
Abstract
Cattle persistently infected with a noncytopathic Bovine viral diarrhea virus (BVDV) are at risk of developing fatal “mucosal disease” (MD). The authors investigated the role of various apoptosis pathways in the pathogenesis of lesions in animals suffering from MD. Therefore, they compared the expression of caspase-3, caspase-8, caspase-9, and Bcl-2L1 (Bcl-x) in tissues of 6 BVDV-free control animals, 7 persistently infected (PI) animals that showed no signs of MD (non-MD PI animals), and 11 animals with MD and correlated the staining with the localization of mucosal lesions. Caspase-3 and -9 staining were markedly stronger in MD cases and were associated with mucosal lesions, even though non-MD PI animals and negative controls also expressed caspase-9. Conversely, caspase-8 was not elevated in any of the animals analyzed. Interestingly, Bcl-x also colocalized with mucosal lesions in the MD cases. However, Bcl-x was similarly expressed in tissues from all 3 groups, and thus, its role in apoptosis needs to be clarified. This study clearly illustrates ex vivo that the activation of the intrinsic, but not the extrinsic, apoptosis pathway is a key element in the pathogenesis of MD lesions observed in cattle persistently infected with BVDV. However, whether direct induction of apoptosis in infected cells or indirect effects induced by the virus are responsible for the lesions observed remains to be established.
Bovine viral diarrhea virus (BVDV) is an economically important animal pathogen that belongs to the genus Pestivirus of the family Flaviviridae. 46 This virus exists as 2 biotypes, cytopathic (cp) and noncytopathic (ncp), according to the effect on cultured cells.11,24,35 In pregnant cattle, ncp BVDV is able to cross the placenta, invade the fetus, and cause persistent infection. After birth, persistently infected (PI) calves may develop fatal mucosal disease (MD). From animals succumbing to MD, both an ncp and a cp biotype can be isolated4,7 that are antigenically very similar.6,8,18 The cp BVDV evolves by mutation from the ncp biotype within PI animals (for reviews, see Lindenbach et al, 26 Peterhans et al, 32 and Rümenapf and Thiel 34 ).
Apoptosis is a tightly regulated physiologic and pathologic process of cell death that may be elicited by several molecular pathways. The most important ones are the extrinsic and intrinsic pathways. In the extrinsic pathway (also known as the “death receptor pathway”), apoptosis is triggered by the ligand-induced activation of death receptors (eg, Fas receptor, tumor necrosis factor [TNF] receptor) at the cell surface, which leads to the activation of the initiator caspase-8. In the intrinsic pathway (also called the “mitochondrial pathway”), apoptosis results from an intracellular cascade of events leading to the release of mitochondrial proteins followed by caspase-9 activation. Both the intrinsic and the extrinsic pathways finally converge in the activation of the executioners caspase-3 and caspase-7.21,39 Among all these processes, antiapoptotic members of the Bcl-2 family, such as Bcl-2, Bcl-xL, and Bcl-w, can provide protection against apoptosis that is mediated through the mitochondrial pathway.22,45,49 This intrinsic pathway is suggested to be the most prominent cell death pathway in vertebrates, 21 and numerous viruses are known to cause apoptosis of infected cells. However, apoptosis might have opposing effects on viral pathogenesis by either preventing viral dissemination due to death of infected cells or by enhancing virus spread.27,33
It has been previously reported that cell death associated with cp BVDV in vitro is mediated by apoptosis.15,17,36,50 Infection of cell cultures with BVDV of the cp biotype is characterized by the predominant expression of the NS3 protein, the cleaved form of the nonstructural protein NS2-3. In contrast, in cells infected with ncp BVD viruses, the uncleaved NS2-3 precursor protein prevails. 23 It has been proposed that the NS3 protein of BVDV induces apoptosis that is correlated to caspase-8 and caspase-9 activation. 37 Madin-Darby bovine kidney (MDBK) cells infected with cp BVDV showed an increase in caspase-3 and caspase-9 activity compared with cells infected with ncp BVDV. The level of caspase-3 and caspase-9 activity was dependent on the proteolytic activity of NS3/4A. On the other hand, caspase-8 levels increased only slightly. 10 Furthermore, it has been demonstrated that macrophages infected with cp BVDV release a factor that promotes lipopolysaccharide (LPS)–induced apoptosis in uninfected macrophages by an interferon-independent pathway.20,31 Similarly, it has been shown that TNF-α is released by primary bovine fetal muscle cells experimentally infected with cp BVDV, and this might be involved in caspase-8 activation, albeit the effect was rather small compared with the strong stimulation of caspase-3 activity. 48 In other studies, oxidative stress, 36 endoplasmic reticulum (ER) stress,19,27 intracellular viral RNA 42 or double-stranded (ds) RNA accumulation, 47 and mitochondria-dependent caspase-9 activation 13 have been reported to be associated with cp BVDV-induced apoptosis.
