitis virus (FIPV), the “virulent biotype that causes FIP in individual cats.” 136 FECV and FIPV cannot be distinguished serologically or morphologically, and for many years, the search for markers that could discern the 2 pathotypes remained unsuccessful.
In the past, it was assumed that the main difference between FECV and FIPV was that FECVs exclusively infect the intestinal epithelium and do not pass the intestinal mucosal barrier, while FIPVs infect and replicate in monocytes/macrophages and can therefore gain access to the blood and induce the disease. 129,130,133 When more sensitive molecular methods became available, this hypothesis was proven too simple. It was shown that FECV can also infect monocytes and that FCoVs generally spread from the initial site of infection, the intestine, via monocyte-associated viremia. 64,97,101,113 Indeed, approximately 80% of cats, healthy or with FIP, in households with endemic FCoV infection were shown to harbor FCoV RNA in their blood monocytes, and healthy cats remained viremic over the 12-month test period. 64 Furthermore, it was shown recently that intraperitoneal inoculation with FECV can, albeit only occasionally, lead to virus shedding within the feces, which confirms FECVs can also spread from extraintestinal sites, again most likely via monocyte-associated viremia. 138
To test whether the difference between FECV and FIPV is based on the exclusive capability of FIPV to replicate in feline monocytes, a new method was developed that demonstrates replicating virus in the blood, through the specific detection of viral M protein mRNA. 150 The protocol was applied in 2 studies on naturally infected cats and showed that FCoV can replicate within monocytes in healthy cats. However, while the first study found a strong correlation between virus replication in the blood and presence of FIP, the second did not confirm this finding. 24,150 Subsequently, a quantitative method detected high levels of viral replication in cats with FIP. 87 Furthermore, a recent experimental study showed that after oronasal infection with known FECV isolates, only very few cats develop viremia, and without evidence of viral replication. 172 Thus, FIPVs seem to be able to replicate to higher numbers in monocytes than FECVs.
Initial molecular studies characterizing FECV and FIPV isolates from the same environment identified deletions in the 3c, 7a, and 7b genes of FIPV and therefore indicated that FECVs are the ancestors of FIPV. 92,167 However, deletion of the 3a–c and/or 7a/7b genes of wild-type FIPV II (79–1146) has been shown to lead to loss of virulence in experimental infections. 69 This initial work was followed by a more rigorous search for virulence markers. The sequencing of structural (S, E, M, N) and accessory (3a–c, 7a, and b) genes of FCoV from feces and diseased tissues of cats with FIP identified significant mutations only in the 3c gene; these resulted in variable truncation of the 3c protein. Virus with the mutated 3c gene was identified in diseased tissues, whereas the FCoV in the feces generally exhibited an intact 3c gene and only in some cases also the mutated form. 137 Together with the results of a previous study, this indicated a role of 3c gene deletions in the viral switch from FECV to FIPV. 137,167 However, the fact that 3c gene deletions were not consistently observed with FIP suggested that additional factors are essential for the acquisition of the FIPV pathotype. 167
Two larger subsequent studies compared the 3c gene of FCoV from healthy cats (FECV) and cats with FIP (FIPV) to further assess its role as a virulence marker. Almost all FECVs carried an intact 3c gene. 28,138 After oronasal inoculation with such FECV isolates, cats became infected and shed the virus with the feces. 138 In contrast, the majority (71%) of FIPVs exhibited 3c mutations (ie, deletions that were sometimes associated with severe truncation and loss of function). Interestingly, FCoVs identified in the feces of cats with FIP generally exhibited an intact 3c gene, which was interpreted as an indication of FECV superinfection. 26 Nonetheless, similar to a previous study, both these studies identified FIPV with an intact 3c gene in diseased tissues in a substantial proportion of cats with FIP (29% and 40%, respectively). 26,138,167 Furthermore, the FCoVs with an intact 3c gene were shown to be indeed FIPV, since they induced FIP after both oronasal and intraperitoneal inoculation. 138 These findings lead to the following conclusions: a mutated/deleted 3c gene alone is not the hallmark of FIPVs, and FCoVs need to carry an intact 3c gene to be able to sustainably replicate in the intestinal epithelium and be infective to other cats.
A nontargeted approach has recently been taken in an attempt to identify further mutations that might be responsible for the change in virulence. Each 11 randomly selected FECVs (from healthy cats) and FIPVs (from cats with FIP confirmed by postmortem examination) were compared, based on full genome sequencing. Differences were found scattered along the entire genome, but a larger genetic variation with 2 hot spots was identified in the S gene. Subsequent sequencing and phylogenetic analysis of more isolates identified the 2 alternative codons in the S gene in more than 95% of the examined FIP cases. Both mutations occurred in the supposed fusion peptide of the S protein, but without any evidence of potential functional consequences. 28 However, due to the mutation rate of RNA viruses and the relative rarity of FIP, the authors concluded that the identified S gene mutations are unlikely solely responsible for the FECV-FIPV virulence switch. 28 More recently, a targeted approach investigated a furin cleavage site in the region between receptor-binding (S1) and fusion (S2) domains of the spike gene. All FECVs showed a well-conserved and functional cleavage motif, whereas FIPVs exhibited several substitutions in critical residues, mostly abrogating the furin cleavage. 110 The authors hypothesized that these mutations could be responsible for the tropism switch of FIPV by allowing the site to be cleaved by alternate proteases (ie, monocyte/macrophage specific proteases, such as cathepsin B and/or matrix metalloprotease-9). 110
A group that has worked extensively on antibody-mediated enhancement (ADE) in FIP used FCoVs that are resistant to virus-neutralizing monoclonal antibodies to search for virulence markers. Their so-called mar-mutant viruses all exhibited mutations in several amino acids in the S1 region. 94 However, when orally administered to cats, only some were found to induce FIP. None of the latter carried mutations in ORF 2 to 7 other than those observed in the S1 region, whereas the avirulent viruses also showed deletions in the 7b gene. 159
A further molecular study on natural cases identified a relatively high diversity of the N protein in endemically infected cat groups, but without any pattern or relation to virulence. 12 Furthermore, analysis of nucleotide substitutions identified residues in the N protein that were subjected to positive selection. These could represent antigenic immunodominant sites, indicating the antigenic role of the N protein in stimulating cell-mediated immunity. 12
