
Abstract
Migrating bats have increased mortality near moving turbine blades at wind farms. The authors evaluated competing hypotheses of barotrauma and traumatic injury to determine the cause. They first examined the utility of lungs from salvaged bat carcasses for histopathologic diagnosis of barotrauma and studied laboratory mice as a model system. Postmortem time, environmental temperature, and freezing of carcasses all affected the development of vascular congestion, hemorrhage, and edema. These common tissue artifacts mimicked the diagnostic criteria of pulmonary barotrauma; therefore, lung tissues from salvaged bats should not be used for barotrauma diagnosis. The authors next compared wind farm (WF) bats to building collision (BC) bats collected near downtown Chicago buildings. WF bats had an increased incidence in fracture cases and specific bone fractures and had more external lacerations than BC bats. WF bats had additional features of traumatic injury, including diaphragmatic hernia, subcutaneous hemorrhage, and bone marrow emboli. In summary, 73% (190 of 262) of WF bats had lesions consistent with traumatic injury. The authors then examined for ruptured tympana, a sensitive marker of barotrauma in humans. BC bats had only 1 case (2%, 1 of 42), but this was attributed to concurrent cranial fractures, whereas WF bats had a 20% (16 of 81) incidence. When cases with concurrent traumatic injury were excluded, this yielded a small fraction (6%, 5 of 81) of WF bats with lesions possibly consistent with barotrauma etiology. Forensic pathology examination of the data strongly suggests that traumatic injury is the major cause of bat mortality at wind farms and, at best, barotrauma is a minor etiology.
Bat mortality is increased near moving, but not static, turbine blades at wind farms (WFs). 2,9,14,21,23,25 The 2 leading hypotheses for the mortality include blunt force trauma from being struck by moving turbine blades and barotrauma. Barotrauma is a phenomenon in which abrupt air pressure changes cause tissue damage to air-containing structures (eg, lungs). It is suggested by some to cause a high proportion of bat deaths at WFs, with up to 90% of bats reported to have lung lesions consistent with barotrauma. 4 Key diagnostic features of pulmonary barotrauma include microscopic detection of hemorrhage and edema in airspaces with vascular congestion and interstitial bullae. 1,4,15,29 However, pulmonary barotrauma diagnosis has primarily been based on examination of lung tissues from salvaged bat carcasses. The time from death to carcass collection (and eventual tissue fixation) in these cases is not typically known, and in some cases only an estimated window of time (eg, died the previous night) is at best available. 3,4 Prior to collection, the carcasses may be exposed to various environmental conditions (eg, temperature, sunlight) that can alter tissue decomposition kinetics and tissue morphology. 35,42 Additionally, biologists frequently salvage bat carcasses from WFs for study and may freeze them for long-term storage, which could cause additional morphologic changes.
We designed a 2-part forensic investigation into the etiology of bat mortality at WFs. First, we hypothesized that postmortem decomposition and environmental conditions would influence the development of morphologic artifacts in the lung that could mimic and confound the diagnosis of pulmonary barotrauma. We examined a laboratory mouse model to longitudinally study the appearance of these morphologic parameters to determine the diagnostic utility of salvaged lung tissue for barotrauma diagnosis. Second, we designed a controlled experiment to evaluate bat carcasses for signs of traumatic injury and barotrauma. To this end, we examined salvaged bats collected from a central Illinois WF compared with a control group collected from downtown Chicago that had died following collision with a building.
Materials and Methods
Experiment 1: Study Animals and Treatments
All mouse trials were performed under laboratory conditions in the School of Biological Sciences at Illinois State University or the Roy J. and Lucille A. Carver College of Medicine at the University of Iowa. All tissue specimens were analyzed at the Comparative Pathology Laboratory at the University of Iowa. We chose to use a laboratory mouse model because of accessibility and previous use in barotrauma studies. 7,8,15 Given the difficulties of performing mortality studies in wild or captive bat populations, the mouse model is a useful option to study decomposition events that are not well described in bats.
The bat lung used in experiment 1 came from a central Illinois WF. The time of death was estimated to be the prior night, based on regular collection of carcasses and lack of external evidence for decomposition. 4
For the freezing and decomposition study, we euthanized retired female breeder mice (n = 65, ∽30 g of body weight) of the ICR outbred strain Hsd: ICR (CD-1) using 5% isoflurane delivered by 30% oxygen and 70% nitrogen for 15–20 minutes. For the temperature study, we euthanized retired female breeder mice (B6 J, n = 18, ∽30 g of body weight) by carbon dioxide intoxication followed by thoracotomy. While many euthanasia techniques are associated with minor lung artifacts such as uncommon congestion and hemorrhage, we believe that the longitudinal nature of the decomposition study allows for sufficient segregation of euthanasia versus postmortem effects. Carcasses were randomly situated for each trial to avoid consistent recumbency, which best mimics the varied presentations of bat carcasses found dead at WFs. All protocols complied with the National Institutes of Health’s Guide for the Care and Use of Laboratory Animals at Illinois State University and the University of Iowa.
Decomposition Trial
We studied mice longitudinally to determine the influence of decomposition on postmortem tissue changes. Immediately after euthanasia, we divided carcasses into treatment groups (5 per treatment): 1 control group (never frozen and necropsied immediately) and 8 experimental groups, where mice decomposed in individual sealed plastic bags (Ziploc sandwich bags) inside sealed Tupperware containers at controlled room temperature (∽22°C) for 1, 2, 4, 6, 24, 48, 72, and 96 hours.
Temperature Trial
We studied the influence of elevated temperature on postmortem tissue changes by placing each euthanized adult mouse (3 per treatment) into double-sealed plastic bags and incubating them at 33°C for 1, 2, 4, 8, 24, and 48 hours. The temperature was chosen for 2 reasons: (1) Evening temperatures can reach the low 30s (°C) during the first portion of the migration period of bats (ie, when bat deaths increase at WFs) in the Midwestern United States; (2) bat carcasses salvaged the following day may have variable exposure to heat-retaining rocks/soil, direct sunlight (ie, thermal radiation), and hot daytime temperatures prior to salvage.
