
Abstract
Inherited osteopetrosis was identified in cattle herds in Wyoming, Nebraska, and Missouri in 2008 to 2010. Ten affected Red Angus calves were examined to characterize lesions in brain, teeth, and skull. Six affected aborted or stillborn calves were homozygous for the recently characterized deletion mutation in SLC4A2. Four affected calves were heterozygous for the SLC4A2 mutation and survived 1 to 7 days after birth. Gross lesions were similar in all 10 calves. Brains were rectangular and dorsoventrally compressed, with concave depressions in the parietal cortex owing to thickened parietal bone. Cerebellar hemispheres were compressed with herniation of the cerebellar vermis into the foramen magnum. Moderate bilateral chromatolysis affected multiple cranial nerve nuclei and, in some calves, the red nucleus. There was loss of retinal ganglion cells with severe atrophy of optic nerves. Periventricular corpora amylacea were in the thalamus, caudate nucleus, and midbrain. Vessels and neuropil in the dorsomedial aspect of the thalamus were mineralized. Dysplastic change in premolar and molar teeth comprised intra-alveolar intermingling of dentin, enamel, cementum, and bone, contributing to dental ankylosis. Changes in the heads of osteopetrotic calves are similar to those in children with malignant forms of homozygous recessive osteopetrosis.
Osteopetrosis (marble bone disease) is a family of rare heterogeneous disorders characterized by defective resorption of bone, particularly of primary spongiosa. 47 It may be appropriate to reserve the term osteopetrosis for diseases in which osteoclast defects are demonstrable, to distinguish it from other sclerosing diseases of bone. 14,30,34,41,49 Osteopetrosis is characterized by increased skeletal mass and a likelihood of fractures and neurologic complications due to compressed cranial nerves. 44 Most forms of the disease in people and animals are inherited. Infectious forms also exist, including intrauterine infection with bovine viral diarrhea virus. 38,47 The best-characterized forms of osteopetrosis occur in inbred or genetically manipulated laboratory rodents 13,49 and in rabbits. 27,33,36,40
Historically, osteopetrosis was a problem in the Aberdeen Angus breed in North America between 1960 and the early 1980s. Affected calves were born 3 weeks premature and had characteristic skeletal defects. Both red- and black-coated Angus cattle were affected. Lesions in the head included inferior brachygnathia, impacted premolars, incomplete and misaligned incisors, a protuberant tongue, fixed temporomandibular joints, and a flat calvarium. 18–20,23–26,48 The disease was inherited as a homozygous recessive trait. 19,24 There are also reports of osteopetrosis in Hereford and Simmental cattle. 24,39 Inherited brachygnathia with impacted molars is described for other breeds, such as the Shorthorn, but it is not clear whether this syndrome represents osteopetrosis. 24 Original morphologic descriptions of osteopetrosis in Aberdeen Angus concentrated on lesions in bone. Since those reports, advances in understanding the cellular and molecular basis of osteopetrosis have resulted in improved treatment. 13,14,44,49 Recent work has focused on laboratory animal models and on identifying the range of mutations in human patients. 27,29–36,41 Characterization of several mutations in mice led to the recognition of comparable mutations in people. 6,22 Establishing whether the defects are intrinsic or extrinsic to osteoclasts is important therapeutically, as it predicts the likely success of allogeneic hematopoietic stem cell transplantation. 2 Little use has been made of inherited osteopetrosis in cattle, other than to note its occurrence in reviews of the disease in mammals. 29
The Red Angus breed is derived from the black-coated Aberdeen Angus. Recently, a fatal congenital form of osteopetrosis emerged in Red Angus cattle in North America, with confirmed cases in Wyoming, Nebraska, Missouri, Kansas, and Saskatchewan. 37 It is similar or identical to the historic disease in Aberdeen Angus. Its basis in the Red Angus breed is an intrinsic defect of osteoclasts due to a 2781–base pair deletion mutation in SLC4A2, 37 which codes for an anion exchanger that exports carbonate and imports chloride. 1 Similar mutations occur in chloride/bicarbonate anion exchanger-deficient strains of mice. 21,55 Comparable mutations have not been identified in affected children. Many features of malignant autosomal recessive osteopetrosis in people resemble those of osteopetrotic Angus cattle. In children the condition is progressive and fatal when untreated. Clinical signs in the first decade of life are fractured long bones, facial palsy, deafness, blindness, myelophthisic anemia, periodic hypocalcemia, and recurrent infections. 49 Developmental delay or mental retardation is variable and of obscure pathogenesis because, with the exception of rapidly progressive “neuronopathic” osteopetrosis, children’s brains are evaluated by imaging rather than histopathology. 10,11,16,45 The histologic basis for dental disease in osteopetrotic children is poorly characterized. 12 Understanding the basis for dental defects is derived from elegant studies using rats and rabbits and from cranial imaging of children. 15,31,32,40
The reemergence of osteopetrosis in the Red Angus breed provided an opportunity to supplement the original terse descriptions of lesions in the heads of affected cattle. Recognized features were cerebral and cerebellar compression, narrow or absent cranial nerve and nutrient foramina, closure defects between frontal and parietal bones, misplaced tooth buds, calcified vessels in brain and meninges, and bony folds in the floor and roof of the cranium. 23–26,47 The purpose of the study was to characterize lesions in the heads of affected calves in greater detail and to compare the disease to its human counterpart.