In vitro studies complement the in vivo approaches and provide strong evidence that FCoV virulence requires the ability to productively and sustainably infect feline monocytes. This was first indicated in an older study that demonstrated less effective and shorter replication of avirulent FCoV than FIPV in feline peritoneal macrophages and was more recently confirmed in isolated feline monocytes, in which an FIPV (79–1146) established sustainable replication, whereas an FECV (79–1683) could replicate but not sustainably. 38,154 It needs to be emphasized, though, that despite rapid virus binding, internalization, and disassembly, even the replication of FIPV is limited to a very small proportion of macrophages and monocytes. 38,154,165 This strongly suggests that most monocytes/macrophages are resistant to the virus at the time of infection, most likely due to inhibition of genome release and/or translation. 165
Attempts have been made to relate the monocyte/macrophage tropism to differences in the viral protein structure. In BMDM cultures, FECV (79–1683) was shown to infect fewer cells than FIPV (79–1146) and appeared unable to spread the infection. 145 This was determined by the S protein alone and, interestingly, by the membrane-proximal S2 domain involved in virus-mediated membrane fusion and not the receptor binding S1 region. 145 In agreement with these findings, a recent study found that deletions in the S1 gene region did not affect the viral capacity to productively infect feline monocytes. However, an N-terminal 29–amino acid deletion in the 7b gene led to a decrease in virulence, although one of these mutants still retained the capacity to productively infect macrophages, suggesting that this ability is not mediated by ORF 7b. 159
Full sequencing of the FIPV type II strain DF2, the cell culture adapted FCoV strain used in the only commercially available vaccine, has shown that it carries a 338-nt deletion in the ORF 3abc, resulting in the truncation of 3a and 3c and the complete loss of ORF 3b. This virus replicated efficiently in isolated feline monocytes. When the DF2 ORF 3abc was replaced with a genetically closely related, intact CCoV ORF 3abc region, the recombinant virus was able to replicate in feline monocytes but yielded significantly lower virus titers. 9 These results are in contrast to those of an investigation into the relevance of the ORF 3 and 7 proteins for the replication of FCoV in monocytes published a year later, using the type II FIPV 79–1146, which is thought to obtain its virulence through its S protein structure but has also been shown to have a truncated 3c. 34,145 The use of genetically modified viruses with deletion of ORF 3abc (FIPV-▵3), 7ab (FIPV-▵7), or both (FIPV-▵3▵7) demonstrated a lower but sustainable replication capacity in the absence of 3abc, whereas viruses lacking 7ab could undergo only 1 replication cycle. 34,69 Since ORF 7 are located at the 3′ end of the genome where transcription begins, 7a and 7b are produced very early in viral replication; it was therefore concluded that they might neutralize the innate immune response to the virus during the early phase of infection, for example, by counteracting the IFN-mediated induction of an antiviral state, resulting in inhibition of viral replication. 34,45 A contradiction remains to be clarified since Rottier and coworkers 145 showed in their study that mutants lacking the 7b gene could still productively replicate in macrophages and mentioned, without showing the results, that the same applied for mutated viruses lacking both 7a and 7b. This difference might be related to the use of different cells in both experiments, since Rottier et al infected BMDMs, whereas the other study used peripheral blood–derived monocytes. 34,145
In summary, while promising, the above results from several studies do not yet provide a conclusive picture (Table 1). This is likely also due to the general diversity of the study material, particularly with regard to the virus isolates, but also the methodological approaches that have been taken. 136
Prevalence
FIP, together with panleukopenia and viral infections of the upper respiratory tract, is one of the leading infectious causes of death in cats. 136 However, despite the generally high prevalence of FCoV infection in the cat population, which can exceed 90% in multicat environments, FIP morbidity is low and rarely surpasses 5% of infected cats. 41,136 In larger cat groups, the proportion of chronic shedders and the overall frequency of virus shedding represent risk factors. 51 FIP is predominantly a disease of young (6 months to 2 years), purebred, male intact cats. 144 Purebred cats appear to be more susceptible also to FCoV infection in general, since they were overrepresented when healthy mixed populations were screened. 49,104 A recent study indicates that the breed predilection is restricted. While Abyssinians, Bengals, Birmans, Himalayans, Ragdolls, and Rexes were found to have a significantly higher risk for the development of FIP, Burmese, Exotic Shorthairs, Manxes, Persians, Russian Blues, and Siamese cats did not exhibit an increased risk. 139
Transmission, Shedding, and Persistence of FCoV
FCoVs are transmitted via the fecal-oral route and primarily infect enterocytes. 134 Cats can become persistently infected and continuously or intermittently shed virus with the feces. They generally remain healthy despite systemic infection, indicating that healthy (FECV) carriers play a key role in the epidemiology of FIP. 3 –5,64,80,97,103,113 In addition, cats can become repeatedly infected with the same or a different virus strain. These animals will also shed the virus intermittently and can do so recurrently for months. 4,50,71,81,113 Furthermore, there is evidence of a correlation between shedding frequency and intensity and high antibody titers. 73
Experimental studies with type I FECV isolates have demonstrated consistent shedding as early as 2 days and for up to 2 weeks postinfection (PI), with a subsequent decline in fecal viral loads and intermittent shedding up to 20 weeks after this period. 103,113,172 They confirmed previous studies in which oral administration of cell culture–adapted FIPV (Wellcome strain) led to viral antigen expression in the small and large intestine between days 1 and 7 PI, as well as restriction to the cecum and colon on day 14. 153 After clearance from the small intestine, FCoV can apparently spread from the persistently infected colon at later stages, leading to renewed shedding. 103
In natural FIP cases, shedding can occur until death. 3,152 However, compared with diarrhoeic or healthy shedders, the amount of replicating virus shed by cats with FIP is only very low, and replication is significantly lower in the intestines than in organs. 87,137
Despite the generally strong evidence that only FECV and not FIPV is transmitted between cats, a recent study confirmed that the FCoV with a truncated 3c gene found in diseased tissues is occasionally also present in the feces of cats with FIP. 137 While this would suggest that horizontal transmission is possible, a subsequent study indicated that oronasal uptake of shed, fecal FIPV does not lead to FIP. 138 It remains to be clarified whether this is a universal characteristic of FIPV. This could be of relevance for epizootic FIP outbreaks. Outbreaks are defined by the occurrence of FIP in more than 10% of cats in high-prevalence establishments, but the percentage can be lower in an environment with generally very low FIP prevalence. 41,136,141