Freezing Trial
We studied the effect of freezing on mouse lung morphology. Immediately after euthanasia, we subjected carcasses (5 per treatment) to 1 of 3 treatments before dissection:
Control: never frozen, immediately necropsied and fixed
Freeze trial: frozen at –20° C for 1 month, thawed, necropsied, and fixed
Freeze/thaw trial: frozen at –20°C for 1 month, thawed for 2 hours, refrozen for 1 hour, thawed, necropsied, and fixed
Tissue Handling and Scoring
Immediately following each trial, we removed lungs, heart, and trachea from each mouse and fixed the organs in 10% neutral buffered formalin (no insufflation of fixative into airways was performed) for routine processing and staining by the Comparative Pathology Laboratory (University of Iowa). Lungs were consistently embedded and sectioned so that left and right lobes were always examined. Sections of whole lung were examined (by DKM—blinded to treatment groups at examination) for signs of vascular congestion, erythrocyte extravasation (erythrocytes detected in airspaces), and edema (seroproteinaceous material in airspaces) (Figs. 1 –4). For each parameter, lung samples were scored using a scale of 0–3: 0, none detected; 1, detectable, rare (< 5% lung field); 2, detectable, moderate extent/distribution (5% to < 50% lung field); 3, widespread extent/distribution (50% to 100% lung field). For each trial, scores for each pathologic parameter were assessed with a one-way analysis of variance, followed by a Dunnett posttest to compare each group to the control (fresh lung immediately necropsied and fixed). Statistical significance was recognized at P < .05.
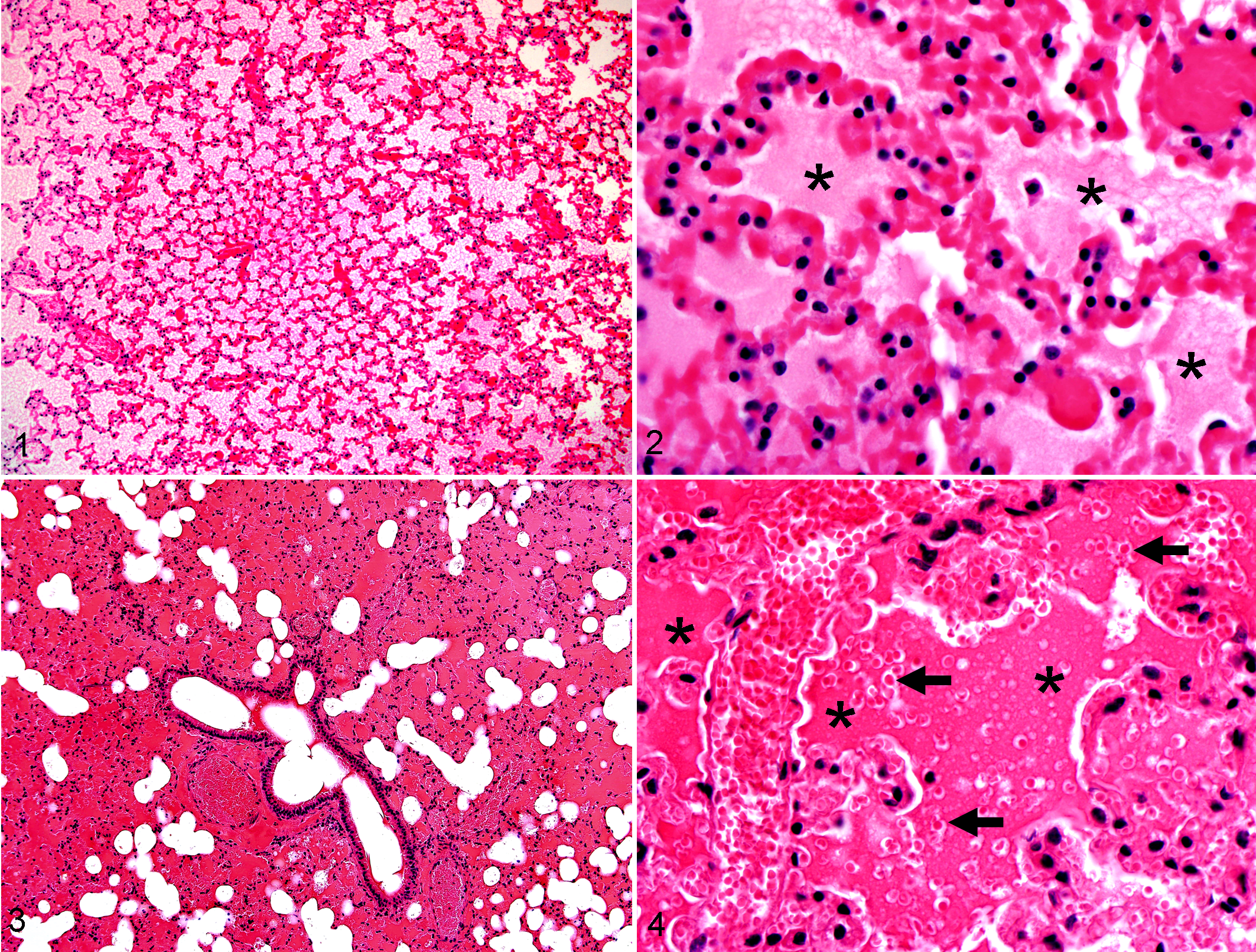
Lung, mouse; after 4 hours at 33°C. Example of diffuse filling of airspaces with eosinophilic material. HE stain.
Since hemorrhage is a common parameter for barotrauma diagnosis, we designed this scoring system to be sensitive for erythrocyte extravasation events (ie, solitary to loose aggregates of erythrocytes detectable in airspaces). The distribution of erythrocyte extravasation and other parameters was the primary means of scoring severity and adapted from previous scoring methods. 12,15,19 For example, an erythrocyte extravasation score of 3 meant that the detection of erythrocyte extravasation was widely distributed, albeit often without necessarily showing overt histologic hemorrhage (ie, complete filling of airspaces by erythrocytes). Additionally, we examined the lung for vascular congestion, as it can be frequently misdiagnosed by inexperienced observers as pulmonary hemorrhage and because it has been reported in bat pulmonary barotrauma. 4
Experiment 2
Animals
We examined bats salvaged near buildings in downtown Chicago, Illinois (building collision [BC] bats, n = 53), and compared these with bats salvaged from a central Illinois WF (Vestas V82 turbines, 1.65 MW) (WF bats, n = 262). BC bats served as controls for WF bats, as they died via collision with buildings 8 and lacked exposure to barotrauma. The BC bats were salvaged during 1907–2009 near buildings in downtown Chicago, Illinois (n = 53), deposited at the Field Museum of Natural History in Chicago and preserved in 70% ethanol; specimens included 37 eastern red bats (Lasiurus borealis), 14 silver-haired bats (Lasionycteris noctivagans), 1 big brown bat (Eptesicus fuscus), and 1 little brown bat (Myotis lucifugus). Mortality in this group was attributed to collision with buildings and, more important, could not have been due to exposure to moving turbine blades.
The WF bats were salvaged during 2008–2010 from a central Illinois WF (n = 262) and deposited at Illinois State University. They included 3 migratory tree-roosting bat species: 78 hoary (Lasiurus cinereus), 120 eastern red, and 64 silver-haired. Bats salvaged in 2008 and 2009 were frozen at –80°C prior to examination, while 2010 bats were examined immediately after salvage.