Materials and Methods
Case Histories
From 2008 to 2010, we obtained carcasses or heads of 10 affected Red Angus calves from herds in Wyoming, Nebraska, and Missouri.
Herd A was a well-managed, registered Red Angus seed stock and cow–calf operation in northern Wyoming. In spring 2009, the herd consisted of 392 cows, 25 heifers, and 6 bulls. The owners had never recognized cases of osteopetrosis before the 2008 calving season. A mixture of natural service and artificial insemination was used; ovum transfer was performed from selected females. In 2008, 7 affected calves were aborted 6 to 8 weeks prematurely; 5 (Nos. 1–5) were submitted for examination. Of 7 affected calves, 5 were the progeny of 1 sire, “Romeo” (BUF CRK Romeo L081). The bull was suspected to be a carrier after a full brother sired osteopetrotic calves in Kansas and so was bred to 25 daughters (because there was no genetic test for carriers). Of 25 calves born of these test matings in 2008, 5 had osteopetrosis. The 2 other affected calves were the progeny of a full brother of Romeo, who had been mated to his dam and to a daughter of Romeo. The herd was genetically tested once the SLC4A2 mutation was defined. According to the owner, 20% of younger females in the herd were heterozygous for the mutation, reflecting recent intensive inbreeding using the Romeo bloodline. Once both bulls had been identified as carriers by production of affected progeny or testing for the SLC4A2 mutation, they were no longer used for breeding. No affected calves were born in herd A in 2009 or 2010.
Herd B was a well-managed cow–calf operation in the Nebraska panhandle. Historical annual death loss from birth to weaning was < 1%. In 2009, the herd consisted of 500 cows, 100 heifers, and 25 bulls. Only natural service was used. The herd was predominantly red coated, due to increased use of Red Angus bulls over the previous 7 years. Genetically, this was a Red Angus herd on a Hereford and Simmental background. The owner had never recognized osteopetrosis before 2009, when 2 affected male calves weighing 34 kg were born at term. One was euthanized at 1 day of age and not examined further. The other (No. 7; Table 1) was examined clinically when 15 hours old by 1 author (L.S.). Two more affected calves were born in 2010: 1 was male (No. 8) and a full sibling of calf No. 7. It was examined clinically and euthanized when 1 day old. Calves Nos. 7 and 8 had the same sire and dam, both of which were clinically normal. The dam had delivered 2 clinically normal calves in 2007 and 2008. The remaining affected calf (No. 6) was born dead at term. This 37-kg calf was identical in gross appearance to calves Nos. 7 and 8.
Case Histories of 10 Red Angus Calves With Osteopetrosis
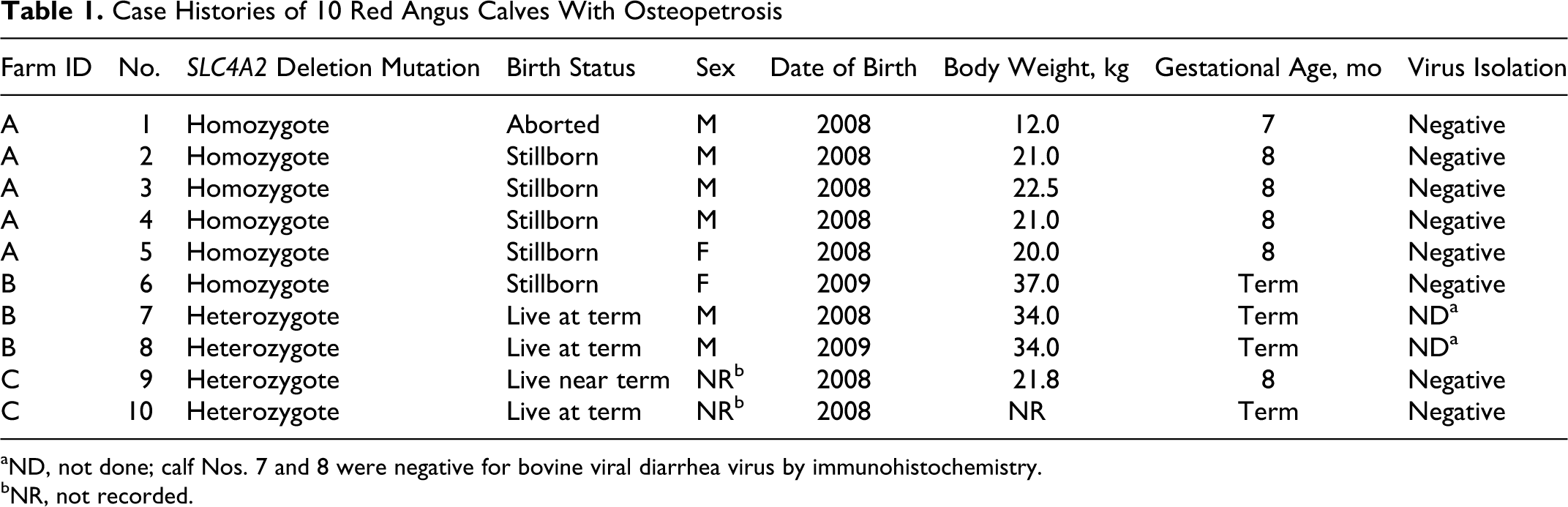
aND, not done; calf Nos. 7 and 8 were negative for bovine viral diarrhea virus by immunohistochemistry.