In this context, a closer look into an “artificially induced” FIP outbreak that we monitored a number of years ago is of interest. As part of a trial to investigate the efficacy of an FIP vaccine, a multicat environment similar to an animal shelter was created. This housed 40 specific pathogen-free (SPF) cats (20 female, 20 male neutered) to which 10 clinically healthy cats (aged 6 months to 3 years) from different animal shelters were introduced. The latter had been selected since they tested positive for circulating FCoV immune complexes. 97,99,101 Within 1 week after introduction of the shelter cats and during which fighting for the establishment of hierarchies occurred, several animals developed transient signs of upper respiratory tract disease. These subsided, but the first FIP cases occurred in week 6 (n = 4), followed by a peak in weeks 7, 8 (each n = 4), and 9 (n = 3), with further individual cases in weeks 14 to 22. A total of 23 animals (45%; 22/40 [55%] SPF cats, 1 shelter cat) succumbed to the disease. Most SPF cats that died with FIP (18/22; 82%) had shown previous or concurrent signs of upper respiratory tract disease. 109 Only 14 of 40 (35%) SPF cats survived the challenge period of 21 weeks. They had all become infected, had shed virus at least intermittently, and exhibited histological features in the hemolymphatic tissues that indicate a strong immune response to the virus. 97,99,102
The characteristics of this experimental FIP epidemic, using natural infections with field viruses, support assumptions that the occurrence of outbreaks is associated with factors related to the environment (such as crowding, concurrent infections, and long-term exposure to shedders), the virus (such as virulence, replication rate, and mutation rate of the strain), and the host (individual differences in the immune response to FCoV). 41 It also appears likely that horizontal FIPV transmission played a role in this particular case. It might indeed be the generally low amount—if any—of FIPV that is shed with the feces, as well as the inability of at least some FIPVs shed with the feces to induce FIP, that prevents more frequent horizontal transmission of the disease. 137,138
The main site of FCoV persistence is the colon, where viral antigen has been found in differentiated enterocytes. 81,103,170 However, virus can also be detected in other tissues in the absence of viremia and has been shown to infect tissue macrophages. 103 Also, there is evidence of recurrent systemic spread. 103 These findings suggest that viremia and, ultimately, FIP can develop in infected animals at any stage after initial viremia, even when the virus is cleared from the intestine. 103
Pathologic Features of FCoV Infection and FIP
FIP
The name given to the disease in the 1960s acknowledges the consistent main gross pathological finding, a peritonitis (Figs. 3–9). Upon gross postmortem examination, FIP is typically characterized by a fibrinous and granulomatous serositis, protein-rich serous effusions, and/or pyogranulomatous lesions in several organs (Figs. 3–8). The latter lesions, however, are often very small and only identified by histological examination (Figs. 7, 8). Clinically, a rather clear distinction is made between an effusive (wet or nonparenchymatous) and a noneffusive (dry or parenchymatous) form of the disease, with a proportion of cases being considered in a transition stage between the 2 forms. 41,73,136 However, the postmortem examination often identifies extensive serosal and parenchymatous granulomatous lesions in organs alongside effusions of a variable quantity (Figs. 3–6), indicating that mixed forms are indeed more common than clinically appreciated.
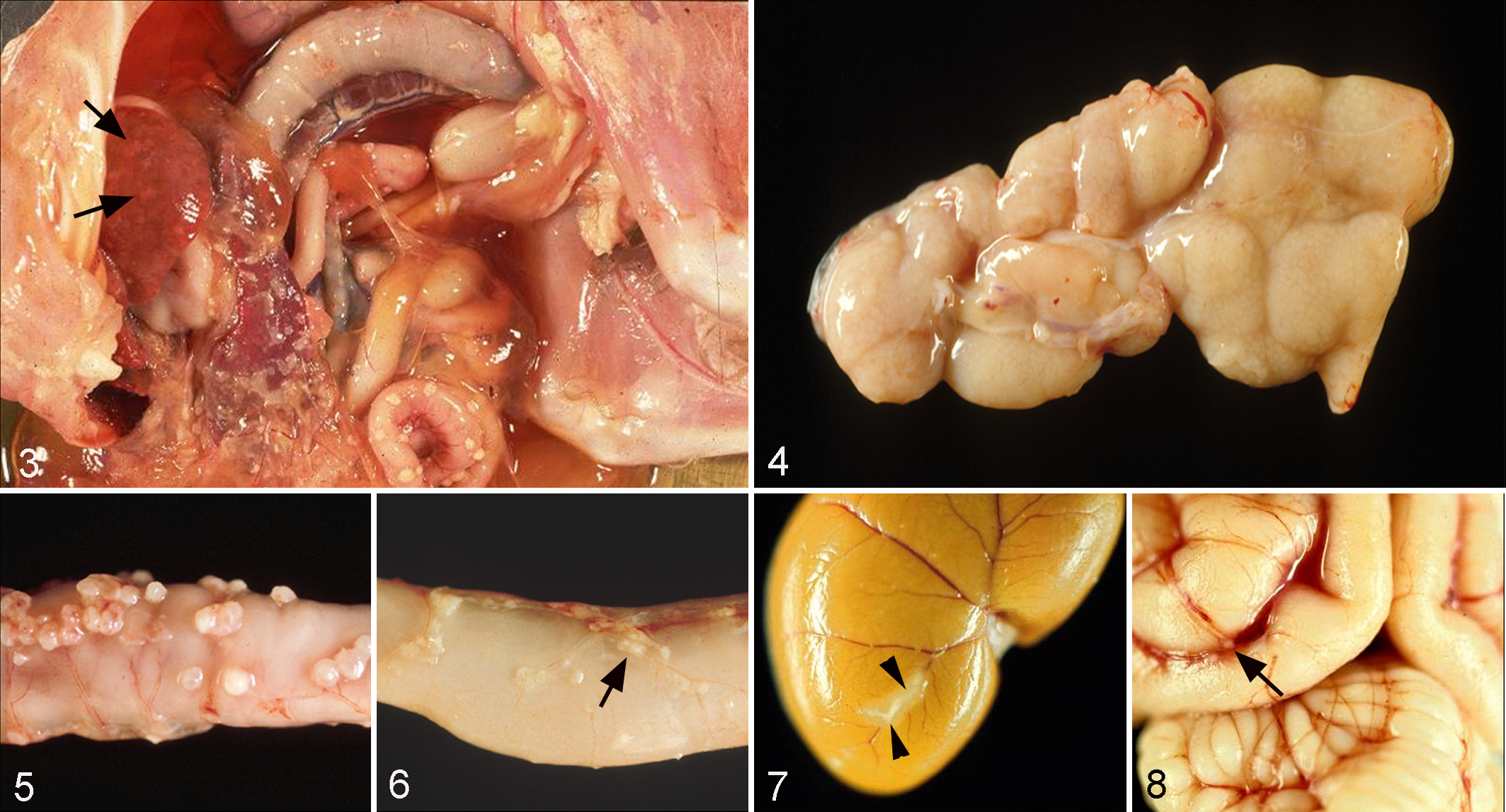
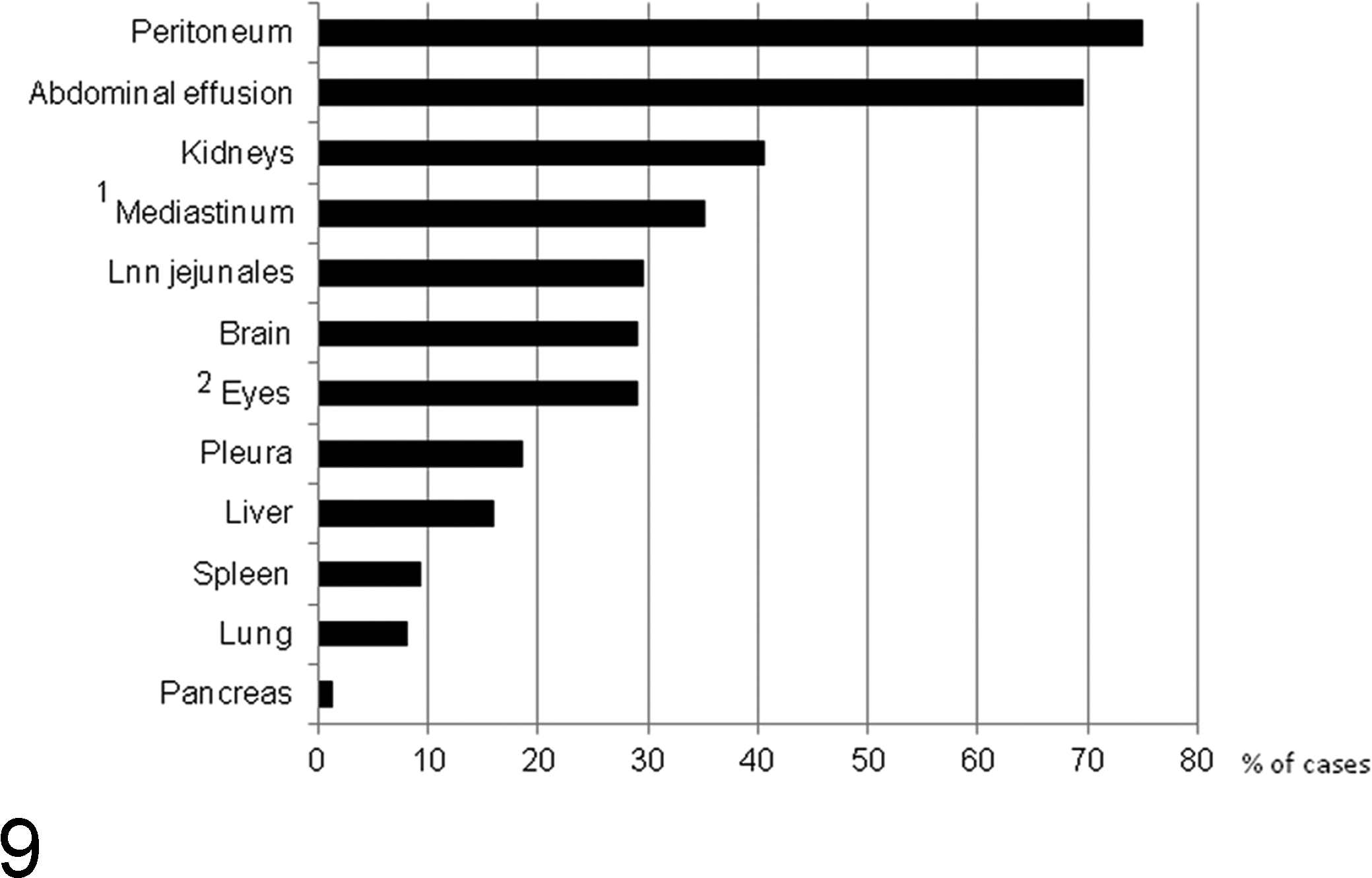
Distribution of lesions of feline infectious peritonitis (FIP) in cats that had undergone a thorough postmortem and histological examination for the diagnosis of FIP, including the histological examination of all major organs and tissues and confirmation of the disease by the immunohistological demonstration of feline coronavirus (FCoV) antigen. N = 77 (1 n = 54; 2 n = 86). Lnn jejunales, jejunal lymph nodes.
When the disease was first observed, several reports provided histopathological descriptions of both spontaneous and experimental cases. 19,76,77,172,175 –178 Several years later, a few studies attempted to categorize FIP lesions. 16,95,123 Based on distribution, cellular composition, and viral antigen expression, 4 types of lesions were described: diffuse alterations on serosal surfaces, granulomas with and without areas of necrosis, focal and perivascular B-cell and plasma cell infiltrates, and a granulomatous to necrotizing phlebitis; these can be found alongside each other. 95
The distribution of lesions varies in each individual case but shows a consistent general pattern. 95,98,136,178 Detailed gross, histological, and immunohistological examinations that we performed on a large cohort of diagnostic cases to identify all potential lesions confirmed peritoneal involvement in 75% of the cases, the majority (69%) associated with abdominal effusion (Fig. 9) and sometimes also with effusion in the thorax. Among organs, the kidneys were affected most often, followed by brain and eyes (Fig. 9). The eyes were always involved alongside the brain, and in a subsequent study that thoroughly examined confirmed FIP cases for the presence of ocular lesions, 29% (25/86) showed involvement of the eye, in the majority of cases (68%) bilateral (M. Weber, unpublished data), suggesting that the actual involvement of the eyes is generally underestimated. 136 FIP lesions are occasionally seen at unusual sites, such as the tunica vaginalis in cats with peritonitis, the skin, or the testicle. 23,33,149
Interestingly, the general distribution of the FIP vasculitis is relatively limited. Only veins are affected, most frequently the small- and medium-sized veins in leptomeninges, renal cortex (stellate veins), and eyes (mainly venules in iris, chorioid and retina) and, less frequently, veins in lungs and liver. 100
For natural FIP cases, an incubation period is not known. However, we gathered some information on the time lag between the onset of FCoV exposure and overt disease when we introduced several FCoV carriers into a large group of SPF cats housed together in a shelter-like multicat environment. The first clinical signs of FIP were detected after 6 weeks of exposure to the carriers. 109 After experimental infection, however, the incubation period has been shown to range between 2 and 14 days for the effusive and several weeks for the dry form. 36,128
The clinical course of FIP in natural cases is usually quite rapid for the wet form but can take several weeks for the dry form. 136 We observed a clinical course of 6 to 42 days (average, 14 days) from onset of clinical signs to death in the above-mentioned group of naturally infected SPF cats. 109 However, evidence of subclinical or protracted disease over a period of weeks to months has been reported. 136 Also, an experimental longitudinal study demonstrated recurring waves of clinical disease, where fever and weight loss coincided with T-cell depletion and increased viral loads in the blood. 36
From such data, histological findings suggestive of “disease waves” would be expected, and indeed, these can be observed. As reported, the typical serosal FIP lesions often exhibit an underlying layer of B cells and plasma cells, and some of the latter contain FCoV-specific antibodies. 95,174 We observed occasional natural cases with serosal lesions dominated by a thick plasma cell layer and evidence of granulation tissue formation with superficial granulomas and/or a fibrinous exudate (Figs. 10, 11). There is histological evidence that in granulomatous infiltrates of FIP, macrophages are progressively replaced by B-cells and plasma cells and that the typical FIP perivasculitis can develop into the focal and perivascular B-cell and plasma cell infiltrates that are frequently observed, for example, in leptomeninges and mesentery. 95,100 These findings provide further evidence that the humoral immune response can limit disease progression at least to some extent or for a limited time.