Recently, in vitro infection of a lymphoid cell line (BL3) indicates that not only the biotype but also the virulence of a given virus strain plays an important role in apoptosis induction. Thus, in contrast to infection with nonvirulent ncp BVDV, infection with a virulent ncp BVDV-2 induced disruption of the mitochondrial transmembrane potential similar to cp BVDV. However, the effect was less pronounced and delayed, and a weaker activation of caspases was observed as compared with infection with a cp BVD type 2 virus. 3
Thus, the majority of the in vitro studies point to an involvement of the intrinsic rather than the extrinsic apoptosis pathway in cell death caused by cp BVDV infection.14,37,48 However, it is still unclear if this mechanism also participates in the development of lesions present in animals dying of MD and whether it is correlated with BVDV antigen expression in vivo. The aim of this retrospective study was, therefore, to investigate the role of the various apoptosis pathways in the pathogenesis of lesions in animals suffering from MD. For this purpose, we assessed and compared the expression of caspase-3, caspase-8, caspase-9, and Bcl-2L1 (Bcl-x) in tissues from BVDV-free control animals, from persistently infected (PI) animals that showed no signs of MD (non-MD PI animals), and in tissues from animals with MD as determined by the pathological findings.
Materials and Methods
Animals
In this retrospective study, a total of 24 cattle, mostly female and of the Brown Swiss breed, were included. These animals were submitted to necropsy between July 2000 and July 2007 to the Institute of Veterinary Pathology, Vetsuisse Faculty, University of Zurich, with the diagnosis of BVDV infection. The cases were classified according to macroscopic changes, histological lesions, and immunohistochemical findings as non-MD PI (n = 7) or as MD cases (n = 11) (see online supplementary material at http://vet.sagepub.com/supplemental). In addition, virus isolation was done retrospectively, and only animals from which an ncp and a cp biotype of BVDV could be isolated were analyzed as MD cases (Table 1). The non-MD cases were classified according to immunohistochemical staining for viral antigen in the skin irrespective of virus isolation, as with this method, only persistently infected animals were indentified.5,16,28 Sample tissues from 6 immunohistochemically BVDV-negative animals were used as controls.
Identification of Bovine Viral Diarrhea Virus Subgroups and Biotypes.
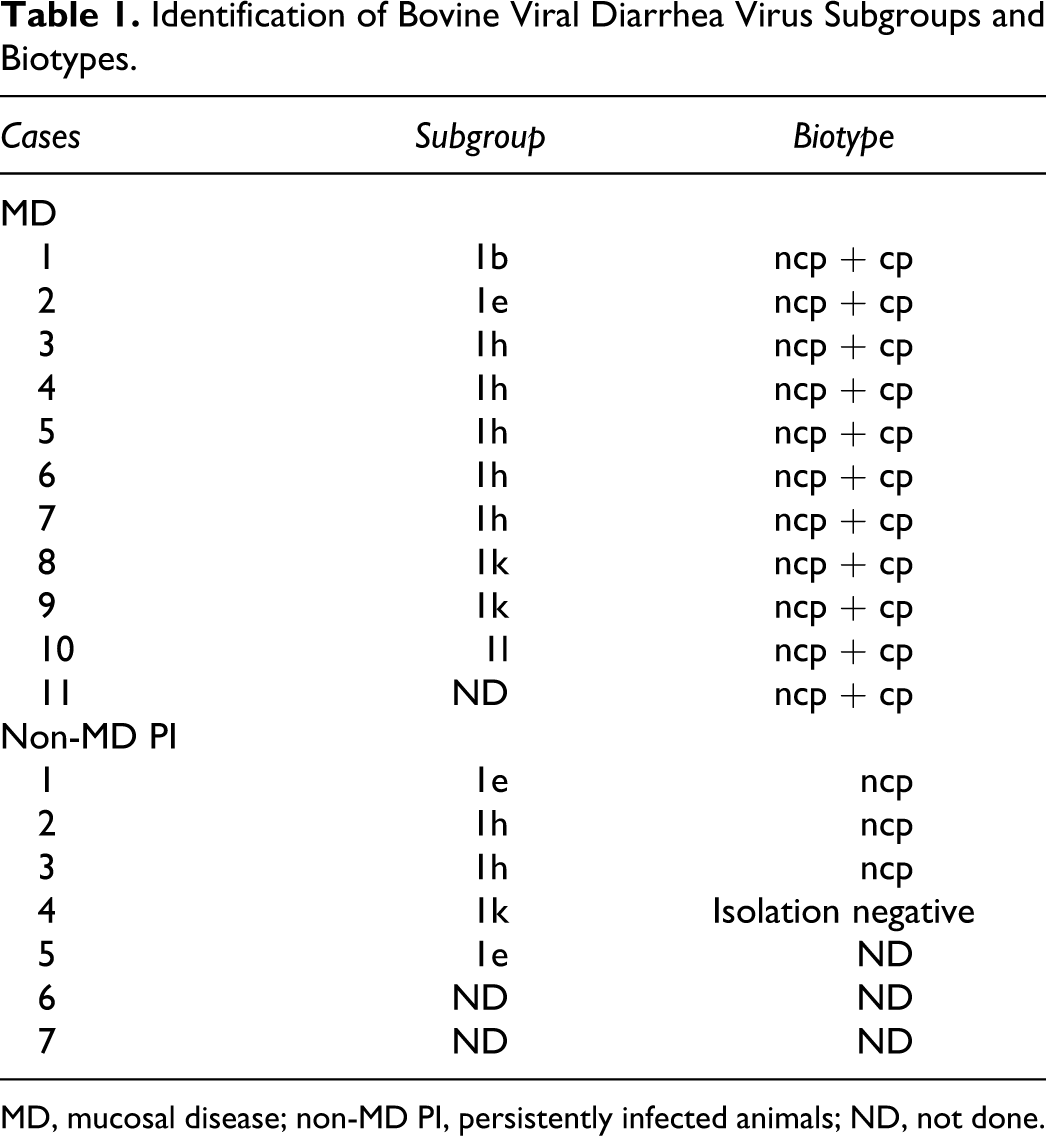
MD, mucosal disease; non-MD PI, persistently infected animals; ND, not done.