bNR, not recorded.
Herd C was a commercial 240-cow herd of Red Angus in Missouri. In 2008, 2 near- or full-term affected calves were born alive (Nos. 9 and 10). Neither could stand, and they died when 2 and 7 days old.
Histology and Virology
Seven carcasses (Nos. 1–5, 9–10) and the heads of another 3 (Nos. 6–8) were submitted for examination; 1 carcass (No. 10) was partly scavenged. The head of a near-term Aberdeen Angus calf that was aborted following in utero pine needle exposure was used as a source of reference tissues for teeth, cranial foramina, and subjective assessment of osteoclast numbers. (Pine needle abortion is not known to affect the skull or brain.) The brain and soft tissues from the head of each affected calf, as well as decalcified teeth and bone, were fixed in 10% neutral buffered formalin, processed routinely, and embedded in paraffin. Sections were made at 5 μm and stained with hematoxylin and eosin. Selected paraffin-embedded tissue sections were stained using the periodic acid–Schiff reaction, Perls Prussian blue reaction, von Kossa method, and Bielschowsky stain. 7,8 Sections included mandibular incisors, maxillary premolars, calvaria, and optic canals. Sections of brains from 8 calves (Nos. 1–8) were reacted immunohistochemically for bovine viral diarrhea virus antigen. 9 Fresh tissues (generally, samples of brain, liver, kidney, spleen, and lung) from 8 calves (Nos. 1–5, 8–10) were processed for virus isolation as described. 54
Results
Clinical Examination
Of 4 live-born calves, 2 (Nos. 6 and 7) were examined clinically when < 1 day old. Neither could stand or suckle, even with assistance. Both lay in lateral recumbency when unsupported. Both were blind, based on unresponsiveness to menace and on the absence of pupillary responses to light. There was no extraocular movement or strabismus. There was a sluggish blink reflex when corneas were touched. The calves responded to tactile stimulation with weak movements of the head and neck. Manipulation of the limbs elicited a withdrawal reflex. An intact but weak swallowing reflex was present. There was no practical way to assess lingual tone or response to stimulation, because of the mandibular deformation and protrusion of the tongue.
Lesions
The head and brain of affected calves were grossly similar, regardless of gestational age or SLC4A2 homozygosity or heterozygosity. Six calves (Nos. 1–6) were homozygous for the mutation; 4 (Nos. 7–10) were heterozygous (Dr Jon Beever, personal communication; Table 1). The globes were mildly proptosed, and the tongue protruded from the oral cavity. The mandibles were undershot by 2 to 4 cm. Movement of temporomandibular joints was restricted. Incisors were partly erupted, loose, and misaligned. Premolars were misaligned and covered by oral mucosa. In contrast to the jaws of normal neonatal calves, the jaws of affected calves were difficult to disarticulate without fracturing the mandibular rami. The cranial surface of the calvarium was abnormally smooth, apart from prominent bilaterally symmetrical boney ridges at the frontal–parietal synchondrosis (Fig. 1) to which there was a corresponding depression (2-cm diameter) in the temporoparietal cortex (Fig. 2). Each calf had a 1- to 2-cm fontanelle between frontal and parietal bones (Fig. 1). A bilaterally symmetrical transverse ledge of bone was at the frontal–sphenoid synchondrosis involving the orbital wing of the sphenoid (Fig. 3). There was moderate dorsoventral flattening of the brain, particularly of the medulla oblongata. The medulla oblongata at the obex, along with the posterior part of the cerebellar vermis, was herniated into the foramen magnum.
