
Abstract
Infection with high-pathogenicity avian influenza virus (HPAIV) has been associated with a wide range of clinical manifestations in poultry, including severe depression in egg production and isolation of HPAIV from eggs laid by infected hens. To evaluate the pathobiology in the reproductive tract of chickens, adult hens were inoculated intranasally with 3 HPAIV strains. All 3 strains induced lesions in the reproductive tract 36 to 72 hours after inoculation. Positive immunostaining was observed in all segments of the reproductive tract, occurring predominantly in stromal cells and superficial germinal epithelium of the ovary, in mucosal epithelial cells and less often glandular epithelium throughout the oviduct, and in vascular endothelium. This study generates important data and explains previously reported virus isolation from yolk, due to ovarian virus replication, and virus recovery from albumin, due to virus replication in epithelial cells in several segments of the oviduct.
Avian influenza virus (AIV) is an important infectious agent that has been associated with a wide range of clinical manifestations in birds and mammals worldwide. 7 Although infection by low-pathogenicity AIV in domestic poultry can cause subclinical disease or mild respiratory signs with decreased egg production, infection with high-pathogenicity AIV (HPAIV) has been associated with severe systemic disease and accompanied by high mortality rates. 7 HPAIV replicates systemically in chickens, 3,4,6,7 and virus isolation has been reported to occur from the eggshell surface as well as within the egg yolk and albumin after experimental infection. 9 The presence of virus in the egg components and the decrease in egg production described in outbreaks of HPAIV in poultry 4 –6 suggest that the virus can successfully replicate in the reproductive tract of affected individuals. However, very little is known regarding the pathobiology of HPAIV infection in the reproductive tract of chickens. The objective of this study was to determine the pathologic changes and immunostaining pattern of HPAIV infection in the different cell types and anatomic parts of the reproductive tract of hens following experimental intranasal infection with 3 different HPAIV strains.
Thirty adult White Leghorn chickens in egg production were divided in 4 groups and inoculated with 106.0 mean embryo infectious doses (EID50) of HPAIV in 0.1 ml into the middle nasal cavity: (1) group 1 (n = 12) received A/chicken/Pennsylvania/1370/83 H5N2 (PA/H5N2); (2) group 2 (n = 8) received A/chicken/Vietnam/NCVD-675/2011 H5N1, clade 2.3.2.1 (Vn/H5N1); (3) group 3 (n = 8) received A/chicken/Egypt/1063/2010 H5N1, clade 2.2.1 (Egy/H5N1); and (4) group 4 (n = 2) served as noninoculated controls. The number of birds per group was determinate based on previous studies that indicated that Vn/H5N1 and Egy/H5N1 induce mortality around 48 hours after challenge, whereas PA/H5N2 takes 60 to 72 hours to induce high mortality. The larger group for PA/H5N2 was selected to ensure tissue sampling during the whole disease development. The study was performed in biosecurity level 3 enhanced facilities at Southeast Poultry Research Laboratory, and all personnel were required to wear a high-efficiency particulate air–filtered respirator (3M, St Paul, MN). General care of hens was provided in accordance with the Institutional Animal Care and Use Committee, as outlined in the Guide for the Care and Use of Agricultural Animals in Agricultural Research and Teaching.