Immunohistochemistry
Immunohistochemical investigations were performed on paraffin-embedded sections from the following tissues: tongue, gingiva, esophagus, rumen, skin, lung, lymph node, spleen, and ileum. Sections of 2 to 3 μm were mounted on positive-charged glass slides, deparaffinized, rehydrated through graded alcohol, and washed in water. Slides for active caspase-3, caspase-9, and Bcl-2L1 (human Bcl-x) staining were twice pretreated by microwave cooking in a citric acid buffer (pH 6, DAKO, Code No. S2031; DAKO, Zug, Switzerland) for 10 minutes at 750 W and then counterstained with hemalaun for 2 minutes. Slides for caspase-8 and 15c5 staining were first counterstained with hemalaun and then pretreated with proteinase K (DAKO, Code No. S2019) for 10 minutes at room temperature (RT). After the corresponding pretreatment of the slides, endogenous peroxidases were inhibited by 3% H2O2 with 0.2% NaN3 in water. Subsequently, a protein blocking step (DAKO, Protein Block Serum-Free, Ready-to-Use, Code No. X0909) was performed. Both steps were carried out for 10 minutes at RT. Between the steps, the slides were carefully washed with phosphate-buffered saline (PBS; pH 8).
After the pretreatment and blocking steps, the slides were incubated with the following antibodies at different dilutions overnight at RT: active caspase-3, dilution 1:200 (rabbit anti–active capase-3, Code No. ab13847; Abcam, Cambridge, UK); Bcl-2L1, dilution 1:500 (rabbit anti-BCL2L1 [human Bcl-x], Code No. ab45002; Abcam); 15c5, dilution 1:10 000 (mouse anti-Pestivirus gp48/Erns; Dr. Dubovi, Cornell University, New York, New York); caspase-8, prediluted (rabbit anti–capase-8, Code No. ab15552; Abcam; this antibody is reported by the manufacturer to recognize both the full-length caspase-8 and the processed forms); and caspase-9, prediluted (rabbit anti–capase-9, Code No. ab13847; Abcam; as described by the manufacturer, the immunogen used corresponds to the N-terminal amino acids 1–134 of human caspase-9). As secondary antibody, anti-rabbit EnVision (DAKO, Code No. K4003) or anti-mouse EnVision (DAKO, Code No. K4001) was added for 30 minutes at RT. Subsequently, all slides were treated with AEC (3-amino-9-ethylcarbazole Substrate Kit, Code No. 00-2007; Invitrogen, Basel, Switzerland) for 10 to 15 minutes at RT. After controlling the color reaction and washing, the slides were mounted with an aqueous mounting solution (Kaiser’s gelatin glycerin, HX631620; Fluka, Buchs, Switzerland).
For each of the immunohistochemical assays, a positive and a negative control were included. The following organs that displayed the highest reactivities were used as a positive control: bovine thymus for active capase-3, bovine thymus and lymph node for Bcl-2L1 (human Bcl-x), bovine lymph node for caspase-8, bovine thymus for caspase-9, and brain tissue from a PI animal for 15c5 staining. As negative controls, the same organs without primary antibody but incubated with PBS were used.
Biotype Analysis
Organ samples of the thyroid gland or, when not available, the skin, tongue, or abomasum that had been stored at −20°C were used to determine the biotype of BVDV present in these samples. Approximately 30 mg of frozen tissue was homogenized in 1.8 ml cell culture medium (Earle’s minimal essential medium [MEM]; Seromed [Biochrom] Munich, Germany) enriched with 2% fetal bovine serum (FBS; Sigma or Oxoid GmbH, Wesel, Germany) using the Qiagen Tissue Lyser (Qiagen AG, Hombrechtikon, Switzerland). The homogenate was cleared by centrifugation at 13 000 × g prior to diluting it 7 times in 10-fold steps in MEM. FBS was free of BVDV and antibody to BVDV as tested by virus isolation and the serum neutralization test, respectively. Each dilution step was distributed to 6 wells (100 μl per well) of a 96-well microtiter plate, seeded with primary bovine turbinate cells prepared as described. 36 After 5 days of incubation at 37°C and 5% CO2, 20 μl of supernatant was transferred to a fresh 96-well microtiter plate, preseeded with bovine turbinate cells. After addition of 80 μl of fresh MEM, the microtiter plate was incubated as before, and the passaging procedure of the supernatant was repeated once more. After each passage, the cells were fixed and stained for viral protein by immunoperoxidase staining using a polyclonal swine-α-BVDV antibody produced at the Institute of Veterinary Virology, University of Bern. In addition, the cells were microscopically examined for the presence of a cytopathic effect.