Trauma assessment
Both BC and WF bats were radiographically examined for bone lesions consistent with traumatic injury (Figs. 5 –8). Initially, whole body radiographs (IDEXX-DR 1417 Digital Radiography System) and skull radiographs (Dentsply Gendex 765 DC) were taken, and specific bones (Table 1) were examined for fractures and dislocations. In addition, 66 of the 262 WF bats were necropsied and examined for supplemental lesions consistent with trauma, including diaphragmatic hernia, self-bitten tongues, external lacerations, and subcutaneous hemorrhage. All BC bats were assessed for self-bitten tongues and external lacerations; however, these museum specimens were not available for full necropsies and assessment of subcutaneous hemorrhage or diaphragmatic hernia.
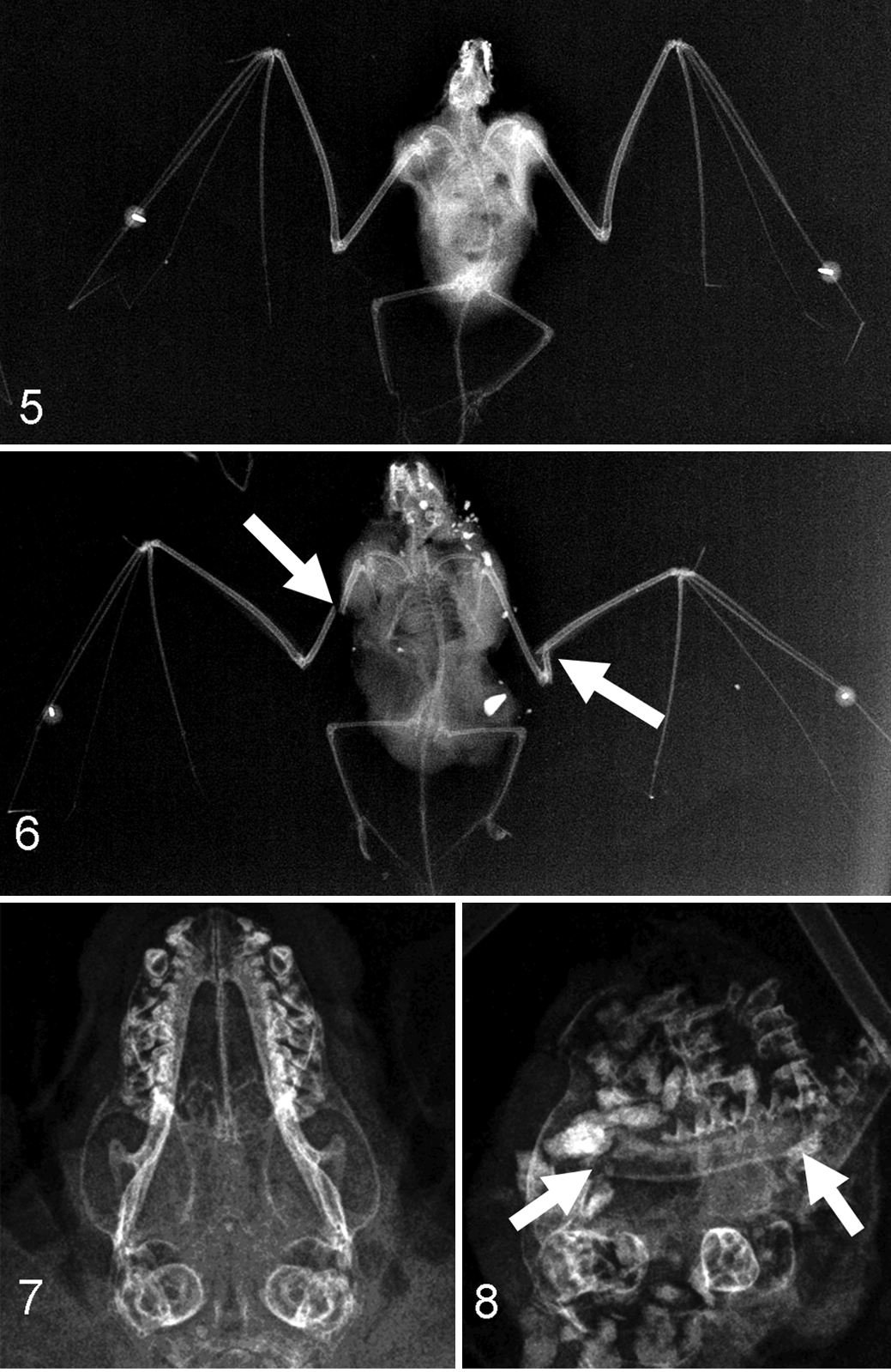
Radiograph of eastern red bat (Lasiurus borealis). Example of a bat carcass that lacked fractures.
Decomposition Study of Mice With Histologic Scoring of Lung Parameters at Different Time Points for Both Temperatures a

aMean ± SEM. n = 5 and 3 per group respectively. Statistical significance compared to fresh tissues using a Dunnett posttest.
*P < .05.
**P < .01.
***P < .001.
Similar to that described in previous work, 4 a subset of 14 WF bats were identified from the 2010 collection season that had died the night before, based on consistent daily carcass searches and appearance of the specimen (cf Materials and Methods, Experiment 2). 4 The lungs of these 14 recently killed bats were collected for histopathologic examination, fixed in 10% neutral buffered formalin, and sent to the Comparative Pathology Laboratory (University of Iowa) where tissues were routinely processed, stained (hematoxylin and eosin), and subsequently examined as in experiment 1.
Auditory barotrauma assessment
BC (n = 42) and WF (n = 81) bats that appeared relatively fresh (eg, absence of maggots) were examined for evidence of tympana rupture. Tympana were photographed using an AM-311H Dino-Lite Digital Ear Scope that was fitted with a custom-made plastic otoscope adaptor (2 mm) to view narrow ear canals of small bats. Specimens with rupture of 1 or both tympana were considered positive, whereas absence of rupture was considered negative.
Results
Experiment 1
Decomposition trial
In controlled postmortem environmental conditions (∽22°C), decomposition caused changes in vascular congestion, erythrocyte extravasation, and edema in the murine lung. Vascular congestion scores were significantly increased as soon as 2 hours postmortem (P < .01, Dunnett posttest), edema scores were increased after 24 hours (P < .05, Dunnett posttest) and increased erythrocyte extravasation scores were increased after 72 hours (P < .001, Dunnett posttest, Table 1).