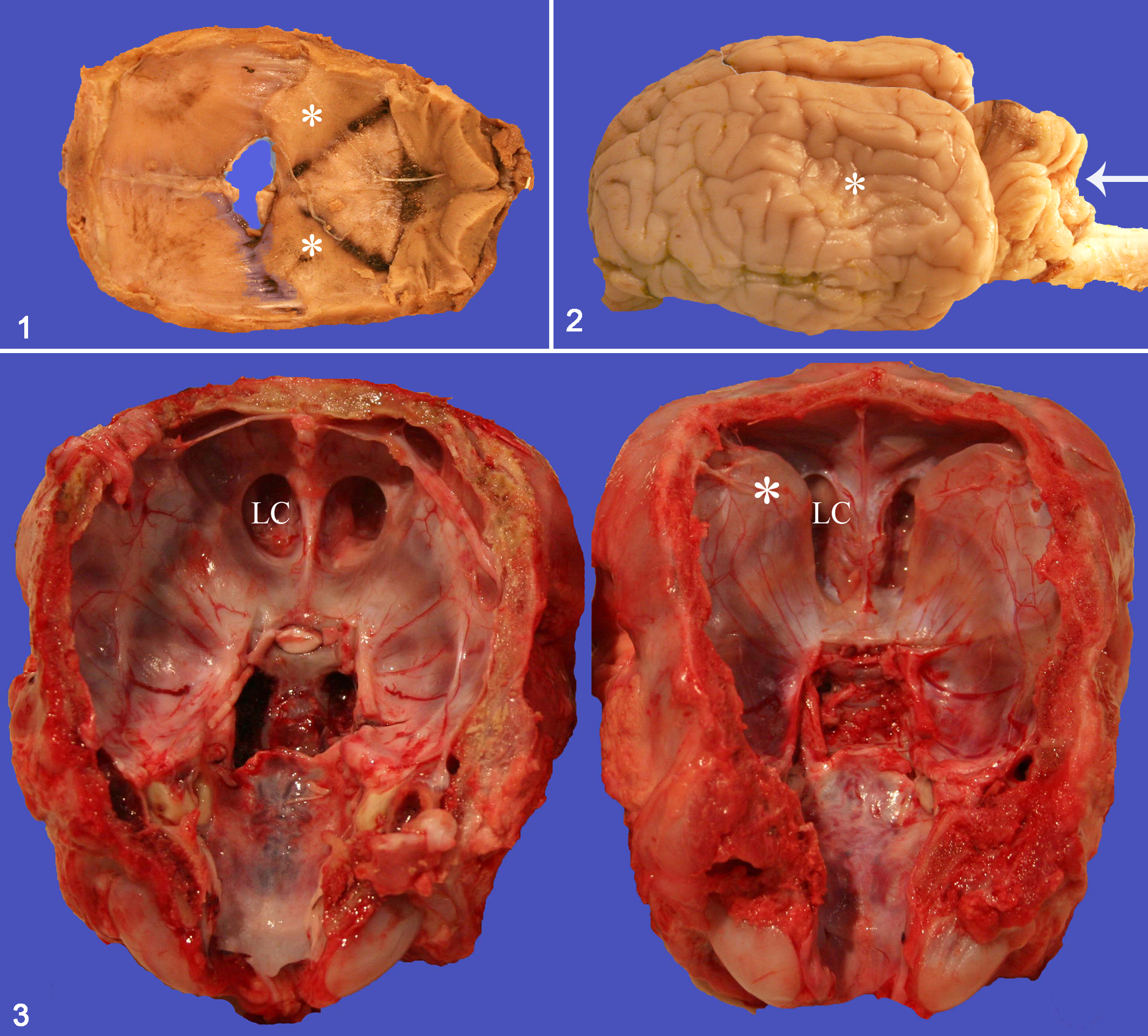
Calvarium; calf No. 3. The concave surface of the calvarium is abnormally smooth with a fontanelle between frontal and parietal bones and with bony ridges (asterisks) in the parietal bones.