BVD Virus Biotype, Genotype, and Subgroup Determination
Viral RNA was isolated from frozen organ samples as used for the biotype analysis. Prior to RNA isolation, approximately 30 mg of frozen tissue was homogenized in RLT buffer (Qiagen) using the Qiagen Tissue Lyser, and the homogenates were cleared by centrifugation. The RNeasy mini and viral RNA kits from Qiagen were used following the manufacturer’s instructions for RNA isolations from tissues and sera, respectively. All reverse transcriptase polymerase chain (RT-PCR) reactions were done as 1-step and single-tube reactions using the OneStep RT-PCR kit (Qiagen). For amplification of the 5′UTR, the pan-pesti primer pair 324/326 was used, 43 yielding a DNA fragment of 288 nucleotides based on the reference strain NADL. PCR products to be sequenced were separated by 1% agarose gel electrophoresis, and DNA fragments of the correct size were isolated using the QIAquick gel extraction kit (Qiagen). After spectrophotometrical quantification of the DNA at 260 nm, the required amount of DNA, primer, and water (aqua ad injectabilia; Dr G. Bichsel AG, Interlaken, Switzerland) was mixed and sent to Microsynth GmbH (Balgach, Switzerland) for sequencing. The economy run and premixed primer options were used. The sequencing data were assembled and aligned using the SeqMan (DNASTAR, Inc, Madison, Wisconsin) and Clone Manager (Scientific & Educational Software, Cary, North Carolina) software, respectively. Phylogenetic analysis was performed using the programs included in the GCG software package as PILEUP for the multiple sequence alignment and PAUP for calculating the distances by the Kimura 2-parameter method. To assess the BVDV genotype and subgroup, phylogenetic trees were constructed according to the neighbor-joining method, including 1000 bootstrap replicates. The genetic classification (Table 1) was performed for BVDV genotype 1 as described. 44
Data Analysis
The distribution of morphologic tissue lesions was compared blindly (ie, not knowing the BVDV status of the sample) with the immunohistochemical detection of active caspase-3, caspase-9, caspase-8, Bcl-x (Bcl-2L1), and the BVD viral protein Erns. The intensity of the staining was analyzed by optic microscopy, the number of positive cells were counted in 10 random fields (magnification: 20×), and the average of the 10 high-power fields were scored accordingly to the following scale: (–) = no immunohistochemical staining; (+) = single positive cells; + = slight positive staining, up to 15% positive cells; ++ = moderate positive staining, from 15% to 30% positive cells; and +++ = strong positive staining with more than 30% positive cells (Table 2).