Temperature trial
Elevated temperature (33°C) accelerated postmortem morphologic changes in the model. Increased edema was detected in airspaces as quickly as 2 hours postmortem (P < .001, Dunnett posttest) with increased erythrocyte extravasation detected by 4 hours postmortem (P < .01, Dunnett posttest, Table 1). Vascular congestion was present, but the scores were not statistically significant, likely due in part to the small number of animals per group (n = 3).
Freezing trial
The tissues from the freeze and freeze/thaw trials had increased erythrocyte extravasation (P < .001, Dunnett posttest) and edema (P < .001, Dunnett posttest) compared to fresh lung tissue but did not significantly differ from each other (Dunnett posttest) (Table 2). Vascular congestion was present but did not significantly differ among freezing treatments.
Freezing Study of Mice With Histopathologic Scoring of Lung Parameters a
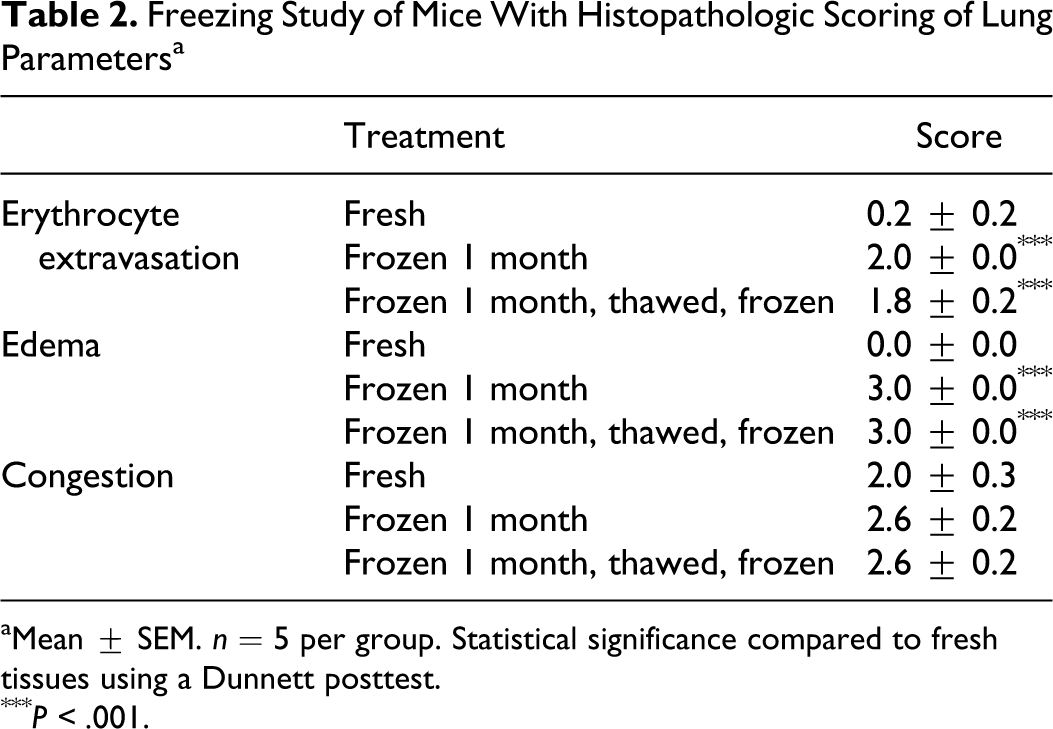
aMean ± SEM. n = 5 per group. Statistical significance compared to fresh tissues using a Dunnett posttest.
*** P < .001.
Morphology
While erythrocyte extravasation was often detected in the treatment groups as solitary erythrocytes to loose cellular aggregates, overt hemorrhage with alveolar filling by erythrocytes was uncommon and, if present, usually focal. In some of the samples, decomposition was associated with severe hemolysis, suggesting that progressive decomposition could confound the histopathologic detection of overt lung hemorrhage. In these trials, the morphologic filling of alveoli by edema or hemorrhage usually ranged from multifocal to diffuse. However, the lung morphology from the decomposition/temperature trials could usually be distinguished from the freezing trials by the appearance of a peripheral rim of pallor (pale staining) along the pleural border that appeared at 2 hours at 33°C and by 24 hours at 22°C (Figs. 9, 10), matching the chronologic appearance of edema (Table 1). This rim of pallor had minor variations in thickness and remained generally distinct in location even with advanced decomposition, in which it was eventually detected in all of the mice of the decomposition studies. These changes were not likely artifacts of histologic processing, as Comparative Pathology Laboratory procedures are designed to avoid possible “batch” effects that could mimic treatment-specific changes in research studies (ie, large treatment groups are divided into mixed batches during processing and staining). The lack of similar peripheral pallor in frozen samples processed at the same time would suggest that it is a decomposition specific change and not artifact. Interestingly, peripheral pallor was detectable in a portion of the lungs from “fresh” bats collected the morning following presumed death (Fig. 11).
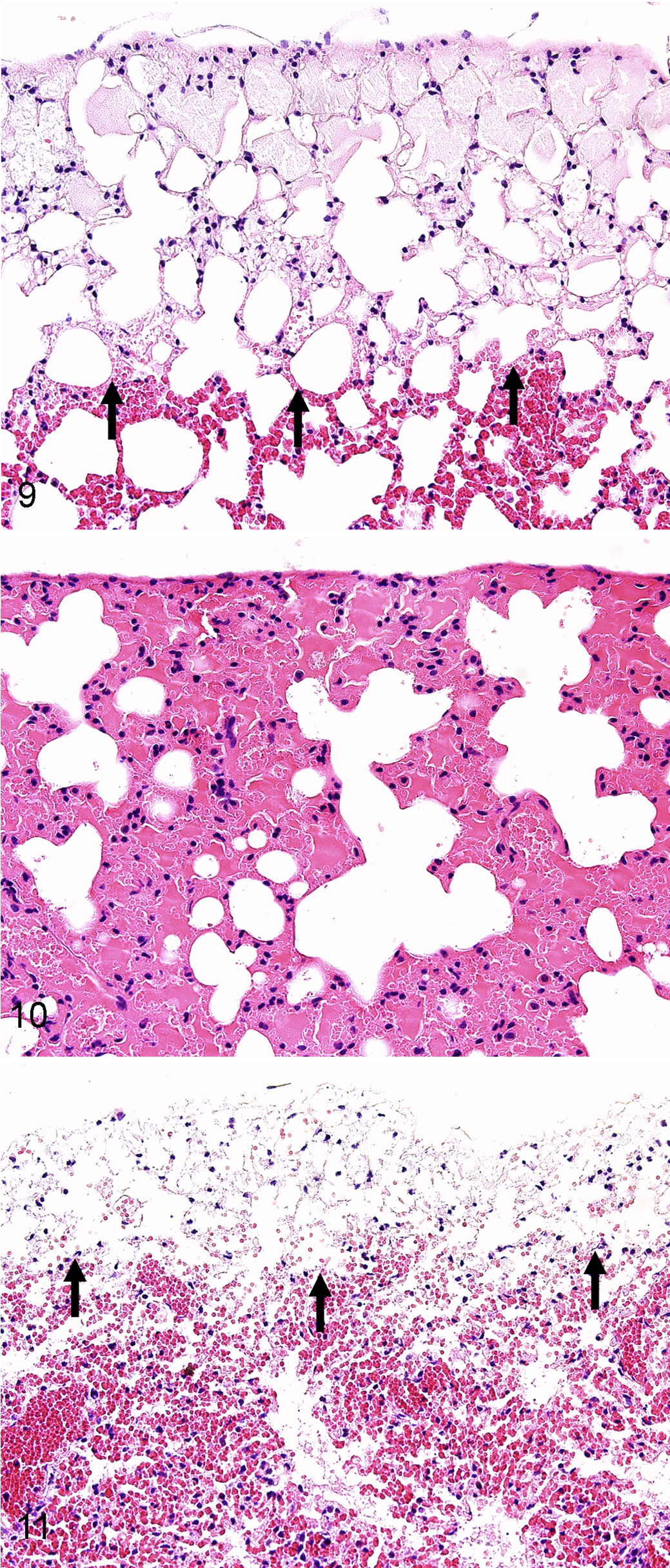
Lung, mouse; after 24 hours at 22°C. Note the rim of pallor (arrows) along the pleural border. HE stain.