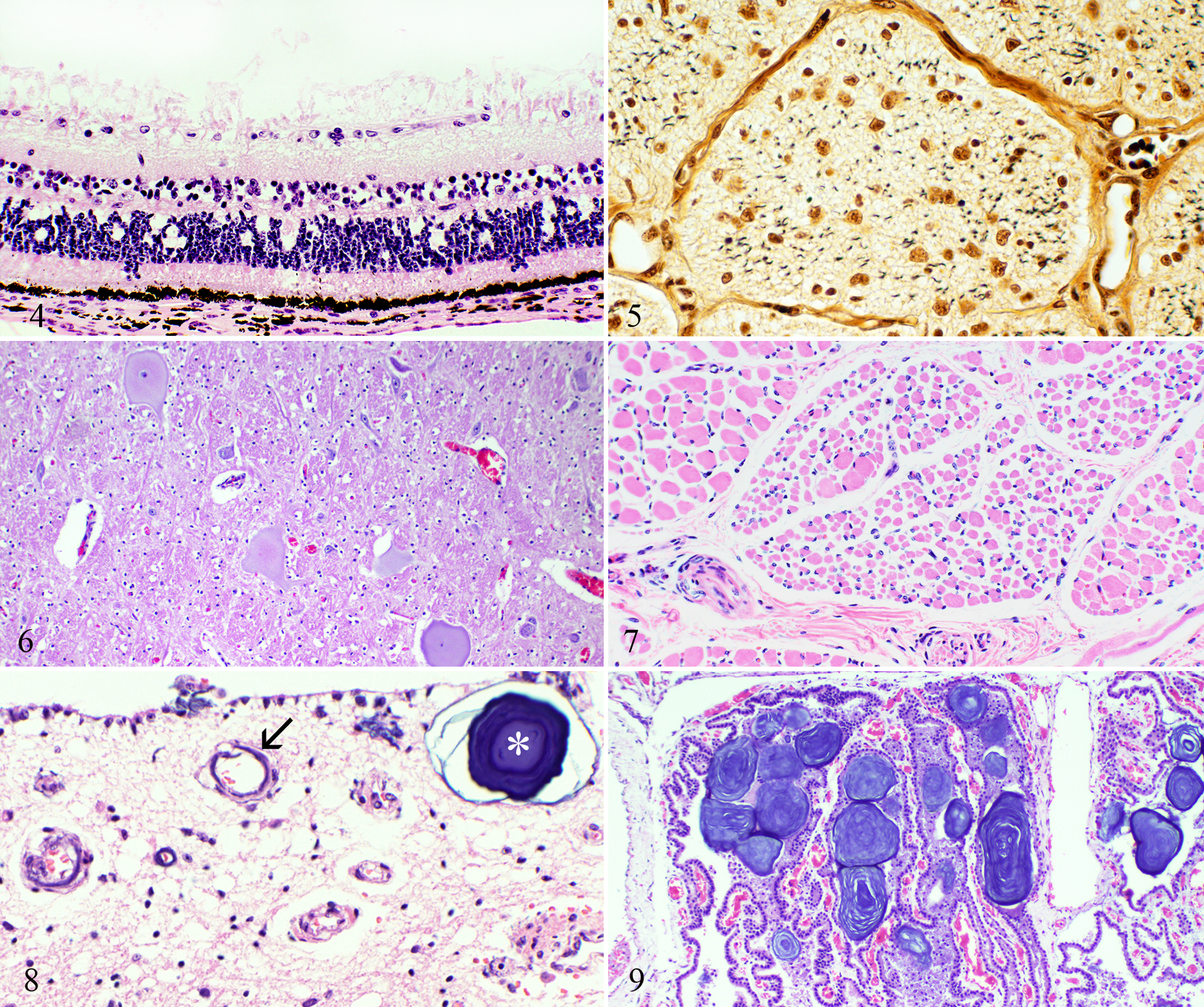
Retina; calf No. 7. Absence of ganglion cells with prominent presumptive glial cells. HE.
Mandible; calf No. 2. Trabecular bone with variably sized osteoclasts and shallow or absent resorption lacunae. Cytoplasmic vacuolation is prominent in 1 osteoclast (arrow). HE.
Histologically, there was marked reduction to absence of retinal ganglion cells (Fig. 4), accompanied by moderate to marked loss of axons and gliosis in optic nerves and optic chiasm (Fig. 5). Chromatolysis and neuronal atrophy were present in multiple nuclei of the medulla oblongata, particularly the facial, vestibulocochlear, and hypoglossal nuclei, as well as in the red nuclei (Fig. 6). This was associated with atrophy of muscles of the tongue and larynx (Fig. 7). There was axonal swelling and Wallerian degeneration in pyramidal tracts and medial longitudinal fasciculi. Perivascular paraventricular corpora amylacea were present in thalamus, basal nuclei, and midbrain (Fig. 8). Similar, larger structures were in the choroid plexus of the third ventricle (Fig. 9). Calcified vessel walls, with or without necrosis, and mineralization of neuropil were associated with the corpora amylacea (Fig. 8). There was no histologically identifiable cortical atrophy in compressed parietal cortex or in herniated cerebellum, apart from reduced numbers of Purkinje cells. Changes in the brains of the 4 calves heterozygous for SLC4A2 mutation were milder and comprised gliosis and axonal loss at the optic chiasm (Nos. 6 and 7), reduced numbers of Purkinje cells (Nos. 9 and 10), and neuronal chromatolysis in the hypoglossal nucleus (No. 7).