All birds were observed every 12 hours postinfection (PI) for clinical signs. Oral swabs and tissues samples were collected from 2 sick or dead birds per inoculated group every 12 hours PI (every bird was swabbed only once, at the time of death or euthanasia); if no sick or dead bird was identified, 2 random birds were selected and euthanized for swab and tissue sampling. The 2 control chickens (group 4) were euthanized at the beginning of the experiment, before challenge. Euthanasia was performed with intravenous sodium pentobarbital. Ovary, infundibulum, magnum, isthmus, shell gland, vagina, and cloaca were collected to assess reproductive tract changes, and nasal cavity, spleen, heart, and brain were collected to confirm systemic infection. All swabs were collected in brain-heart infusion (BHI) media and tested for virus using quantitative real-time polymerase chain reaction (PCR) according to a previous protocol. 8 Virus quantity was calculated by converting CT values to mean embryo infectious dose (EID50/ml) equivalents as previously reported. 8 All sampled tissues were fixed in 10% buffered formalin (from 2–5 days), processed for histology by routine procedures, and stained with hematoxylin and eosin (HE). Pathologic changes were qualitatively and quantitatively evaluated in each portion of the reproductive tract (Table 1). Replicate sections of the reproductive tract were stained immunohistochemically as previously reported 10 to identify the site of virus replication in each portion of the reproductive tract and the affected cell types. Although virus was detected in oral swabs collected from euthanized hens as early as 24 hours PI in group 1 (PA/H5N2) and continually detected in the other sampled birds at 36, 48, 60, and 72 hours PI, only 2 birds died from infection in this group, with death occurring only at 72 hours PI. The virus used in this group was originally isolated from an outbreak of HPAIV in layers in Pennsylvania, 2 and the mortality rates per day for this strain were lower when compared with the other strains involved in the same outbreak. 2 Groups 2 (Vn/H5N1) and 3 (Egy/H5N1) had earlier mortality, observed at 36 to 48 hours PI, but virus was detected in oral swabs as early as 24 hours PI. All the sampled birds had high levels of viral shedding detected by oral swabs collected at the time of death or following euthanasia with similar average titers (PA/H5N2 = 4.8 EID50/ml, Vn/H5N1 = 4.6 EID50/ml, and Egy/H5N1 = 3.5 EID50/ml), although chickens from the PA/H5N2 group shed higher titers at 48 hours PI, whereas the other 2 groups had high titers at 36 hours PI.
Score of Histologic Lesions and Immunohistochemistry Staining for HPAIV in Tissues of Hens Inoculated With A/Chicken/Pennsylvania/1370/83 (Group 1), A/Chicken/Vietnam/NCVD-675/2011 (Group 2), A/Chicken/Egypt/1063/2010 (Group 3), and Examined Over 72 Hours.

The results in this table represent the higher score of lesions for each time point collected. Histologic lesions are classified by intensity as follows: –, no lesions; +, mild lesions; ++, moderate lesions; and +++, severe lesions. The immunostaining was scored as follows: –, no staining; +, small clusters of positive cells; ++, multiple accumulations of positive cells; and +++, extensive staining. HE, hematoxylin and eosin; HPAIV, high-pathogenicity avian influenza virus; IHC, immunohistochemistry; NA, not applicable (no birds alive).
Gross lesions were observed at 36 hours PI in birds from groups 2 and 3; each group had 1 hen that was dead and another that was euthanized, but all 4 birds lacked prior clinical signs. One of the group 1 hens euthanized at 48 hours PI exhibited mild listlessness, but both euthanized hens had gross lesions, characterized by marked hyperemia all through the reproductive tract and hemorrhages in the follicle membranes. Gross changes were more severe in affected chickens from groups 2 and 3, consisting of extensive areas of hemorrhage in the ovary and less often in the brain and on the epicardium. Histologic changes in the reproductive tract were first observed at 36 hours PI in chickens from groups 2 and 3 and at 48 hours PI in chickens from group 1. Overall, lesions in the ovary consisted of perifollicular accumulations of heterophils and nuclear debris, with follicular disruption, hemorrhage, and consequent leakage of yolk material into the adjacent tissues and coelomic cavity (Fig. 1). The oviduct (infundibulum, magnum, and isthmus) contained areas of heterophil accumulation with fewer lymphocytes and plasma cells admixed with foci of hemorrhage throughout the edematous lamina propria. The superficial epithelial lining and less often the glandular epithelium had scattered areas of necrosis with multifocal heterophilic exocytosis (Figs. 2 and 3). Depending on the severity of these lesions, hemorrhage and cell debris were also observed within the lumen (Fig. 2). The most common change in the shell gland was variable degrees of edema in the lamina propria, with separation of endometrial glands and occasional accumulations of heterophils, lymphocytes, and plasma cells. Microscopic changes in the vagina and cloaca varied from small foci of heterophils to extensive areas of hemorrhage and accumulations of cell debris in the lamina propria and muscle layers.