Experiment 2
Radiographic and soft tissue examination
Bone fractures were detected in 9 of 53 (17%) BC bats; this frequency is similar to the 9%–18% reported for bird collisions with stationary objects (eg, towers, building windows). 41 The mandible was the most frequently broken bone in BC bats (7.5%, Table 3), which is also consistent with the collision patterns of window-killed birds. 41 In contrast, bone fractures were detected in 152 of 262 (58%) WF bat cases, which was significantly greater than the fracture rate in BC bats (Fisher exact test, P < .0001). Furthermore, WF bats had increased percentage incidence in specific bone fractures compared with BC bats (Table 3). Recently, a small study of 39 WF bats reported a similarly high fracture rate of 74% as detected by a veterinary radiologist in salvaged WF bat carcasses, higher than the 20% incidence (3 of 15 non-WF bats) of free fall–related fractures. 16 Cumulatively, our data along with recent studies 4,16 suggest that WF bats have exposure to severe trauma events and are of greater blunt force than that expected for collision with stationary structures (eg windows, buildings) and subsequent free fall in bats and birds.
Bone Fracture Incidence in Building Collision and Wind Farm Bats a
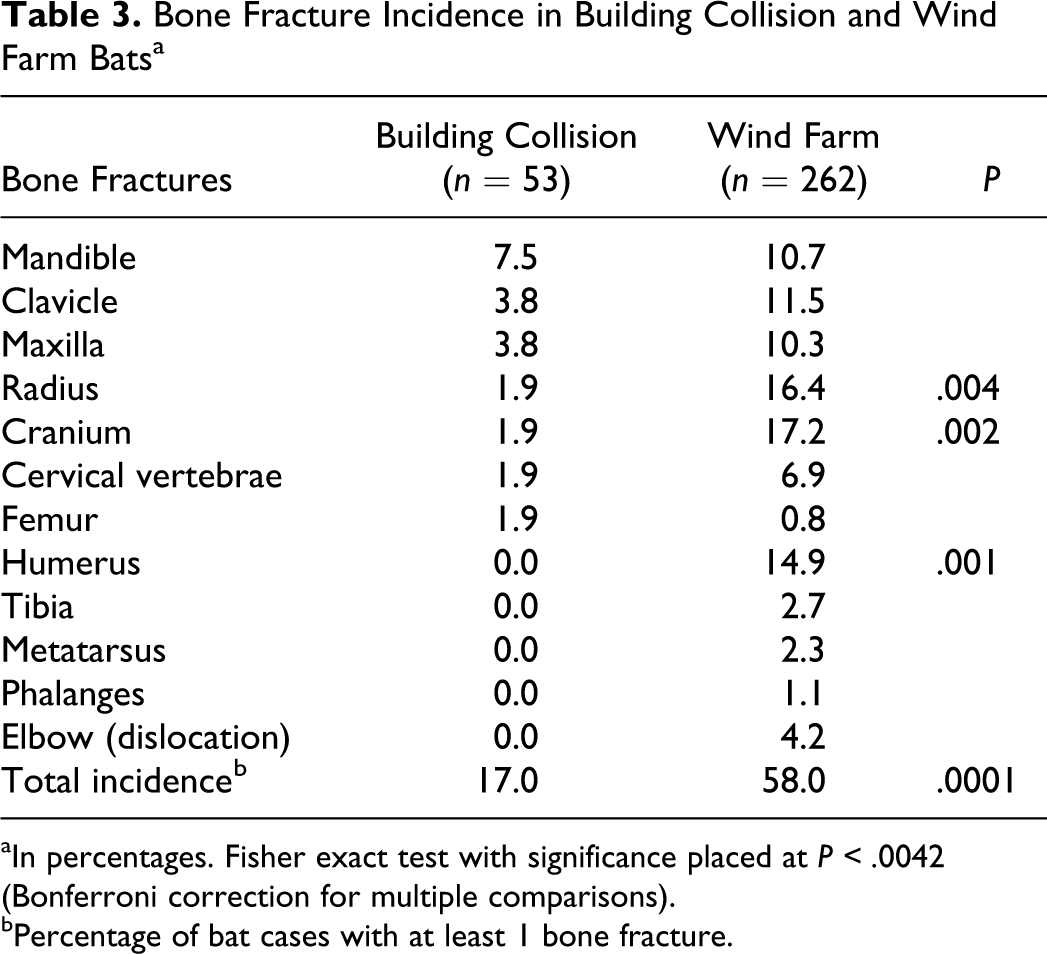
aIn percentages. Fisher exact test with significance placed at P < .0042 (Bonferroni correction for multiple comparisons).
bPercentage of bat cases with at least 1 bone fracture.