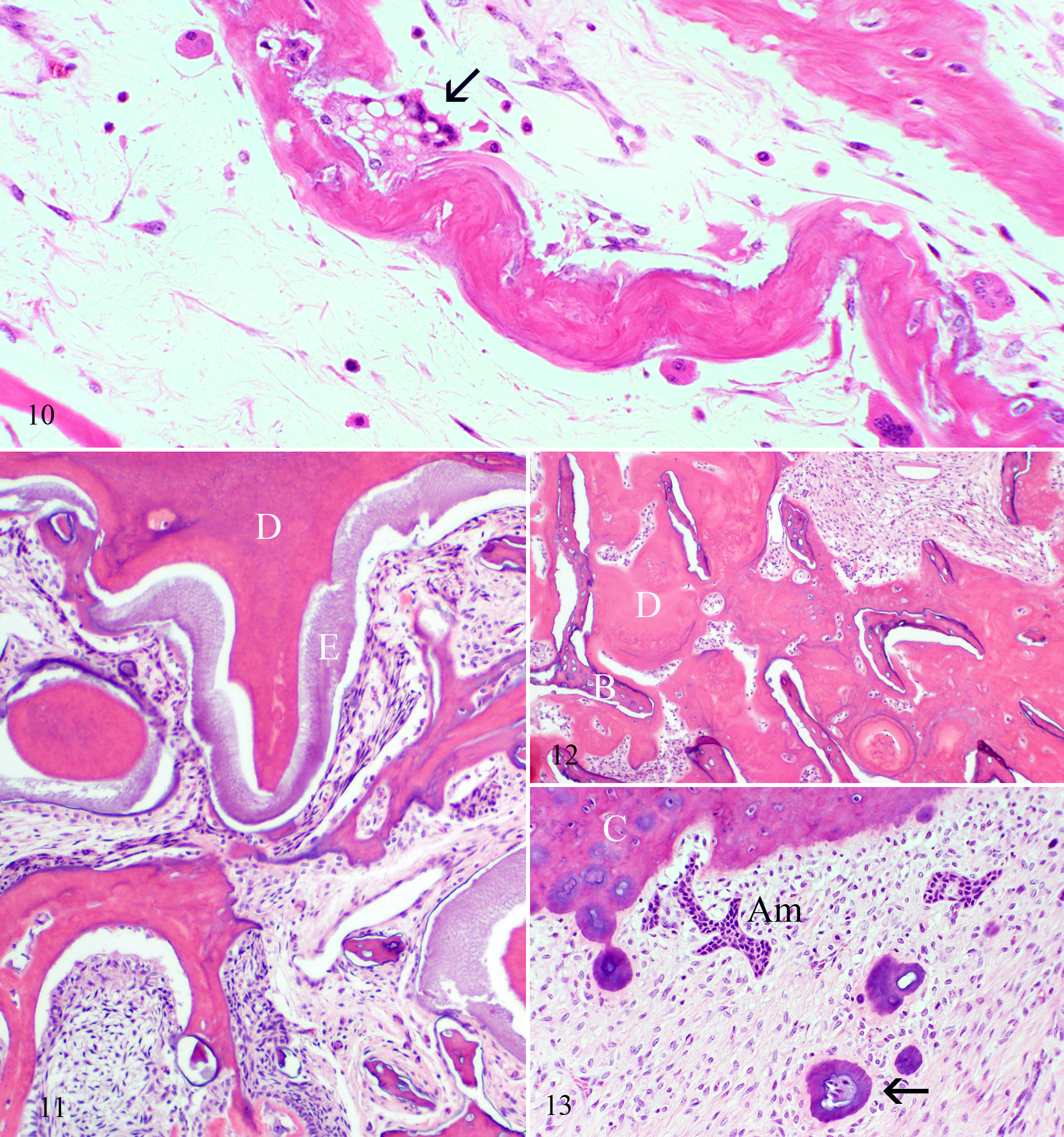
The classic histologic finding of osteopetrosis—trabecular bone with cores of hyaline cartilage 44 —was present in all 8 affected calves whose bones were examined microscopically. Unlike long bones, in which identifiable osteoclasts were rare, osteoclasts in the skull were relatively numerous. As described, 37 the histologic appearance of osteoclasts varied, particularly in size, extent of cytoplasmic vacuolation, presence of fragmented nuclei, and shallow-to-absent resorption lacunae (Fig. 10). The most noteworthy change in teeth affected the intra-alveolar portion of premolars, particularly around apical foramina. The orderly lamination of cementum, dentin, enamel, and trabecular bone in normal neonatal calves was replaced by a disorganized mixture of all components (Fig. 11). Islands of dentin, with or without margins of cementum, abutted or invested trabecular bone (Fig. 12). Anomalously, vessels were in some plates of dentin. Small pearls of cementum underlay the cementum layer, in many instances associated with aggregates of ameloblasts (Fig. 13). The crowns of teeth were normal, with unremarkable dentin and enamel. Other abnormalities in and around teeth included mineralized and ossified connective stroma in the tooth chamber, periodontal hemorrhage, and a higher ratio of acellular to cellular cementum.