Immunohistochemical Analysis.
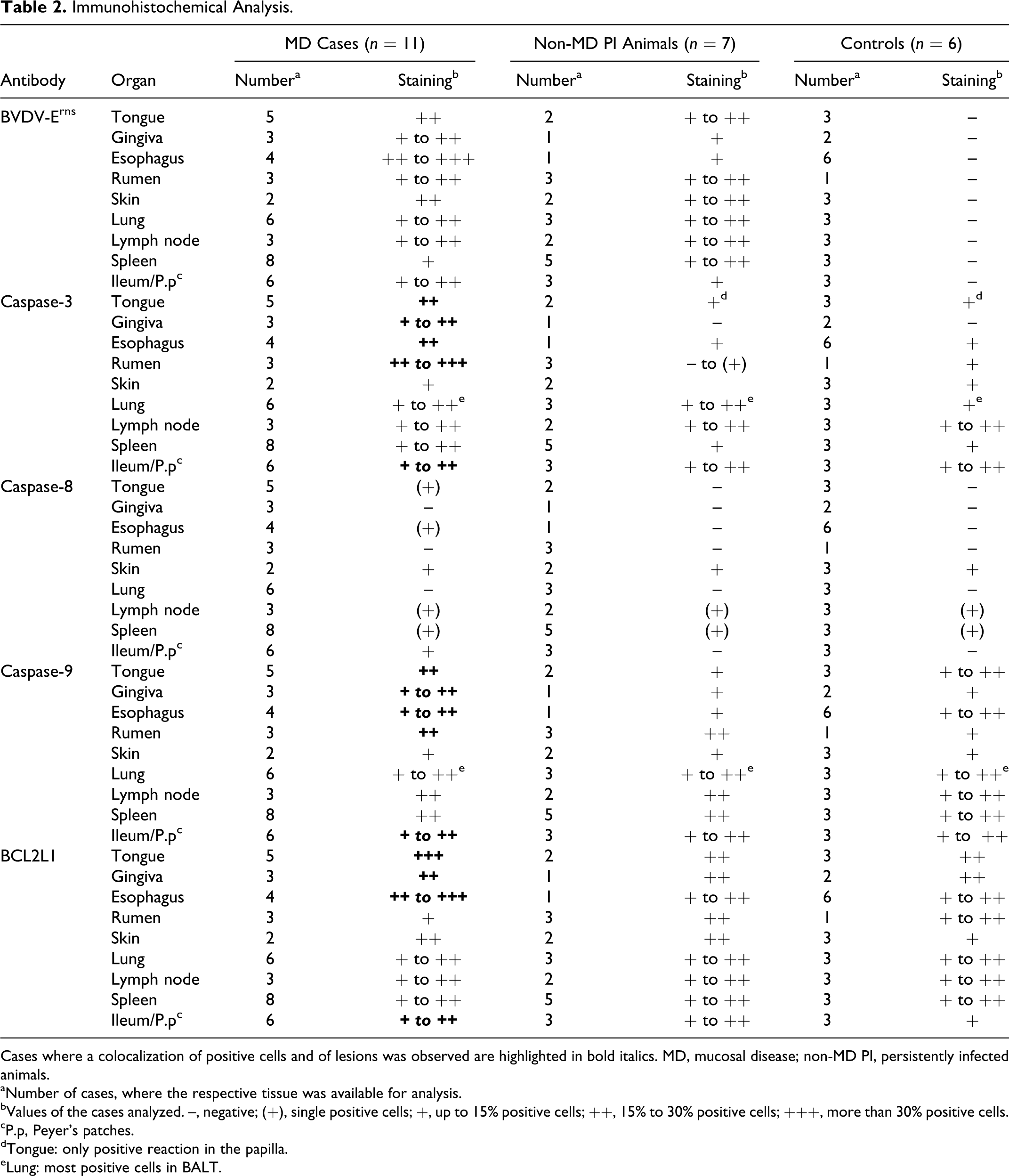
Cases where a colocalization of positive cells and of lesions was observed are highlighted in bold italics. MD, mucosal disease; non-MD PI, persistently infected animals.
aNumber of cases, where the respective tissue was available for analysis.
bValues of the cases analyzed. –, negative; (+), single positive cells; +, up to 15% positive cells; ++, 15% to 30% positive cells; +++, more than 30% positive cells.
cP.p, Peyer’s patches.
dTongue: only positive reaction in the papilla.
eLung: most positive cells in BALT.
Results
BVDV Antigen Expression
Cells that were positively labeled with BVDV/MD immunohistochemistry were mostly epithelial cells, including basal cells as well as hair root sheath epithelial cells in the skin, and most layers of the squamous epithelium of the tongue, esophagus, and rumen. In the ileum, only a few epithelial cells in the crypts were positive. Also positive were fibrocytes, endothelial and medial cells of blood vessels, a few lymphocytes in various lymphatic tissues, and occasionally lung macrophages. In the brain that was used as a positive control, neurons, endothelial cells, and some glial cells were positive.
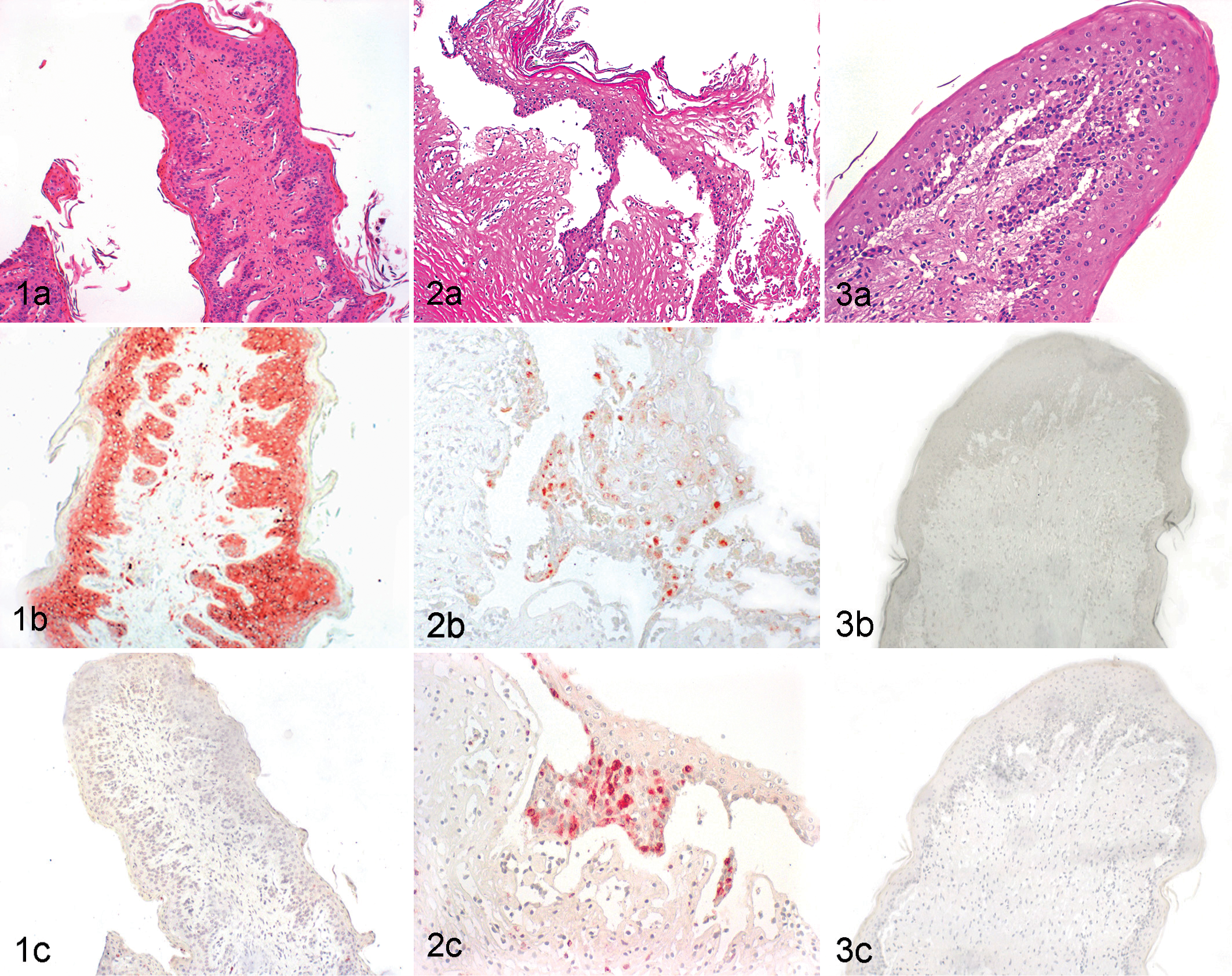
All MD and non-MD PI cases displayed positive immunohistochemical staining with the 15c5c antibody directed against the viral glycoprotein Erns. Most layers of the epithelial cells in non-MD PI cases showed a strong intracytoplasmic labeling with the 15c5 antibody. No difference was visible between the staining intensity in tissue without lesions in both MD and non-MD PI groups. In areas of mucosal lesions, the staining was diminished due to loss and degeneration of epithelial cells. Control animals were negative with the same antibody (Figs. 1–3, panels b; Table 2).
Expression of Active Caspase-3
All animals with MD displayed a positive intracytoplasmic staining for active caspase-3 in all tissues analyzed (Fig. 2c; Table 2). In tongue, gingiva, esophagus, rumen, and ileum, the positive caspase-3 staining was correlated to areas of lesions, such as erosions or ulcerations in the tongue and depletion of Peyer’s patches. Positive staining was mainly seen in epithelial cells of the basal layer in and adjacent to the eroded and ulcerated areas. In addition, positive staining was observed in neutrophils, fibrocytes, and crypt epithelial cells in the ileum and in lymphocytes of the Peyer’s patches.