Among bats without fractures, WF bats had significantly more cases with external lacerations than BC bats (11 of 66 vs 0 of 44, respectively; Fisher exact test, P < .01) but not self-bitten tongues (2 of 66 vs 0 of 44 respectively; Fisher exact test, not significant). We performed necropsies on this subset of WF bats to examine for trauma-specific lesions; however, necropsy was not performed on BC bats because all were museum specimens. We detected subcutaneous hemorrhage in 41% (27 of 66) of WF bats (Table 4), which is a common consequence of soft tissue trauma. 20 The subcutaneous hemorrhage was distributed in a pattern comparable to the fractures incidence, suggesting that it was related to trauma and not to postmortem change. Furthermore, upon opening the chest cavity, we observed abdominal contents displaced into the thorax in 6% (4 of 66) of WF bats, consistent with traumatic diaphragmatic hernia. This condition is a marker of severe traumatic injury, is seen in approximately 5% of humans involved in automobile crashes, and may occur in the absence of concurrent bone fractures. 30
Location and Frequency of Subcutaneous Hemorrhage in Wind Farm Bats (n = 66) a

aIn percentages.
Lung assessment
All recently killed WF bats (14 of 14; cf Materials and Methods, Trauma Assessment) had some evidence of traumatic injury, including fractures (6 of 14, 43%), facial lacerations (1 of 14, 7%), subcutaneous hemorrhage (11 of 14, 79%), and bone marrow embolism (4 of 14, 29%; Figs. 12, 13). Bone marrow emboli are commonly seen with severe trauma such as bone fractures but may be present following concussive blows that lack ensuing fractures. 32 In this recently killed group, bone marrow emboli were detected within pulmonary arteries (4 of 14, 29%), but only 1 bat (1 of 4) had concurrent bone fractures. The prospective trend toward less bone marrow emboli in bats with bone fractures is at first counterintuitive but actually is consistent with a severe traumatic event (ie, struck by a moving turbine blade) resulting in instantaneous death. In such cases, there is insufficient time posttrauma for the dislodged marrow to travel via blood circulation to the lungs; this concept is often applied to forensic examination of aircraft crash victims to determine the time of death as instantaneous or shortly following the accident. 6,28 Lungs from freshly killed WF bats were also examined because microscopic lung lesions have been used as a basis for barotrauma diagnosis in bats. 4 The bat lungs had parameters similar to that seen in the mouse studies of experiment 1 (see also Figs. 1 –4), including vascular congestion (14 of 14, 100%), edema (5 of 14, 36%) and erythrocyte extravasation (14 of 14, 100%) (Table 5).
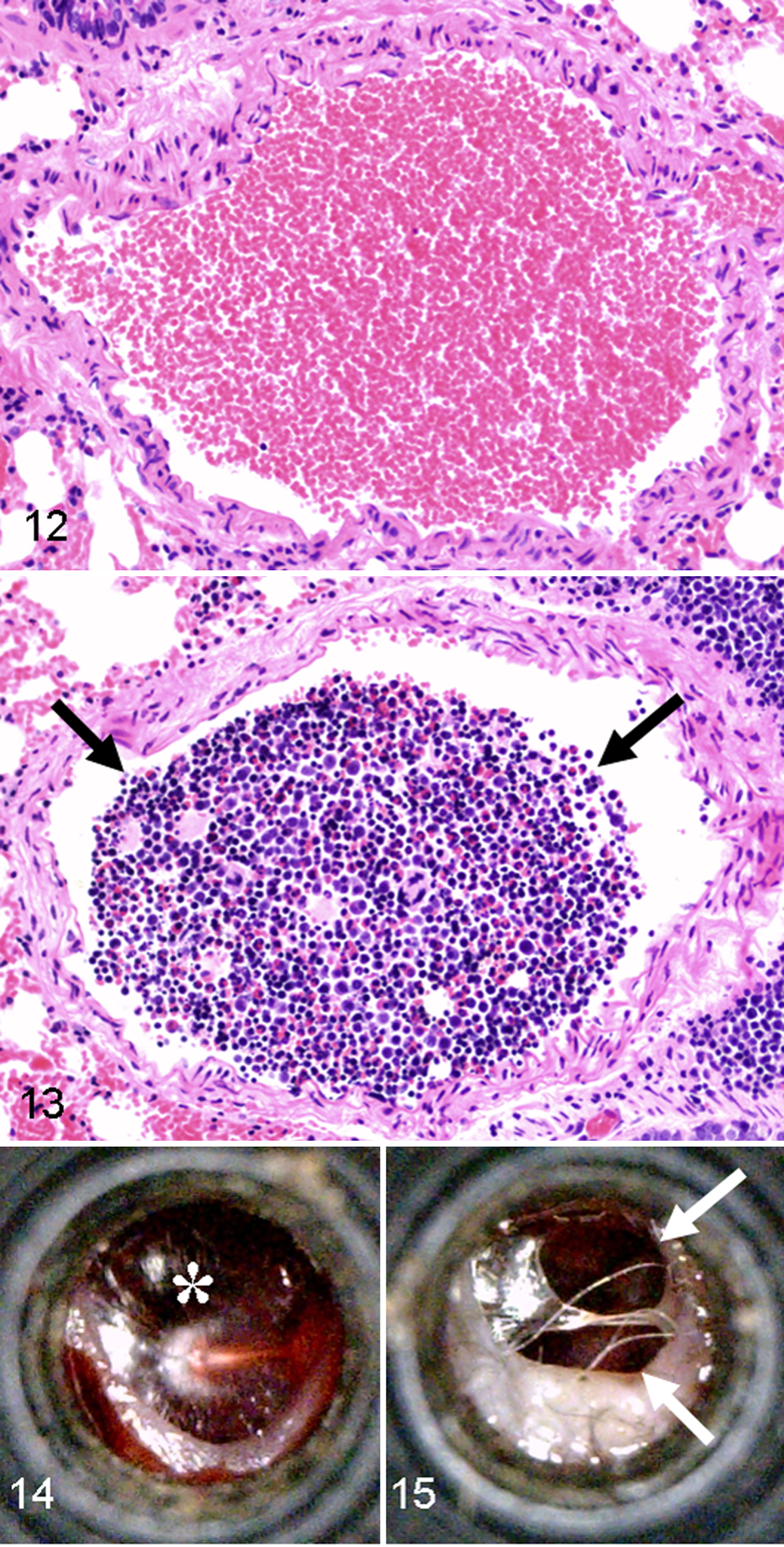
Lung, artery, eastern red bat (Lasiurus borealis); salvaged from a wind farm the morning after death, not frozen, and tissues fixed the same day. Example of a bat lung artery without bone marrow embolus. HE stain.