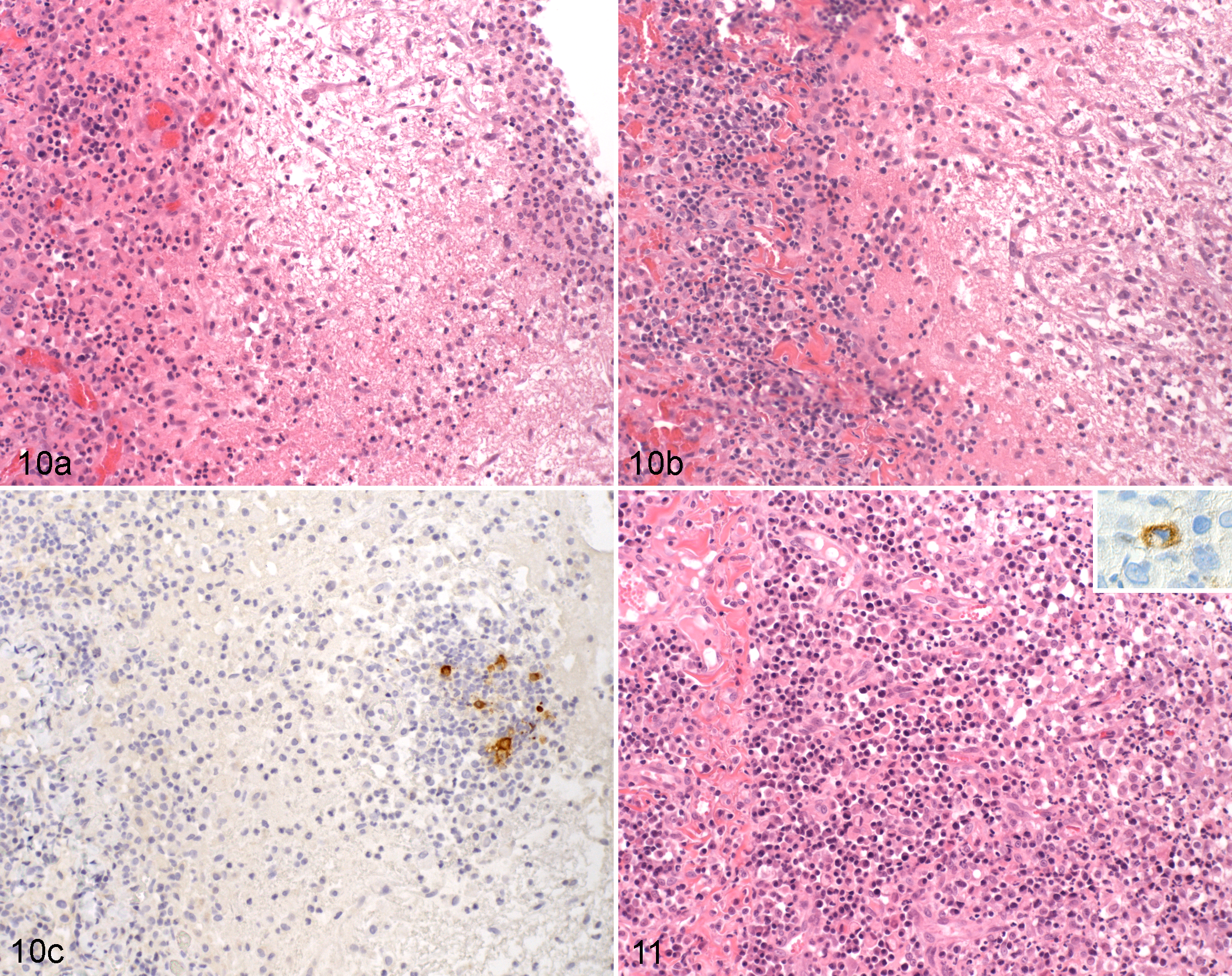
The typical FIP vasculitis is a phlebitis, mediated and dominated by activated virus-infected monocytes, with only few T-cells and neutrophils. On the basis of these features, it can be distinguished from an immune-mediated vasculitis, including an immune complex vasculitis. 100 However, in fulminant cases, necrosis of the vessels has been seen. 19,76,175 Interestingly, such an acute necrotizing inflammation can occasionally be observed in veins with apparent previous changes (Fig. 12), which further confirms the multiphasic nature of the disease. However, due to the morphological features of the acute vascular lesions, it also provides evidence that a type III hypersensitivity reaction does contribute to the pathogenesis at least in some cases. 19,76,115,175,176
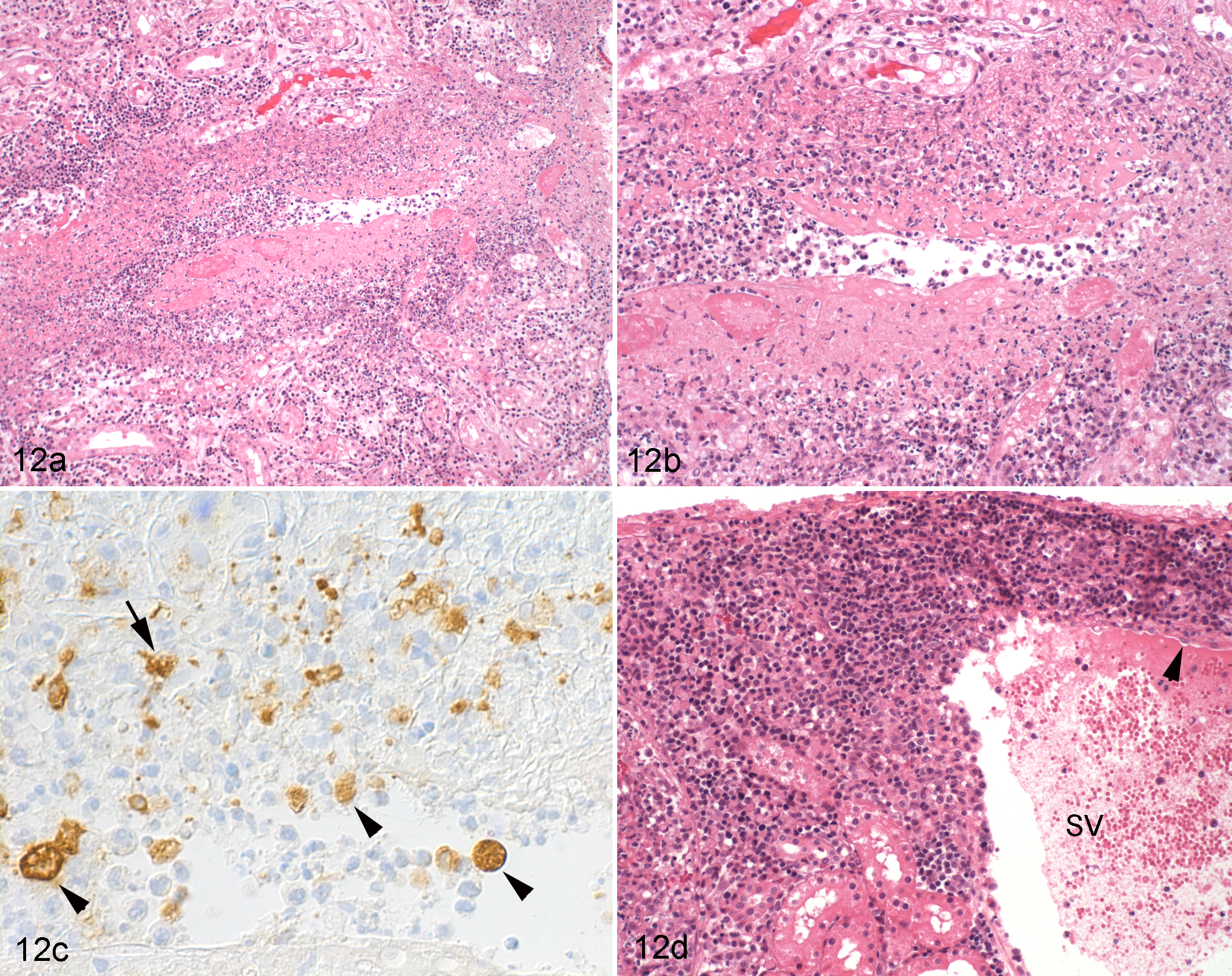
Kidney, cortex, cat, natural feline infectious peritonitis, dry form. (a) Stellate vein with fibrinoid necrosis and granulomatous inflammation, partly occluded by leukocytes. Hematoxylin and eosin (HE) stain. (b) Closer view of a, highlighting the necrosis of the vessel wall and the inflammatory infiltrate, dominated by often-degenerate macrophages. HE stain. (c) The same vein shown in b. Feline coronavirus (FCoV) antigen is present within monocytes in the vascular lumen (arrowheads) and in the inflamed vessel wall (arrow). Immunohistology for FCoV, as above. 95 (d) Stellate vein distant from that shown in a to c, exhibiting focal extensive perivascular plasma cell accumulation and activated endothelial cells (arrowhead). HE stain.