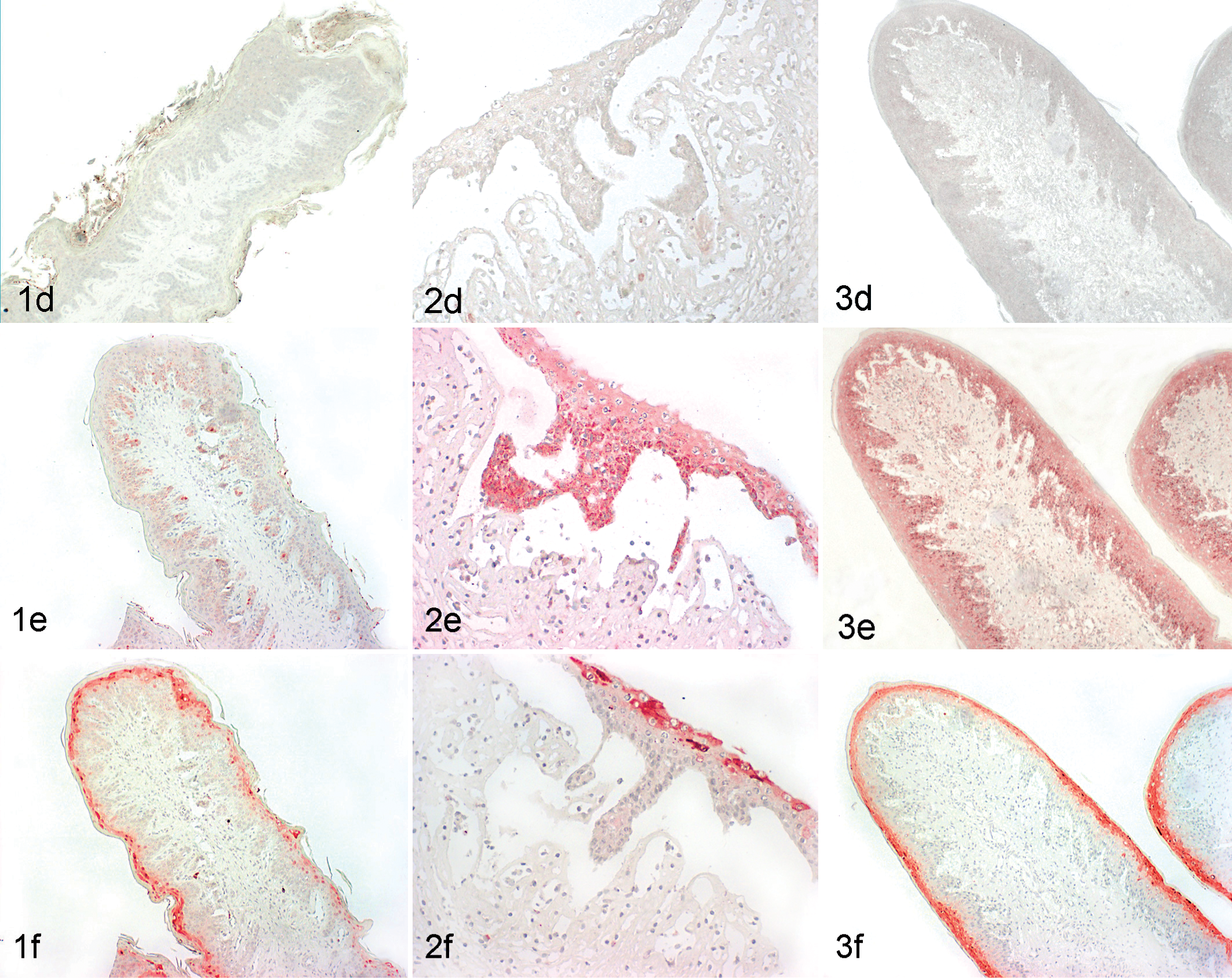
In contrast to the MD cases, the non-MD PI animals showed no or very discrete and localized caspase-3 staining in tongue (mostly tongue papillae), gingiva, esophagus, and rumen. In lymphoid tissues (bronchus-associated lymphoid tissue [BALT], lymph node, spleen, ileum), however, the expression of caspase-3 was similar as compared with the MD cases. In the skin, sebaceous glands and hair follicles showed moderate positive staining in all groups (non-MD, MD, and controls).
The negative control animals showed only slight caspase-3 staining in mucosal tissues, and only a few positive cells were detected in lymphoid tissues such as spleen, lymph node, and ileum.
Expression of Caspase-8
The expression of this protein was mainly detected in sebaceous glands and hair follicles of the skin of all 3 groups of animals. In the Peyer’s patches, only a few positive lymphocytes were visible in the MD cases. All other organs were negative or very weakly positive (eg, single positive epithelial cells) (Figs. 1–3, panels d; Table 2).
Expression of Caspase-9
The expression pattern of caspase-9 in MD cases was very similar to the results found with the active capase-3 antibody (Table 2). All samples analyzed from animals with MD displayed a positive staining in tongue, gingiva, esophagus, rumen, and ileum. In addition, caspase-9 positive labeling was correlated to areas of erosion and ulceration in these organs and to the depletion of Peyer’s patches (Fig. 2e). In lymph nodes and spleen, a few lymphocytes in the follicle area and in the lung in the BALT region were positive.
Non-MD PI animals showed discrete staining in tongue, gingiva, ruminal epithelium (Fig. 1e), and esophagus. The expression of caspase-9 in lymphoid tissues and lung was similar to the staining in the MD cases.
Negative control animals displayed a weak to moderate positive labeling in the epithelium of the digestive tract organs analyzed and in organs such as lymph node, spleen, and lung. The staining was similar or even stronger than seen in the non-MD PI animals (Fig. 3e; Table 2). Thus, it was not possible to distinguish the intensity of expression of caspase-9 between non-MD, MD, and control groups in the lymph node, spleen, and lung. However, the intensity of staining of epithelial cells was stronger in the MD cases than in the non-MD and control animals and correlated with ulcerative or erosive lesions.