Histologic Scoring of Lung Parameters in Wind Farm Bats Collected the Day After Death
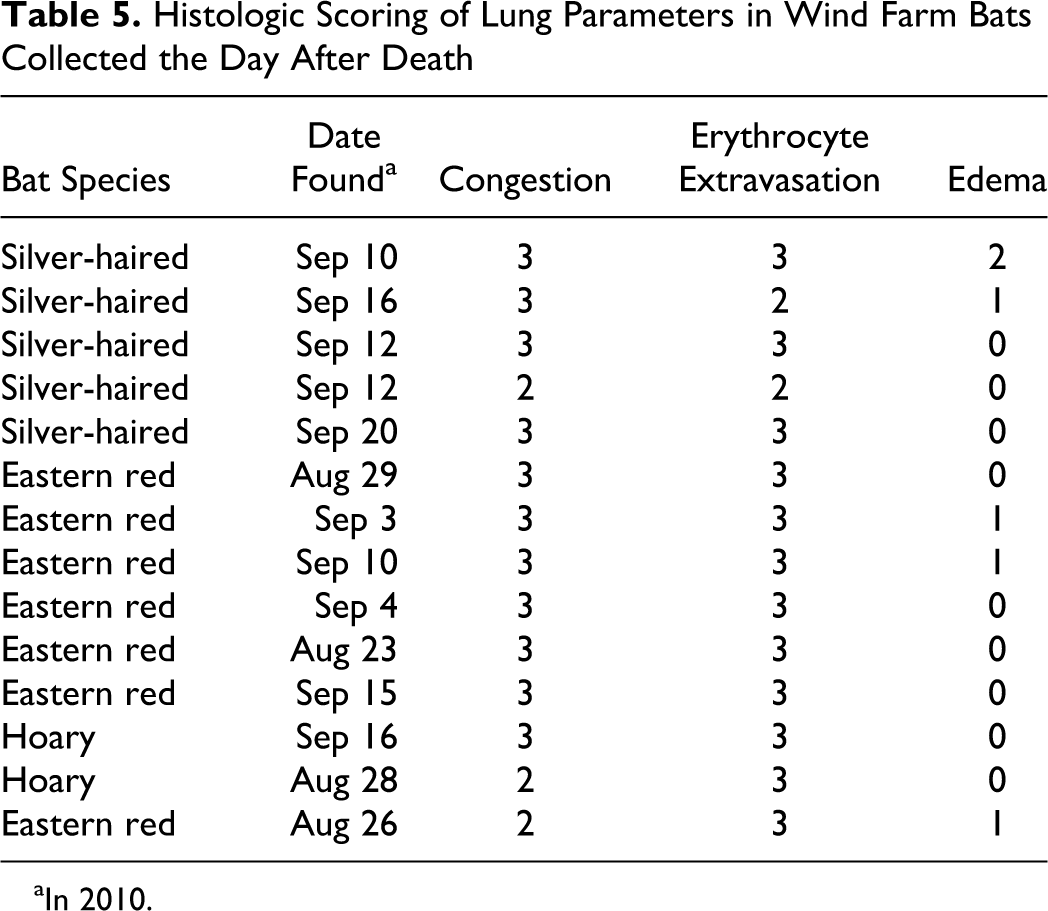
aIn 2010.
Tympana examination
Because experiment 1 demonstrated that salvaged bat lungs are inadequate for diagnosis of barotrauma, we examined tympana (ear drums) of a subset of BC (n = 42) and WF bats (n = 81). Tympana are sensitive tissues to barotrauma insults and are typically the first organ affected after a sudden change in pressure, followed by the lung. 11,18,26 In BC bats, we detected tympana rupture (bilateral) in only 1 bat (2%) that also had concurrent evidence of traumatic injuries (ie, skull fractures), which suggests that trauma may have been the cause of the rupture as has been reported to occur in humans. 17 Sixteen WF bats (20%, 3 bilateral and 13 unilateral of 81) had ruptured tympana (Figs. 14, 15). Specifically, 14% (11 of 81) of WF bats had ruptures with concurrent traumatic injuries that likely contributed to those ruptures. However, just 6% (5 of 81) of the bats had tympana ruptures without detectable evidence of traumatic injury; thus, only this small fraction of WF bats had lesions possibly consistent with a barotrauma etiology.
Discussion
Wind-powered turbines are a renewable energy resource projected to generate 20% of the US electricity demand (about 300 GW) by 2030. 39 Bat mortality significantly increases near moving wind turbines at many WFs. 3 It is estimated to be as high as 20–40 bats per turbine per season during fall migration, resulting in thousands of bat deaths at some WFs. 33 The principal etiologic cause for these deaths has not been well defined. Some scientists assume the cause to be blunt force trauma from hitting the turbine blade, and images of bats being struck by turbines have been documented. 25 However, pulmonary barotrauma has also been proposed to be the major cause for these deaths. One recent report 4 documented the first and only field evidence of pulmonary barotrauma pathology in bats; this has received widespread publicity and resulted in its acceptance as fact in some quarters.
In this study, we examined the utility of lungs from salvaged carcasses for diagnosis of pulmonary barotrauma as characterized by lung edema, hemorrhage, congestion, and/or interstitial bullae. Pulmonary edema is defined as accumulation of eosinophilic seroproteinaceous material within airspaces. This material leaks into the airspaces as a result of insults to the cellular integrity of endothelium and/or epithelium of the alveolar septa. 27,35 Freezing of bat carcasses can cause destructive ice crystal formation, while postmortem decomposition may lead to autolytic/enzymatic changes, both of which may cause tissue damage and loss of cellular integrity. 24,35 Elegant experiments in rabbits by Durlacher and colleagues 13 demonstrated that postmortem pulmonary edema appeared within just a few hours of death and that lung weights continued to progressively increase for up to 4 hours postmortem. Results of that study indicated that 2 variables influenced development of postmortem edema—alveolar wall permeability and the pressure gradient between vasculature and the airspace. Because of its confounding influence, Durlacher and colleagues proposed that pulmonary edema cannot be confidently assessed as antemortem change unless the necropsy is done immediately after death. 13
Edema was the most prominent morphologic feature within airspaces and was detected in all trials of experiment 1. Pulmonary edema has been recognized as a potential postmortem event in animals and humans 13,34,35 and may be associated with livor mortis due to increased hydrostatic pressures. 13,34 The common and potentially rapid appearance of edema in our experiment suggests that differentiation of postmortem versus antemortem edema formation would be exceedingly difficult and precarious in salvaged bat carcasses. 13
Hemorrhage is another diagnostic feature of pulmonary barotrauma. 1,4,16,29 Erythrocyte extravasation into tissues meets a classic definition of hemorrhage; however, it has been suggested that even the experienced observer cannot confidently distinguish it as having occurred in a postmortem or antemortem setting. 35 Postmortem hemorrhage tends to occur in sites of livor mortis, and it may be progressively associated with increased postmortem time. 19,35 In our mouse experiment, the presence of erythrocytes in airspaces was diagnosed as pulmonary hemorrhage, which this was common, but overt hemorrhage was uncommon. Chronologically, erythrocyte extravasation often occurred after edema leakage into airspaces of the decomposed lung (Table 1), which is logical given the progressive loss of cellular integrity during decomposition and size–permeability requirements for fluids versus cells.