It has long been suspected that some cats can survive clinical FIP. 131 In the above-mentioned longitudinal experimental study, most cats died of FIP, but several animals that had undergone 1 or more episodes of clinical disease survived the 4-month study period and were seen to be “free of lesions” postmortem. 36 Similarly, in our longitudinal study undertaken on naturally infected SPF cats, a proportion of SPF cats became infected but survived the experiment despite consistent direct contact with virus-shedding carriers for 7.5 months, from the age of 21 weeks onwards. 97,109,113 The latter animals had remained clinically healthy throughout the experiment, apart from 2 cats that had developed uveitis. This did not resolve in 1 animal, which also showed transient nonspecific clinical signs. The postmortem examination of the latter revealed a moderate chronic B-cell– and plasma cell–dominated leptomeningitis and perivascular encephalitis as well as severe mononuclear conjunctivitis, iridocyclitis, and perineural leptomeningitis with FCoV antibody-positive plasma cells but no evidence of viral antigen in the infiltrates (Fig. 13). These findings support the results of an older experimental vaccine study and provide further evidence that lesions can remain limited and that macrophages in lesions can be replaced by B cells and plasma cells with time. 8,131
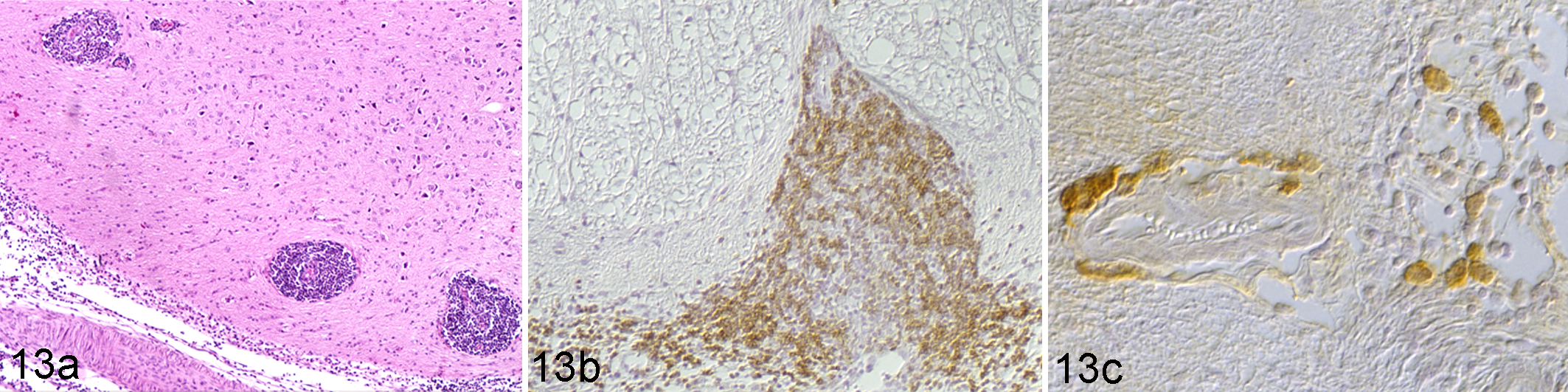
Brain, medulla oblongata. Specific pathogen-free cat euthanized without clinical signs and postmortem changes consistent with feline infectious peritonitis (FIP). The animal had been housed for 30 weeks with animals dying of FIP and had shown a clinical episode of central nervous system signs and ocular changes consistent with FIP. (a) Mononuclear perivascular infiltrates (arrows) in the white matter and diffuse infiltrates in the leptomeninges. Hematoxylin and eosin (HE) stain. (b) The infiltrate is dominated by CD45R-positive B cells. Immunohistology (IH), rat anti–mouse CD45R (clone B220/Ly5). 95 (c) The perivascular infiltrate as well as the leptomeningeal infiltrate contains plasma cells with feline coronavirus (FCoV)–specific antibodies. IH: DF-2 FIPV suspension, followed by mouse anti-FCoV (clone FCoV3-70). 95
Some animals can apparently confine the disease locally, at least for some time. One case series reported FIP lesions restricted to the mesenteric lymph nodes, and another described single mural intestinal lesions, partly in association with local lymph node involvement. 75,98 These findings suggest a strong local response to the virus. In the latter cases, however, progression to overt FIP was generally observed despite surgical removal of the intestinal lesions. 75
The lymphatic tissue of cats with FIP generally exhibits B- and T-cell depletion. However, in most cats, this occurs with previous follicular hyperplasia and is associated with markedly increased numbers, proliferation, and activation of macrophages in the splenic red pulp, lymph node sinuses, and bone marrow. 95,99,102
Coronavirus Enteritis
FECV is generally regarded as the avirulent pathotype of FCoV, and indeed, in older cats, oral FECV infection does not lead to any, or only very mild, nonspecific clinical signs, such as transient anorexia. 170 However, in young SPF kittens, at an age when animals would usually be protected by maternal antibodies, and after waning of maternal antibodies in non-SPF kittens, oral FECV infection can induce severe enteritis. 1,130 There have also been reports of fatal coronavirus enteritis in naturally infected juvenile and adult cats. Affected cats presented with catarrhal to hemorrhagic enteritis, and immunohistology confirmed that the virus infected the fully differentiated villous epithelial cells. 78,96,112
Coronavirus Infection Without FIP
In an environment of high infection pressure, such as an FIP outbreak, cats become FCoV infected and develop monocyte-associated viremia and a systemic immune response to the virus, as reflected by the development of antibody and circulating immune complex titers. 109,113 This is associated with distinct T- and B-cell hyperplasia in lymphatic tissues and the presence of plasma cells expressing FCoV-specific antibodies, which does not prevent virus shedding and viral spread to tissues. 97,99,103,113 A similar reaction of the lymphatic tissue has also been described after experimental FECV infections. 68,113 Interestingly, it is associated with macrophage proliferation in hemolymphatic tissues, similar to but less intense than that seen in cats with FIP. 102
Pathogenesis
The pathogenesis of FIP has been a research focus for several groups worldwide. Although the picture is still not clear, the results of both in vivo and in vitro studies, although sometimes controversial, have contributed more and more pieces to the puzzle. At present, 3 key features have been identified as essential prerequisites for the development of FIP lesions: systemic infection with virulent FCoV (ie, FIPV), effective and sustainable FIPV replication in monocytes, and activation of FIPV-infected monocytes.
Systemic Infection With Virulent FIPV
Two theories have been proposed for the infection of the host: the “in vivo mutation transition” or “internal mutation” hypothesis and the “distinct circulating avirulent and virulent strains” hypothesis. 21 The first model assumes that FIPVs arise in vivo from mutations of FECV in infected animals, and there is indeed strong evidence that the initially acquired FCoV of most cats is not an FIPV per se. Initial comparative genome analyses of FECV and FIPV laboratory and field strains has shown these to occur as closely related pairs. 79,129,167 Also, an experiment performed in chronically feline immunodeficiency virus (FIV)–infected cats showed that FIPV arise de novo from the FECV inoculum. 140 Furthermore, many studies demonstrated phylogenetic clustering of FIPV and FECV according to geographic distribution rather than disease phenotype. 10,27,137,140,167 Finally, it is well known that although FECVs are endemic in cat populations, FIP develops only sporadically, providing further strong evidence that FIPVs are generally not transmitted horizontally from cat to cat but emerge de novo in each cat that succumbs to FIP. 136
FCoVs exhibit in vivo genetic diversity, as shown by the frequent occurrence of viral quasispecies both in individual infected animals and in infected cats from the same household. 11,42,66,105 Indeed, experimental infection with fecal matter of naturally FECV-infected healthy cats leads to the occurrence of quasispecies in the large intestine of individual animals. 105 In natural infections, cats with FIP showed more extensive viral quasispecies formation than healthy animals, suggesting that a higher viral mutation rate is relevant for the generation of virulent mutants. 11 This is also supported by the fact that cats that develop FIP are less able to contain viral replication. 38 Surprisingly, however, recent phylogenetic analyses indicated that the observed genetic diversity mainly applies to type I FCoV, whereas type II viruses appeared to be less diverse. 42,111 The “distinct circulating avirulent and virulent strains” hypothesis is based on phylogenetic analyses and suggests that both virulent and avirulent strains circulate in the feline population and that, independently of geographic location, sequences tend to cluster with disease phenotype. 20 The occurrence of occasional FIP epidemics indicates that this theory applies sporadically.