Expression of Bcl-x (Bcl-2L1)
In MD cases, the expression of Bcl-x was detected in areas of erosions and ulcerations of tongue, gingiva, and esophagus and in areas of depletion of the Peyer’s patches (Table 2). The cells showing strong positive staining were primarily epithelial cells from the upper layers of the stratified squamous epithelium, and weaker staining was visible in the basal cell layers (Fig. 2f). In addition, some inflammatory cells such as neutrophils and lymphocytes of the Peyer’s patches were positive.
In non-MD PI animals, Bcl-x expression was also observed, although weaker and mainly in the upper epithelial cell layers but in the same tissues as described for the MD cases (Fig. 1f). Weak Bcl-x staining was also observed in other tissues such as in the follicles of the lymph node, in a few lymphocytes in the periarteriolar sheaths of the spleen, and in some lymphocytes of the BALT in the lung from all animals studied. Generally, the staining pattern was overall stronger in MD cases than in non-MD PI and BVDV-negative cases (Figs. 1–3, panels f; Table 2).
Biotype and Subgroup Analysis
BVDV of the cp biotype in addition to the persisting ncp strain could be isolated from 11 animals that were therefore classified as MD cases (Table 1). As expected, no cp biotype was isolated from the animals of the non-MD group.
The viruses present in the MD and non-MD PI animals belonged to the subgroups 1 h (7), 1 k (3), 1e (3), 1b (1), and 1 l (1). From 3 animals, the subgroup identification was not done (Table 1). The spectrum of subgroups was similar in the MD and the non-MD PI animals and reflects that of BVD-1 viruses described to circulate in Switzerland.1,38
Discussion
In this retrospective study, we investigated the apoptotic pathways, extrinsic or intrinsic, implicated in BVDV persistently infected animals, either suffering from MD or showing no macroscopical or histological signs thereof. The immunohistochemical staining used in this study is a highly specific and sensitive method to detect apoptotic cells in archival paraffin-embedded material. 12 Due to the limited availability of the study material, further analyses (eg, using Western blotting or real-time RT-PCR) could not be applied. However, this might not be disadvantageous as the latter methods are bulk cell measurements that are inherently less informative than immunohistochemistry.