Virology and Immunohistochemistry
No bovine viral diarrhea virus antigen was detected in the 8 calves examined by immunohistochemistry. No viruses, including bovine viral diarrhea virus, were isolated from brain or other tissues in 8 calves examined virologically (Table 1).
SLC4A2 mutation
Of 10 affected calves that were homozygous for the SLC4A2 mutation, 6 were stillborn or aborted (Nos. 1–6). By contrast, the 4 affected heterozygous calves (Nos. 7–10) were born alive and survived < 1–7 days (Table 1).
Discussion
Osteopetrosis of Angus cattle is likely to become rare in North America with the advent of a specific genetic test to screen for carriers. 37 One impetus for this study was to exploit the transient availability of affected animals to compare bovine to human osteopetrosis (Supplemental Table 1, available online at http://vet.sagepub.com/supplemental). Comparison is made to malignant infantile osteopetrosis, as this genetically heterogeneous group most closely resembles the disease in Red Angus cattle. Currently, it includes mutations of 3 genes that establish an acidic environment in the osteoclast resorption area; most untreated patients die in the first decade of life. Milder forms of human osteopetrosis exist, such as autosomal recessive osteopetrosis due to carbonic anhydrase II deficiency, 2,43 autosomal late-onset dominant forms (Albers-Schönberg disease), 5 and an autosomal recessive form of intermediate severity. 4 Transient and X-linked human osteopetrosis also exist. 13
Most affected Red Angus calves die in utero or at term, unlike children with infantile malignant osteopetrosis. A plausible basis for intrauterine death is compression of the brain, particularly of the medulla oblongata, compounded by herniation of the cerebellar vermis and compression of the temporoparietal cerebral cortex. Formation of corpora amylacea with associated paraventricular mineralization may contribute to neurologic impairment. The modest histologic changes in the brains of the 4 heterozygous calves, all born alive, compared to the severe lesions in the brains of homozygous calves support the notion that intracranial lesions contribute to perinatal death or abortion.
We did not attempt a comprehensive survey of all cranial nerves and their respective nuclei, given the opportunistic nature of this study, which used nonstandardized diagnostic material. Nevertheless, neuronal degeneration was consistently present in nuclei of the facial and vestibulocochlear nerves, along with loss of retinal ganglion cells and atrophy of optic nerves. Human patients may have compression of cranial nerves I–VIII. 45 We were unable to identify the site (or sites) of compression in optic canals. Perhaps bony compression during gestation results in sufficient atrophy of optic nerves such that, at birth, stricture sites are no longer apparent. Compression in the optic canals is the generally accepted basis for blindness in children with juvenile osteopetrosis and the rationale for surgical amelioration by deroofing the orbit. 3,42,45 Increased intracranial pressure due to obstructed cerebral venous outflow may be contributory in some patients. 42,45 In the absence of an identifiable site of compression in the optic canals, we wonder whether dorsoventral compression and/or posterior displacement of the optic chiasm with stretching of the optic nerves and their blood supply contributes to atrophy of the optic nerves.
Four calves were born alive with profound neurologic impairment. Live-born osteopetrotic calves are considered rare; Leipold’s group described 2 live-born affected calves that survived 12 hours. 24 In children, in addition to cranial nerve atrophy, neurologic signs are generally due to a ceroid lipofuscinosis-like neuroaxonal dystrophy or to hydrocephalus. 45 Varying degrees of cerebral atrophy and absence of the corpus callosum occur in some patients. In affected calves, there was no evidence of lysosomal storage disease, hydrocephalus, or agenesis of the corpus callosum. Children and adults with some forms of osteopetrosis, particularly those with carbonic anhydrase II deficiency, have distinctive multifocal paraventricular and cerebellar calcification. 10,11,43,49 Affected calves had mineralized vessels in the thalamus and corpora amylacea in the thalamus, basal nuclei, midbrain, and choroid plexus of the third ventricle. These modest, localized lesions occur in some of the same locations of mineralization in some forms of human osteopetrosis (“marble brain disease”). Calves, unlike human patients, had no involvement of the cerebral cortex, subcortical white matter, dentate nucleus, putamen, or globus pallidus. 10,11,43,49,52 In osteopetrotic children, intracerebral mineralization is attributed to decreased carbonic anhydrase activity in neurons or to systemic acidosis. 43 We have no explanation for its occurrence in calves, other than to note that the HCO3/Cl– anion exchanger is expressed in many epithelial cell types, including choroid plexus epithelium. 1 The pronounced paraventricular distribution is consistent with the location of corpora amylacea due to other causes. 17 Some neurologic impairment in children with osteopetrosis is attributed to stenosis of cranial arteries and venous drainage routes. 45 No gross or histologic evidence of such stenosis was seen in affected calves.