Effective and Sustainable FIPV Replication in Monocytes
The second essential prerequisite for FIP appears to be the viral capacity to replicate effectively and sustainably in monocytes of the infected host. In vitro, both FECV and FIPV can replicate in isolated feline peritoneal macrophages, BMDMs, and monocytes, but only FIPVs undergo sustainable replication and spread the infection in the culture. 38,145,154 Moreover, cats exhibit individual differences in the sustainability and susceptibility of monocytes to infection, indicating that host factors also play a role. 38
These results support in vivo studies that have shown that FCoV infection generally leads to monocyte-associated viremia but that viral replication in the blood (ie, in monocytes) and viral loads in tissues are generally significantly higher in association with FIP. 64,87,101,113,150 On the basis of older experimental studies, Pedersen 136 suspected that viral replication in monocytes is very slow, at least during the first 2 weeks after FIPV infection, but then increases rapidly, around the time when specific antibodies occur. 125,176 So far, this hypothesis has not been tested in experimentally infected cats. However, it has been shown that FCoV infection of cats induces macrophage/monocyte proliferation in hemolymphatic tissues. 99,102 We did not find this to be associated with upregulation of cytokines that stimulate macrophage proliferation in these tissues; it could therefore represent a systemic effect of infected monocytes that we found to transcribe granulocyte-monocyte colony stimulating factor (GM-CSF) and interleukin (IL)–6, cytokines that both induce proliferation and differentiation of monocyte and neutrophil precursors, within hours after in vitro FIPV infection (A. Kipar, unpublished data). The proliferation of monocytes/macrophages likely ensures the supply of viral target cells (ie, mature circulating monocytes or tissue macrophages). Proliferating macrophage populations, such as macrophages in the splenic red pulp or myelomonocytic cells in the bone marrow, appear not to allow viral replication, since viral antigen cannot be detected in these cells. 99,103 Higher blood cytokine levels due to cytokine release from a larger number of infected monocytes and from macrophages in FIP lesions could account for the more pronounced proliferation and generalized activation of macrophages in hemolymphatic tissues observed in cats with FIP. 102
Based on the fact that FECV can also replicate at least briefly in monocytes, it was recently suggested that monocytes, rather than the intestinal epithelial cells, might be the cells in which the FECV-FIPV mutations occur. 138 Given the high mutation rate of the virus, this would allow positive selection for macrophage tropism and progressive viral adaptation to replication in monocytes/macrophages. 138,154 It would also suggest that viral clearance from the blood and even the intestine might not prevent recurrent viremia and possibly even the development of FIP, since persistently infected, healthy, nonviremic FECV carriers were found to bear virus in tissue macrophages (ie, in sinus macrophages in mesenteric lymph nodes and in pulmonary intravascular macrophages). 103 This indicates that macrophages in the intestine take up the virus from enterocytes and carry it to the regional lymph nodes and eventually in the blood. Viral RNA was also detected in the liver, where the virus most likely infects Kupffer cells. 103,126 Both pulmonary intravascular macrophages and Kupffer cells phagocytose particles from the blood but could also replicate and release virus into the circulation or transmit it to monocytes. If the mutation and transformation of FECV to FIPV can take place in these macrophages, this could result in FIP at any time after initial infection. 138
Activation of FIPV-Infected Monocytes
The morphological hallmark and initiating lesion of FIP is a granulomatous phlebitis and periphlebitis that is mediated by highly activated monocytes, most likely during a phase of high-level monocyte-associated viremia with substantial viral replication. 87,100,150 Studies on natural cases have shown that the phlebitis develops through direct interaction between monocytes and activated endothelial cells. The monocytes strongly express cytokines, such as tumor necrosis factor (TNF)–α and IL-1β, and adhesion molecules, such as CD18, that allow their interaction with activated endothelial cells and express enzymes, such as matrix metalloproteinase-9, which dissolve the vascular basement membrane at sites of monocyte emigration. The endothelial cells appear systemically activated, and the restrictive distribution of vascular lesions (ie, affecting veins and only in selected organs) is likely a consequence of selective responsiveness of the endothelium. 100 A recent flow cytometry study showed that not only monocytes but also T- and B-cells exhibit adhesion molecule upregulation in association with FIP. 119
The observed simultaneous, generalized activation of both vascular endothelial cells and macrophages in hemolymphatic tissues could be mediated by activated monocytes alone, provided they release sufficient amounts of cytokines. 100,102 The latter appears likely, considering also that cats with FIP show increased vascular endothelial growth factor (VEGF) transcription in (virus-infected) monocytes and increased serum VEGF levels. 161 Furthermore, peritoneal exudate cells of cats with FIP exhibit high TNF-α mRNA levels and were previously shown to release IL-1β and IL-6, and even alveolar macrophages collected by bronchoalveolar lavage from FIP cats show significant upregulation of TNF-α, GM-CSF, granulocyte (G)–CSF, IL-6, and other B-cell differentiation factors, all suggesting strong generalized monocyte/macrophage activation in response to FIPV. 59,60,155,157,158
What ultimately triggers the fulminant monocyte activation in infected cats is not yet known. However, FIPV infection of the monocytes is apparently an essential prerequisite, which was recently shown in vitro in isolated feline monocytes and macrophages. FIPV rapidly induced activation of the p38 mitogen-activated protein kinase (MAPK), which directly regulates the expression of proinflammatory cytokines via phosphorylation of a range of signaling molecules in PBMCs, likely early during entry and, although less intensely, between 6 and 12 hours PI (HPI), when virus is being produced. 38,107,142 This was associated with the induction of TNF-α and IL-1β but not IL-6 production, as demonstrated in the PBMC supernatant at 24 HPI. 142 VEGF transcription was shown to be significantly upregulated at 48 HPI in isolated feline monocytes and alveolar macrophages, and feline alveolar macrophages showed increased TNF-α production at 48 and 72 HPI, all exclusively in association with viral replication. 160 A similar increase was seen for G-CSF and GM-CSF transcription at 72 HPI; however, only upregulation became significant, and TNF-α and VEGF levels were further increased when cells were inoculated with virus in combination with an antibody against the FCoV S protein that is known to induce ADE. 152,157,158,160
The Immune System in FIP and FCoV Infection
With the aim to identify the role of the immune system in the pathogenesis of FIP, the transcription of cytokines in the blood (IL-4, IL-6, IL-10, IL-12 p40, IL-18, IFN-γ, TNF-α) was monitored in experimentally infected animals. A first study reported an initial mild increase in IL-6 and IFN-γ transcription in PBMCs that correlated with transient pyrexia, followed by a drop of all other examined cytokines and IFN-γ, possibly as a consequence of the lymphopenia that developed simultaneously. 65 A second study demonstrated TNF-α upregulation during the development of FIP. 106 Another group then screened serum VEGF levels by enzyme-linked immunosorbent assay (ELISA) and found an increase in association with body effusions. 160 In natural FIP cases, at the time of death, the blood showed very high interindividual variation in cytokine mRNA levels; however, IFN-γ mRNA was generally scarce or absent. 54 Blood IFN-γ levels of FIP cats were then found to be similar to those of healthy FCoV infected cats, but high IFN-γ concentrations were present in effusions. 57 These were considered a likely consequence of the observed IFN-γ transcription within lesions. 16 Believed to be released by the T-cells in the lesions, IFN-γ could be responsible for macrophage attraction and local activation, which would also enhance Fc receptor expression on their surface and thereby virus uptake (via antibody to FCoV) and replication. 16 In light of the above results, the generally variable pathological changes, and in particular the apparent multiphasic nature of the disease, it is possible that the fulminant monocyte activation, which is essential for the development of FIP vasculitis, does only occur as brief bouts, followed by a phase in which self-sustained granulomatous lesions develop. Such cytokine peaks might be missed when PBMCs or whole blood are sampled on a regular basis or immediately prior to death.
FCoV infection of cats, regardless of the development of FIP, initiates a humoral immune response, as evident by the development of antibody titers and, morphologically, the formation of secondary follicles in lymphatic tissues, as well as by the presence of FCoV-specific antibodies within plasma cells both in lymphatic tissues and in older FIP lesions. 41,95,97,99,102,170 Some cats can eliminate the infection and then become reinfected, with a different or the same virus strain, but tend not to develop FIP as a consequence of this. 4,5,50 It is assumed that protective immunity is mainly cell mediated, and there is evidence that the development of high anti-S titers and a high S:M antibody ratio leads to virus clearance. 62,136 However, in many animals, the antibodies that are formed, and in clinical FIP often with very high titers, are not able to eliminate the virus and/or the infected cells.