Thus, by means of immunohistochemistry, MD cases showed more caspase-3 and slightly more caspase-9 expression than non-MD PI animals. Interestingly, the caspase-3 and -9 staining was correlated to erosive and ulcerative lesions in organs of the digestive tract (tongue, gingiva, esophagus, rumen, and ileum), whereas this was not the case in the other organs analyzed (skin, lung, lymph node, and spleen). This phenomenon might be correlated to the presence of bacteria or bacterial constituents such as LPS in the digestive tract. As previously shown, cp BVDV primes uninfected macrophages for LPS-induced apoptosis,20,31 which might also contribute to the cell death observed in the gastrointestinal tract. However, in contrast to caspase-3 staining that was markedly stronger in BVDV-infected animals, caspase-9 expression was also quite high in the negative control animals.
These results suggest that caspases-3 and -9 and, thus, the intrinsic apoptotic pathway are involved in the pathogenesis of the mucosal lesions typically observed in MD cases. This is corroborated by the fact that caspase-8 was not elevated in our study, leading to the conclusion that the extrinsic pathway is rather not involved. However, it remains to be clarified whether the intrinsic pathway is directly or indirectly induced by BVDV or rather by unspecific activation promoted by the tissue damage. In a recent study of fetal and neonatal small ruminants naturally infected with Border disease virus (BDV), neuropathological analysis of the central nervous system indicated that the intrinsic pathway of apoptosis was most important but that both the intrinsic and extrinsic pathways are involved in infection with a noncytopathic BDV. 41 Whether the participation of the extrinsic pathway is restricted to certain pestiviruses or to different cell types is, however, not yet known.
Recently, experimental acute infection of calves with an ncp biotype of BVDV-1 led to a progressive depletion of the Peyer’s patches. The depletion was due to massive lymphocyte apoptosis with concurrent activation of caspase-3 as analyzed by immunohistochemistry. 30 However, lymphocyte apoptosis in the gut-associated lymphoid tissue (GALT) was paralleled by an increase in caspase-8 and rather a decrease in caspase-9, indicating that the extrinsic rather than the intrinsic pathway of apoptosis might predominate in acute BVDV infections. 29
This is in contrast to the results presented in this study that showed that the intrinsic pathway is a key element in the induction of apoptosis in MD pathogenesis in persistently infected calves. Therefore, cell death induced by a cp biotype of BVDV in an immunotolerant animal seems to occur through different pathways than during acute infections with ncp BVD viruses. Moreover, changes in cell proliferation or immune-mediated cell damage induced by cp BVDV might also contribute to the mucosal atrophy observed in MD.9,25 Thus, it is rather probable that direct virus-induced cell death and indirect mechanisms contribute to the overall pathology observed in MD.
Interestingly, not only the expression of caspases-3 and -9 but also the protein Bcl-x colocalized with the mucosal lesions of the MD cases. It was shown in one study in vitro that induction of cellular apoptosis by a cp strain of BVDV-1 correlates with downregulation of the antiapoptotic Bcl-2 protein. However, at early time points postinfection, there was rather an increase in Bcl-2 expression. 19 In another study, the Bcl-2 protein was upregulated by ncp BVDV infection, presumably suppressing the activation of executioner caspases and the initiation of cell death, thus enabling persistent infection in vitro. 2 In our study, Bcl-x had slightly stronger expression in tissues from MD cases than in non-MD PI and noninfected control animals. Thus, it remains to be investigated whether Bcl-x, its splice variant Bcl-xS, 40 or other members of the Bcl-2 family have a role in inhibition of apoptosis or even in activation of cell death and whether they are directly modulated by BVDV or rather in an unspecific way.
The viral subgroups present in the samples analyzed encompass the complete spectrum of strains circulating in Switzerland. 1 There was no correlation of a certain genetic subgroup to a specific type of macroscopic or immunohistochemical finding. This makes strain-specific differences in apoptosis very unlikely, despite that only a small number of animals per subgroup could be analyzed.
In conclusion, markers of the intrinsic apoptotic pathway were activated in mucosal lesions of animals suffering from MD, which corresponds to the in vitro findings that activation of the initiator caspase-9 and the executioner caspase-3 might play a major role in cp BVDV-induced cell death. However, we were unable to demonstrate that cells with enhanced staining for activated caspase-3 were directly infected with cp BVDV. The fact that strong caspase-3 and -9 staining was predominantly detected in lesions of the gastrointestinal tract (ie, in regions rich in LPS or other bacterial constituents) might indicate that priming of uninfected cells by cp BVDV for activation-induced apoptosis also contributes to the formation of the lesions observed. Thus, further in vivo studies are required to unravel the complex pathogenesis of the clinical and necropsy findings observed in this lethal disease.
Footnotes
Acknowledgement
Thanks to the technical staff of the Institute of Veterinary Pathology for excellent technical work.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.