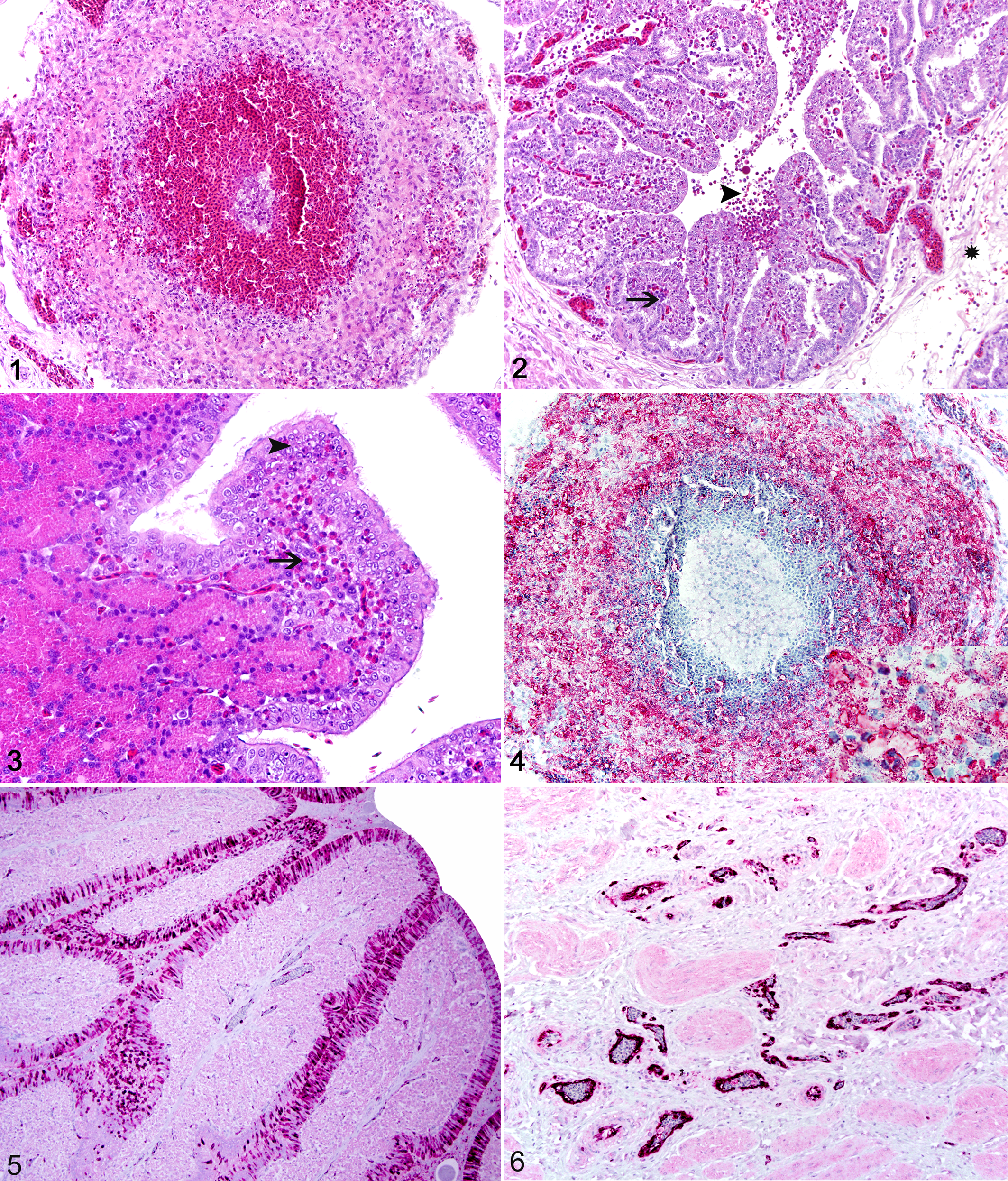
Positive immunostaining for HPAIV coincided with initial histologic changes. Finely granular, strong cytoplasmic, and/or nuclear immunostaining was detected in the ovarian stromal cells and superficial germinal epithelial cells, as well as in the mucosal epithelial and vascular endothelial cells along the infundibulum, magnum, isthmus, shell gland, vagina, and cloaca (Figs. 4 and 5). Glandular epithelial cells in the oviduct had scattered cytoplasmic staining. In addition, moderate to strong cytoplasmic immunostaining was detected in heterophils and macrophages in the ovary (Fig. 4) and lamina propria, as well as in the mucosal-associated lymphoid tissues throughout the reproductive tract. Vascular endothelial staining in the mucosa and muscle layers of the oviduct and lower reproductive tract was particularly strong in affected chickens in group 2 (Vn/H5N1) compared with the other groups (Fig. 6).
The nasal cavity, brain, heart, and spleen had histologic lesions and immunostaining similar to those previously reported for HPAIV infection in chickens 1,6,7 and were consistent with systemic viral infection.
A previous study was performed comparing susceptibility and distribution of H5N1 HPAIV in chickens, quail, and ducks; however, oviduct samples were not tested for antigen staining for chickens or ducks. In quail, 78% of the oviduct samples were positive for virus staining, but no details were given as to the cell types infected and which section of the reproductive tract was affected. 1 All hens analyzed in this study after 36 hours PI for groups 2 and 3 and after 48 hours PI for group 1 presented variable levels of positive virus staining in several portions of the oviduct.
HPAIV has been known to induce a decrease in egg production and egg quality, 4,9 and the virus has been detected on the eggshell and also in internal components of the egg after intranasal experimental challenge 4,9 and in the reproductive tract after natural infection, 1,7 indicating that the virus replicates in the reproductive tract. The presence of virus alone