In contrast, experimental studies provided evidence that ADE plays a role in FIP. 127,132,176 FIPV ADE is based on the enhanced uptake of virus and anti-S antibody by macrophages/monocytes via the Fc receptor and can occur in vivo when immunized cats are infected with FIPV of the same serotype. 29,117,156 However, it has been seen only with some highly virulent FIPV and appears to be of less relevance in the natural disease. 2,136
While ADE could explain the rapid spread of infection in the monocyte/macrophage population, it would not explain why the infected cells are not eliminated by the immune system. The latter can be explained by recent in vitro studies. In the presence of specific antibodies, FCoV-infected isolated feline monocytes rapidly internalize the viral glycoprotein that is expressed on the surface as antigen-antibody complexes. 39,40 The internalization is mediated by the S and M proteins and, interestingly, does occur with both FIPV and FECV. FIPV also inhibits the complement-mediated lysis of infected cells, even if they express viral antigen on their surface. 30 This process is independent of the accessory 3 and 7 proteins, but the underlying mechanism has not yet been identified. 30 Nonetheless, the internalization of FCoV proteins by infected cells may play a role in the pathogenesis of FIP, since the lack of an immune response that eliminates infected cells will allow more pronounced virus production and/or a quiescent infection state. 39 This phenomenon could also allow the virus to persist in tissue macrophages. 103
As mentioned, there are marked differences between the composition and activity of the hemolymphatic tissues in FCoV-infected cats with and without FIP. Whereas healthy carriers exhibit distinct lymphoid hyperplasia with lymphocyte proliferation, FIP cats show depletion that is morphologically reflected in the often markedly depleted, although mainly secondary, lymphatic follicles, the depletion of T-cell zones, and in particular the generally marked thymus atrophy in young cats. 36,97,99 This is mainly a consequence of lymphocyte apoptosis, and TNF-α expression by lymphocytes, particularly in lymphatic tissues with FIP lesions, might be the underlying mechanism. 32,68,99 The findings are matched by the marked blood lymphopenia observed in the terminal stage of FIP and a persistent drop in circulating CD4+ and CD8+ T-cell numbers as well as an increased rate of apoptosis in PBMCs over the course of the disease. 32,36,155
A recent transcriptional profiling study on Crandell Rees feline kidney (CRFK) epithelial cells identified upregulation of genes associated with monocyte and T-cell immune response, apoptosis, cell cycle, cytokine, and ubiquitin activities early (3 hours) after FIPV infection. A similar trend was observed in cats with clinically diagnosed FIP. 74 Among PBMCs, mainly CD8+ cells were shown to be affected by apoptosis, and peritoneal exudate cells and PBMCs of cats with FIP seem to produce the necessary TNF-α to mediate this. 155 An in vivo study on experimentally infected cats confirms this finding, as it demonstrated increased TNF-α transcription in PBMCs of cats that developed FIP, whereas animals that remained healthy had low TNF-α and high IFN-γ mRNA levels. 106 High IFN-γ transcription, together with an increase in IL-1β, was also seen in the blood of naturally infected healthy FCoV-infected cats regardless of antibody titers and virus shedding, prior to the occurrence of FIP cases in the catteries. This was considered a consequence of an increase in CD8+ T-cells and coincided with the release of acute phase proteins, indicating that production of these cytokines by PBMCs contributes to the protection of FCoV-infected cats against FIP. 54
Despite the obvious functional differences in the T- and B-cell component, FCoV infection is associated with proliferation and activation of monocytes/macrophages and their precursors in hemolymphatic tissues of infected cats and, to a higher degree, cats with FIP. 99 In an attempt to identify the underlying mechanism, hemolymphatic tissues of FCoV-infected cats with and without FIP were assessed for the transcription of cytokines that mediate macrophage activation. 102 SPF cats served to establish constitutive transcription levels, which highlighted the general flaw of any such “global expression” study: the high variability in individual transcription levels even in sex- and age-matched SPF animals, let alone a group as heterogeneous as naturally infected cats with FIP with regard to age, sex, disease type, and stage. 54,102 Nonetheless, some relevant differences in the cytokine transcription were identified. Naturally infected healthy cats exhibited significantly higher IL-10 transcription levels in the spleen and lower IL-6, G-CSF, and M-CSF levels in mesenteric lymph nodes than cats with FIP, whereas FIP was associated with significantly lower IL-12 p40 mRNA levels in lymphatic tissues. 102 A similar trend of reduced IL-12 p40 transcription was found in lesional vs virus-free mesenteric lymph nodes of cats with FIP and in brains with FIP lesions. 32,52 This suggests that an effective immune response together with an IL-10–mediated limitation of macrophage activation and increased cellular cytotoxicity allows infected cats to limit the viral infection and remain healthy, whereas a lack of IL-12 inhibits an effective immune response and allows monocyte/macrophage activation and ultimately FIP, probably as a consequence of impaired T-cell–mediated macrophage activation. 102
The fact that cytokines produced by macrophages and known to activate and induce proliferation of these cells (such as G-, M- and GM-CSF; IL-6; and TNF-α) were not significantly upregulated in these tissues despite the presence of abundant activated macrophages was considered further evidence of a systemic effect (ie, the release of the relevant cytokines by infected, activated monocytes). 102 The results of a recent study on cats with natural FIPV infection align with this theory, since they provide further evidence that diseased animals cannot mount an effective immune response: cats with FIP showed depletion and reduced cytotoxic activity of natural killer cells in blood and lymphatic tissues, together with a reduction in regulatory T-cells, which will decrease the animal’s capacity to inhibit the inflammatory/immunopathological processes. 168
Several attempts have been made to match the pathogenesis of FIP with known immunopathogenic mechanisms. For example, FIP has long been considered an immune complex–mediated type III hypersensitivity disease. This is based on the demonstration of cell-free fibrinogen and C3 as well as viral antigen, IgG, and complement within leukocytes in vascular and focal granulomatous-necrotizing lesions. Also, cats with FIP exhibit FCoV-specific immune complexes in blood and glomerula and show high γ-globulin and C3 serum levels. 88,89,122,127,176 However, circulating FCoV-specific immune complexes not only are found in diseased animals but can also be detected, at least transiently, in infected cats that remain healthy. 95,112 Also, the typical FIP vasculitis does not show features of immune complex vasculitis, such as the involvement of arteries and the dominance of neutrophils. 100 While this does not confirm a type III reaction as the essential pathogenic mechanism, it does not exclude its contribution to the disease, for example, in its acceleration (see above).
Other authors have considered a type IV hypersensitivity reaction as the basis for the development of the granulomatous lesions, due to the dominance of CD4+ cells in the otherwise macrophage-dominated lesions. 16,123 In any case, the available data provide definite evidence that immunity to FIP is cell mediated, and it has been postulated that it requires viral persistence. 136 Furthermore, there is evidence from both experimental and natural infections that an effective early T-cell response to FCoV is essential for the prevention of FIP, since it appears to ensure the limitation of viral replication. 36,124
There has been a lot of speculation about why cats develop the dry, the wet, or a mixed form of FIP, and it is widely agreed that a strong humoral together with a very weak cellular immunity leads to the wet form, with lesser effusion with increasing cellular immunity. 136 However, a recent study provided evidence that the effusions are also initiated by FIPV-infected monocytes/macrophages, since the latter have been shown to produce VEGF. 160 VEGF is a very strong mediator of vascular permeability, which also has this effect on feline vascular endothelial cells. 44,160 Also, serum VEGF levels were seen to correlate with the quantity of body effusions. 160 These findings do not necessarily contradict the existing theories but are very much in line with all other data that render the infected, activated monocyte the key mediator of the disease.
Several studies have provided data that indicate individual differences in susceptibility to FCoV infection in general and to the effect of FIPV on monocytes. Molecular studies found evidence that individual cats can be entirely resistant to FCoV infection and that the monocytes of some cats can be completely resistant to FCoV, or to FECV alone, while the monocytes of some cats are more prone to productive FCoV infection than those of others. 4,5,38,162 Finally, there appear to be differences in FIPV serotype I internalization in individual cats, suggesting that its receptor is expressed differentially between cats. 165 In an attempt to identify genetic factors that could predispose cats to FIP, a genome-wide association study analyzed a wide range of single-nucleotide polymorphisms (SNPs) in large cohorts of healthy Birman cats and those with FIP. 61 Despite the use of a highly inbred population, 5 potential candidate genes involved in processes such as immune regulation and apoptosis were identified, but no single association was 100% concordant with the disease phenotype. 61 At present, the knowledge is still lacking as to the reason for the individual differences and the key event that blocks the capability of monocytes to inhibit virus production.