The foremost differential diagnosis for bat deaths at WFs is traumatic injury, which could result in hemorrhage and edema in airspaces from direct lung injury through pulmonary contusion or laceration following rib fracture. 10,37 Furthermore, extrapulmonary traumatic injury (eg, sinonasal or oropharyngeal cavities) can lead to aspiration of blood, which also gives the appearance of lung hemorrhage. 20,38 Interestingly, external injuries consistent with traumatic injury (eg, fractures) were detected in the majority of the bats diagnosed with pulmonary barotrauma. 4
While not specifically addressed in this study, interstitial bullae are an additional lesion potentially seen in pulmonary barotrauma. However, putrefaction by gas-producing bacteria is a common postmortem event, 35,42 which can produce morphologic artifacts appearing as interstitial bullae. While the role of only a few conditions (eg, postmortem time, environmental temperatures, carcass freezing) was examined in our controlled study, other postmortem factors—such as exposure to direct sunlight or moisture (precipitation, humidity) or even their interaction under field conditions—may further influence postmortem decomposition and morphology. 42
Results from experiment 1 and a review of the literature suggest that lungs from salvaged bats are expected to be confounded by common postmortem artifacts as well as the competing etiologic hypothesis of traumatic injury (Table 6 ). Because these factors cannot be confidently excluded during pathologic examination, pulmonary barotrauma should not be diagnosed from salvaged bat carcasses.
Major Differential Diagnoses for Lung Hemorrhage/Edema From Salvaged Bat Carcasses at Wind Farms

How can researchers maximize pathology data from salvaged carcasses? (1) As previously stated, diagnosis of pulmonary barotrauma from salvaged bat lungs is not possible due to confounding artifacts and competing diagnses. Ancillary techniques, such as assessment for ruptured tympana, a tissue sensitive to barotrauma, 11,18,26 may be a useful alternative to lungs in salvaged bat carcasses. (2) Histologic markers (eg, peripheral pallor in lungs; Fig. 11) may be useful for exclusion of decomposed carcasses, but future studies are needed to determine whether it is sensitive enough to detect early decomposition-related changes in lung morphology. (3) Consistent gross and histologic examination of carcasses may further identify traumatic injuries (eg, diaphragmatic hernia and bone marrow emboli) and other etiologies. (4) Examination protocols that maximize sensitivity for lesion detection are preferred. For example, consistent use of radiographic images to assess for bone fractures is more sensitive than just external manipulation of the carcass as often done in the field. (5) Whenever possible, inclusion of control animals is useful to give proper perspective to any observed pathologic changes.
Following this framework, we were able to study salvaged WF bats in experiment 2 to determine the forensic cause of death. The control group of BC bats provided evidence that collision with a stationary object can cause death with no overt signs of external trauma or fractures (similar to bird collision data 41 ), and from this group we also had background pathology data for bat collisions with static (nonmoving) objects. In comparison, WF bats had a significantly higher incidence of fracture cases, specific bone fractures, and external lacerations than did BC bats, and 73% (190 of 262) of WF bats had lesions consistent with blunt force trauma. Collectively, these features suggest that the WF bats experienced blunt force trauma from various angles consistent with being struck by a moving blade of a wind turbine. Therefore, we conclude that the majority of WF bat deaths were caused by traumatic injury.
Interestingly, the previous description of pulmonary barotrauma pathology 4 in salvaged bats had the following: (1) Most (54%) salvaged bats had concurrent traumatic injury (eg, fractures) as assessed with less robust techniques than in our current study; (2) all salvaged bats had lung lesions (eg, hemorrhage, edema, congestion); and (3) a control group was not available. From that data set, pulmonary barotrauma was diagnosed; however, thorough consideration against leading differential diagnoses of postmortem artifact and traumatic injury was lacking. While our study does not exclude barotrauma as an etiologic differential or the potential that barotrauma can occur with traumatic injury as a combined etiology, the forensic evidence suggests that if barotrauma is a real cause of bat deaths, then it is likely of low incidence. In contrast, the forensic pathology evidence from our study and other salvaged bat data 4,16 provides compelling evidence that traumatic injury is in fact the major cause of mortality for WF bats.
Our results point to a need for improvements in the current monitoring procedures at WFs and in the experimental approaches required to answer whether barotrauma or other etiologies contribute, if at all, to these deaths. 25,33 The implications of previous pulmonary barotrauma diagnosis from salvaged bat carcasses are more than just an academic dialogue. Recent studies indicate increasing cut-in speed of turbines can reduce bats deaths; 3,5 however, significant numbers of bat mortalities still occur under these conditions and economic factors would likely prevent all WFs from enacting these mitigation measures. Further, ongoing research efforts and public debate on effective mitigation for adverse wildlife events are currently shaping regulations that affect wind energy development. 36,40 Some presume that pulmonary barotrauma is the dominant cause of bat mortality at WFs even though the field evidence is highly questionable. In fact, the US Fish and Wildlife Service recently argued that barotrauma may also result in sublethal injuries, such as hearing impairment and other internal injuries, that would allow bats to move away from turbines before debilitation and death. 40 If fatality estimators 22,40 were adjusted to account for such presumed off-site deaths due to barotrauma, total fatality estimates would significantly increase. Fatality rates are not trivial, as they are used in the postconstruction risk assessment of WFs to evaluate for negative impacts and determine whether mitigation measures are needed. For example, the Oregon Department of Energy recently developed specific fatality triggers that determine the extent of additional, prescribed mitigation efforts for WFs. 31 In light of the US Fish and Wildlife Service’s proposed iterative learning process, 40 we believe that the results of our study can help guide future surveillance of bat mortality events at additional WF sites.
Footnotes
Acknowledgements
This work was supported by Sigma Xi (G20100315152134), the Beta Lambda chapter of Phi Sigma and the School of Biological Sciences at Illinois State University. We thank staff at the Field Museum of Natural History for their help in arranging specimen loans, especially Lawrence R. Heaney and William T. Stanley. We are grateful for the excellent assistance of Matt Fraker, Brandi Read, John Vanscoyoc, Paul Naumann, Steven A. Juliano, Sandrine Clairardin, and Joseph Smaldino. We also thank S. A. Juliano, K. Gibson-Corley, A. K. Olivier, and C. F. Thompson for comments on the manuscript. We appreciate Western EcoSystems Technology, Inc. technicians who salvaged bat specimens from the wind farm.
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: The authors received financial support from Sigma Xi (G20100315152134), the Beta Lambda chapter of Phi Sigma, Office of Research and Sponsored Programs, and the School of Biological Sciences at Illinois State University.