There are no published accounts of the histologic basis for dental abnormalities in bovine osteopetrosis. Dental disease is important in affected children, who are prone to caries, secondary osteomyelitis, malocclusion, and persistent primary dentition. Reported defects in affected children, based largely on radiology and histologic studies of extracted teeth, are malformed roots and crowns, missing or poorly calcified teeth, abnormal pulp spaces, reduced enamel, absent tooth buds, twisted dentin tubules, irregular enamel prisms, and thick lamina densa. 12,15,50 Delayed or arrested dental eruption is a feature of most but not all mouse strains with osteopetrosis. 6,22 Normal dental eruption with failure of enamelization and with disorganized ameloblasts is reported in 1 SLC4A2 knockout strain. 28 A useful gross diagnostic feature of osteopetrosis in affected Angus calves is unerupted teeth with crowding and abnormal alignment, combined with brachygnathia. Histologic changes were most marked in premolars and molars, with dysplastic intermingling of dentin, enamel, cementum, and bone at the base of the pulp chamber, which resulted in dental ankylosis. Failure to form an eruption pathway in bone overlying teeth, due to defective osteoclasts, occurs in the osteopetrotic (os) rabbit and incisors-absent (ia) rat. 31,32,35,40 This causes continuous proliferation of dental tissue in the interstices of alveolar bone, leading to dental ankylosis. 35 This probably occurs in Red Angus cattle. There was no histologic evidence that dental tissue was reduced in Angus calves.
Most forms of human autosomal recessive osteopetrosis, affecting the TCIRG1-encoded proton pump and the LCN7-encoded voltage-gated chloride channel, are associated with normal or increased numbers of osteoclasts and impaired acidification of resorption lacunae. 13 In affected calves, the number of osteoclasts in long bones is decreased, as in the less common osteoclastopenic forms of human osteopetrosis. 49,51,53 Though difficult to find in long bones, osteoclasts were readily found in the skull and mandibles, suggesting differences in the ability of separate osteoclast subpopulations to mature or persist in affected calves.
A stereotypical feature of bovine osteopetrosis is the presence of 2 bony ridges in the cranial cavity abutting the frontal–occipital and frontal–parietal sutures. The simplest explanation for these is differential growth and/or resorption. These features have not been reported in the skulls of affected children. 15,16
An unexpected finding was that 4 of 10 affected calves were heterozygous for the SLC4A2 mutation. The recent report of the genetic basis for osteopetrosis in cattle concluded that the disease was restricted to cattle homozygous for the defect. 37 One explanation for a phenotypically similar disease in calves regardless of whether they have one or both copies of the mutation is that affected live-born calves are compound heterozygotes. An analogous situation occurs in some affected children with intermediate to severe osteopetrosis phenotypes who are heterozygous for mutations of the CLCN7 gene. 46,49 Work is ongoing to establish the basis for osteopetrosis in affected Red Angus calves that are heterozygous for the mutation (J. Beever, personal communication).
In summary, bovine osteopetrosis is associated with stereotypical lesions of brain, teeth, and skull, regardless of whether calves have one or both copies of the SLC4A2 mutation. Heterozygotic calves differ from homozygotes in their tendency to be born alive and to have milder lesions in the brain. Dorsoventral compression of the medulla oblongata and degeneration of cranial nerves, combined with cerebellar herniation and compression of the parietal cortex, is the likely basis for abortion, stillbirth, and perinatal death. Many similarities exist between bovine and human osteopetrosis, as noted by Leipold. 23 One exception is that most forms of malignant osteopetrosis in people are associated with normal or excessive numbers of osteoclasts, 14 whereas in cattle the number of osteoclasts, particularly in long bones, is reduced. Now that the mutation in cattle is defined, there may be merit in using the bovine disease to study the pathogenesis of osteopetrosis.
Footnotes
Acknowledgements
Portions of this study were reported in BMC Genomics (May 27, 2010; 11:337) and are published here with permission from the journal. This study was supported by the US Department of Agriculture’s Animal Health program (WYO-397-05). We are grateful to the owners of herds A and B for sharing pedigree information and to Dr Jon Beever at the Laboratory of Molecular Genetics, Department of Animal Sciences, University of Illinois, Urbana-Champaign, for information on the SLC4A2 mutation status of the calves. We thank Paula Jaeger, Mercedes Thelen, and Katy Fambrough for histologic preparations and Jacqueline Cavender and Marce Vasquez for microbiology.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.