Diagnosis of FIP
The postmortem diagnosis of FIP relies on a combination of gross and histological examination, in combination with the demonstration of viral antigen in the lesions. 58 In contrast, the noninvasive antemortem diagnosis of FIP still remains a challenge, especially in the dry form of the disease. 73,98,136 A combination of indirect and/or direct virus detection with evaluation of blood hematological and chemical parameters as well as medical history and clinical signs, the so-called FIP algorithm, is so far the best predictor of disease. 6
Host Blood Parameters
Common blood alterations include lymphopenia, mild to moderate regenerative anemia, and hyperproteinemia due to hypergammaglobulinemia. 73,136 Other laboratory parameters, such as liver enzymes, bilirubin, urea, and creatinine, might be helpful, but high values merely reflect organ damage, which is most likely a consequence of FIP lesions. 6
Several studies showed that more than the single parameters, the serum albumin to globulin ratio (A:G ratio) has a high diagnostic value, and at values above 0.8, FIP is extremely unlikely. 6,72,151 A recent retrospective study evaluating the A:G ratio showed a very poor positive predictive value (PPV) even for a cutoff value of 0.6. However, the negative predictive value (NPV) was 100% and 99% for an A:G ratio of >0.8% and >0.6%, respectively. 91 It is important to mention that PPV and NPV very much depend on the prevalence of the disease in the tested population, and in the latter report, the study population was small and the disease prevalence much lower than in previous ones. 6,72,91,151
Since the first report addressing the diagnostic values of acute phase proteins, many studies have focused on the acute phase protein, α1 acid glycoprotein (AGP). Serum levels are highly elevated in cats with FIP (>3 mg/ml) but are also high in other inflammatory conditions or neoplastic diseases, such as lymphoma. 25,31,43,146 Furthermore, AGP levels may also rise in asymptomatic FCoV carriers, especially from households with endemic infection. 55 However, when interpreted alongside pretests (ie, epidemiological factors, clinical information, and FCoV serology), moderate AGP increases are useful discrimination parameters when the probability of FIP is high, whereas with low FIP probability, only very high AGP levels support the diagnosis of FIP. 125 A recent retrospective study found complete concordance between AGP levels and immunohistology in a small series of challenging diagnostic cases. 58
Analysis of Effusions
The presence of effusions facilitates the diagnosis, since tests on effusions have a higher diagnostic value than blood tests. 6,72 FIP effusions typically have a very high protein content (>35 g/l) but a low cellularity (<5000 nucleated cells/ml), with a dominance of macrophages and neutrophils. When sufficient cells are present, the demonstration of viral antigen in macrophages confirms the diagnosis with a very high PPV. 22,72,73
The Rivalta test, commonly used to differentiate between FIP effusions and effusions due to other diseases, is not very specific. The high protein content, including fibrin and inflammatory mediators, in FIP effusions normally induces a positive reaction. However, a recent study on a large cohort of cats with effusions has shown that, while it has a high NPV, this test has a lower sensitivity, specificity, and PPV than previously reported. 48,73
The A:G ratio can also be measured in effusions; this test has a high PPV if the ratio is <0.4 and a high NPV if the ratio is >0.8. 147 Also, very good correlation exists between AGP values in effusions and serum. 14 The demonstration of FCoV-specific antibodies in the effusions is only meaningful when the titer is high (≥1:1600), whereas the absence of antibodies has a good NPV. 72
Indirect Virus Detection: Serology
Serology, based on the detection of FCoV antibody titers by a range of methods (immunofluorescence, ELISA, rapid immunomigration), is widely used commercially to assist in the diagnosis of FIP and for quarantine purposes. The tests are applied to blood and effusions, and apparent false-negative results are a known problem. A recent study addressed this issue and showed a correlation of lower antibody levels in samples containing higher amounts of virus, as shown by quantitative reverse transcription polymerase chain reaction (RT-PCR). 114 It was hypothesized that these false-negative results were due to antibody binding to virus in the sample instead of the virus in the serological tests. 114
Also, a high percentage of healthy FCoV carriers are antibody positive, of which only a small percentage develops FIP. 1,151 Very high titers (≥1:1600) in combination with pretests that suggest FIP indicate an increased likelihood of FIP, unless obtained from animals in an endemic environment, such as multiple-cat households. 72,135 It should also be noted that different methodological approaches and even different laboratories might yield different results from the same sample, depending on the antigen used. 73 Serology is generally considered a useful tool for the screening and management of catteries and quarantine purposes, particularly since antibody titers are correlated to shedding intensity and frequency. 41
The detection of circulating antigen-antibody complexes, for example by a competitive ELISA on serum, was shown to have a PPV of 67% and an NPV of 84% for FIP. 72,88 However, healthy FCoV carriers can also show circulating immune complexes. 113
Direct Virus Detection
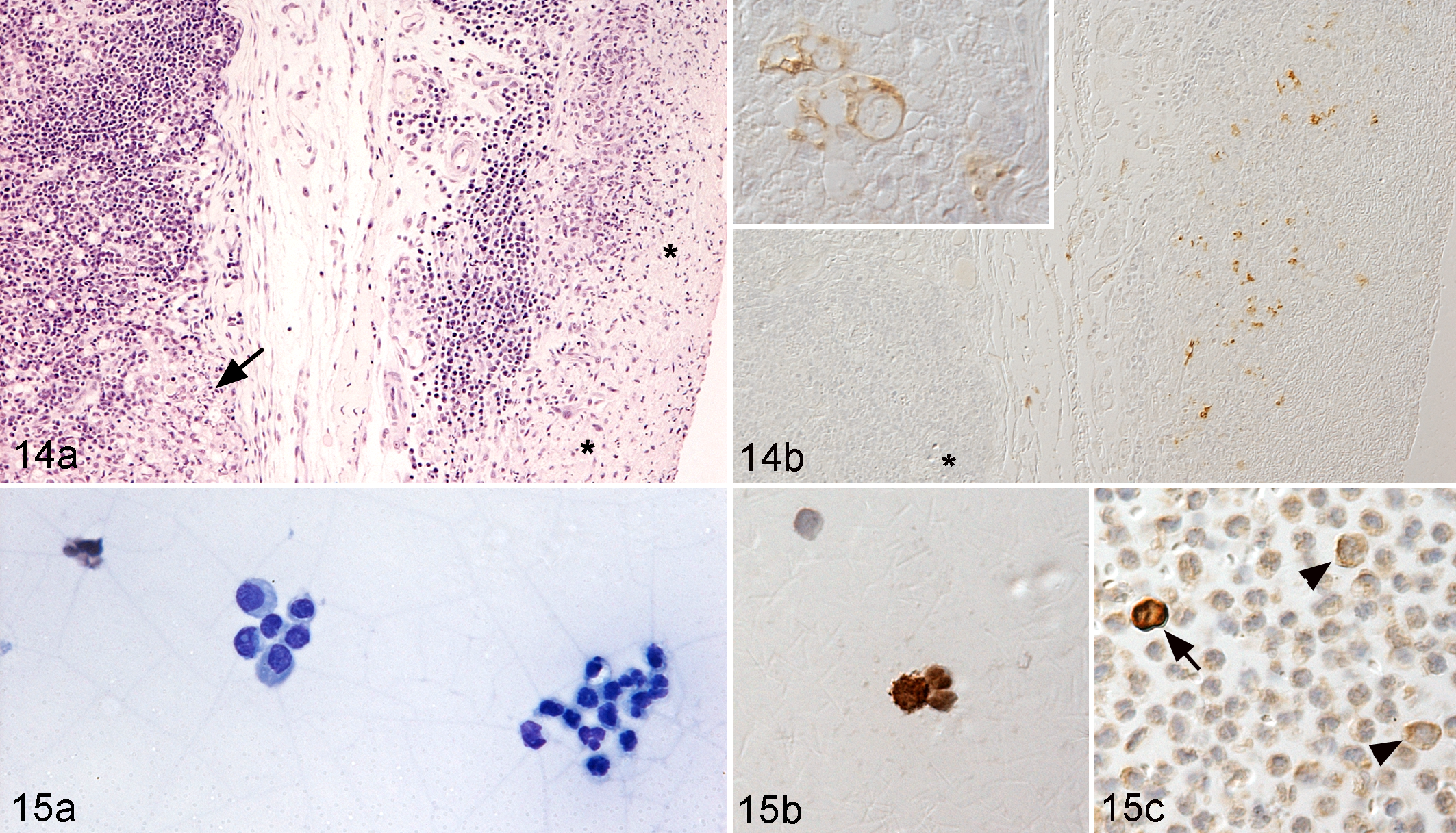
Immunohistology has been used for 2 decades to detect FCoV antigen in lesions and has a very high PPV. 22,95,123,161 It is therefore considered the gold standard and by many diagnostic pathologists an essential component of the definite diagnosis of FIP, particularly in histologically inconclusive cases. 41,58,136 For the definite antemortem diagnosis, optimally, surgical biopsies of granulomatous lesions are used (Fig. 14), while random Tru-cut biopsies or fine-needle aspirates are often not helpful. 56 The demonstration of FCoV antigen in macrophages in effusions, as mentioned above, is an alternative, noninvasive tool for the intravitam diagnosis of FIP. A positive result, obtained in the context of appropriate positive and negative controls, is highly predictive of FIP, whereas a negative result does not exclude FIP. 22,72 The latter is due to both the low cellularity of the effusions and the relatively low sensitivity of the method, which can detect only heavily virus-laden cells. In the authors’ experience, based on the parallel staining of cytological preparations and formalin-fixed, paraffin-embedded cell pellets prepared from the effusions (minimum 1 ml), more reliable results can be obtained from the concentrated cell preparations in the pellets (Fig. 15).
RT-PCR, especially real-time RT-PCR, is a sensitive method to detect virus RNA in different samples, such as feces, blood, effusions, and tissues of FCoV-infected cats and those with FIP; however, these cannot differentiate between the pathotypes. 46,67,80,105 The detection of FCoV RNA in the feces is mainly used for management purposes in catteries (ie, to determine the kinetics of viral shedding). 4,41 FCoV is known to spread systemically with infection, regardless of the development or presence of clinical signs of FIP; therefore, diagnostic tests that identify viremia can only be used to support other tests toward a diagnosis of FIP in cats with relevant clinical features. 41,64,113 The detection of virus in effusions, however, has proven to have a high PPV, but a negative result does not exclude FIP. 53,164
A recent study identified 2 alternative amino acid differences in the putative fusion peptide of the S protein in FECV and FIPV and confirmed that together these 2 substitutions distinguish FIPV from FECV in >95% of cases. 28 Although it cannot be excluded, due to the quasispecies nature of FCoV, that other mutation patterns could lead to disease, this is so far the most promising potential diagnostic tool that involves the direct virus detection. 11,42,66,105 Nevertheless, as these alterations are not present in the virus shed in feces by cats with clinical FIP, any routinely employed diagnostic PCR test based on this result will need to overcome the poor sensitivity of this method in the blood to be conclusive. Also, as a commercial test, a protocol would be preferable that could directly detect these mutations without the need of a further, time-consuming sequencing step.
A recent publication takes previous attempts to correlate FIPV replication in monocytes with FIP further and suggests a new methodological approach for the robust simultaneous detection of virus replication and viral load, by a real-time PCR based on primer-probe energy transfer. 24,87,150 Rather than a tool to identify infected cats, the authors proposed this method for the identification of persistent shedders and thereby the potential sources of emerging FIP variants. Since the test was also able to reliably detect virus replication in FIP effusions, it might be useful for the confirmation of FIP at least in the wet form, provided the results are confirmed on larger case cohorts. 87
With increasing knowledge of the pathophysiological mechanisms that drive the virus-host interaction in FCoV infection and with the constant improvement of molecular techniques, there is reasonable hope that in the near future, the diagnostic tools for the diagnosis of FIP can be refined to specifically detect FIPV and to integrate the assessment of more host response parameters tailored to FIPV.
Footnotes
Acknowledgements
We thank those who encouraged us and consistently supported our work on FCoV infection and FIP for several years, Professor Hans Lutz, formerly Clinical Laboratory, Vetsuisse Faculty, University of Zurich, Switzerland, and Professor Manfred Reinacher, Department of Veterinary Pathology, Faculty of Veterinary Medicine, University of Giessen, Germany. We are also grateful to Dr Enrico E. Ortelli for his help with the schematic drawings.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.