on the surface of the egg could be due to the high replication titers in the digestive tract and contamination of the egg as it passes from the reproductive tract into the cloaca, which is shared with the digestive tract. Reports of HPAIV in albumin and yolk and also in the oviduct suggest the virus also replicates in the reproductive tract and can infect the internal contents of the egg. However, very little has been known about the dynamics of the process. The present study showed differences in pathobiology for infections with different HPAIV strains, the time to develop lesions, and also the site of preference for virus replication, with particularly strong staining in the vascular endothelia in the mucosa and muscle layer of the oviduct and also lower reproductive tract in the affected chickens in group 2 (Vn/H5N1), compared with the other groups (PA/H5N2 and Egy/H5N1), where the staining was predominantly detected in the glandular epithelial cells along the oviduct. The virus replication in the glandular epithelia of the first portions of the reproductive tract (specifically ovaries, infundibulum, magnum, and isthmus) supports the already reported presence of virus in the yolk and albumin of eggs from infected hens, and the detection of virus in the lower part (ie, shell gland and cloaca) is probably responsible for the majority of the virus detected on the eggshell surface and could be augmented by intestinal contamination into the cloaca. The present study generated important data indicating HPAIV replicates in epithelial, inflammatory, and endothelial cells in the reproductive tract of hens, and this explains the presence of virus in the yolk and albumin of eggs laid by infected hens.
Footnotes
Acknowledgements
We thank Kira Moresco and James Doster for excellent technical support.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received the following financial support for the research, authorship, and/or publication of this article: This research was funded by Agricultural Research Service, Cooperative Research Information System (ARIS) project 6612-32000-063-00D.