
Abstract
During the severe acute respiratory syndrome (SARS) outbreak of 2003, approximately 10% of SARS patients developed progressive respiratory failure and died. Since then, several animal models have been established to study SARS coronavirus, with the aim of developing new antiviral agents and vaccines. This short review describes the pathologic features of SARS in relation to their clinical presentation in human cases. It also looks at animal susceptibility after experimental infection, animal models of SARS, and the pathogenesis of this disease. It seems that adaptation of the virus within the host animal and the subsequent abnormal immune responses may be key factors in the pathogenesis of this new and fatal respiratory disease. The proteases produced in the lung during inflammation could also play an important role for exacerbation of SARS in animals.
Keywords
The first case of severe acute respiratory syndrome (SARS) was identified in November 2002 in Guangdong Province, China, and a similar outbreak was reported in Hong Kong in March 2003. By April 2003, the epidemic had rapidly spread across the world. 10,23,28,50,70 In September 2003, the World Health Organization reported 8,422 cases and 916 deaths in 30 countries as a result of the SARS epidemic.
Within 4 weeks after the global outbreak began, a novel coronavirus was isolated from respiratory specimens taken from SARS patients and cultured in FRhK-4 cells (fetal rhesus kidney) or VeroE6 cells (African green monkey kidney cells). Particles typical of coronavirus were detected by electron microscopy, measuring 80 to 140 nm in diameter with complex, peripheral, surface projections of 20 to 40 nm. Based on the results of animal experiments (including viral infection of monkeys) and fulfillment of Koch postulates, this coronavirus was designated as SARS-coronavirus (SARS-CoV). 11,25 In these animal experiments, SARS-CoV was administered intratracheally and intranasally to cynomolgus macaques (intratracheal inoculation was the most effective delivery system to the lower respiratory tract), which resulted in respiratory and constitutional clinical signs (mild cough, sneezing, high body temperature, lethargy, slight decreased activity), viral shedding, and pulmonary lesions.
Soon after the monkey experiments, cats, ferrets, mice, pigs, hamsters, guinea pigs, hamsters, chickens, and rats were infected to study various pathogenetic aspects of the disease. All these animals were susceptible to SARS-CoV infection after intrarespiratory inoculation, and most exhibited excretion of the virus in pharyngeal or nasal swabs, histopathologic pulmonary changes, and seroconversion. However, the course of the infection in these animals was shorter than that in humans, and none of these animal models replicated the human disease in all its aspects. Some laboratories have tried to establish an animal model for SARS. This short review presents a summary of the pathologic studies in human cases and an overview of the experimental animal models developed to understand the pathogenesis of SARS-CoV infection.
Human Cases of SARS
Clinical Features
Common symptoms of SARS include fever, a nonproductive cough, myalgia, and dyspnea. Poor outcomes are associated with the presence of comorbidities, male sex, age over 60 years, high neutrophil counts, and severe biochemical abnormalities. The incubation period may vary from 1 to 14 days (mean, 4–6 days). A prospective study of one outbreak found that the clinical progression of SARS was mostly uniform and followed a triphasic pattern. 49 Fever, myalgia, coughing, and a sore throat characterized the first week, followed in the second week by frequent recurrence of fever, diarrhea, and hypoxemia. Half the patients had abnormal chest radiographs. Virus shedding from the respiratory tract peaked on the 10th day after onset and was excreted in the feces from 12 to 14 days after onset. Immunoglobulin G seroconversion occurred 10 to 15 days after onset and correlated with a decreased viral load. Some patients showed clinical worsening at this stage and developed acute respiratory distress syndrome by week 3, although this can develop within the first week. The most severe cases died between 4 and 108 days after the onset of symptoms. Research suggests that immunopathologic damage from an overexuberant host immune response, rather than uncontrolled viral replication, was the major cause of lung damage at this stage. 48,50,69,77
Pathologic Changes
The autopsy results of some SARS patients have been released since the first outbreak, in 2003. The major pathologic finding in these cases was diffuse alveolar damage (DAD). This acute lung injury was associated with viral replication and immunopathogenic factors. Histopathologically, the lungs of SARS victims showed various stages of DAD that are related to the duration of the illness, such as exudation, regeneration, and repair. During the early phase of the illness (7–10 days after the onset of symptoms), the lungs showed signs of severe damage, including hyaline membrane formation, extensive edema, fibrin exudation, and some infiltration by inflammatory cells (Fig. 1). Cellular infiltration by macrophages has been observed, as has a combination of macrophages and lymphocytes, with or without neutrophils. 6,8,17 –19,48,58,69 Large multinucleated cells identified as macrophages and pneumocytes by immunohistochemistry are frequently observed in the lungs of SARS patients. Bronchiolitis obliterans organizing pneumonia, 48 hemophagocytosis in the mononuclear cells residing in pulmonary tissues, 48 and apoptosis of epithelial cells, monocytes/macrophages, lymphocytes and pneumocytes 17 are also seen.
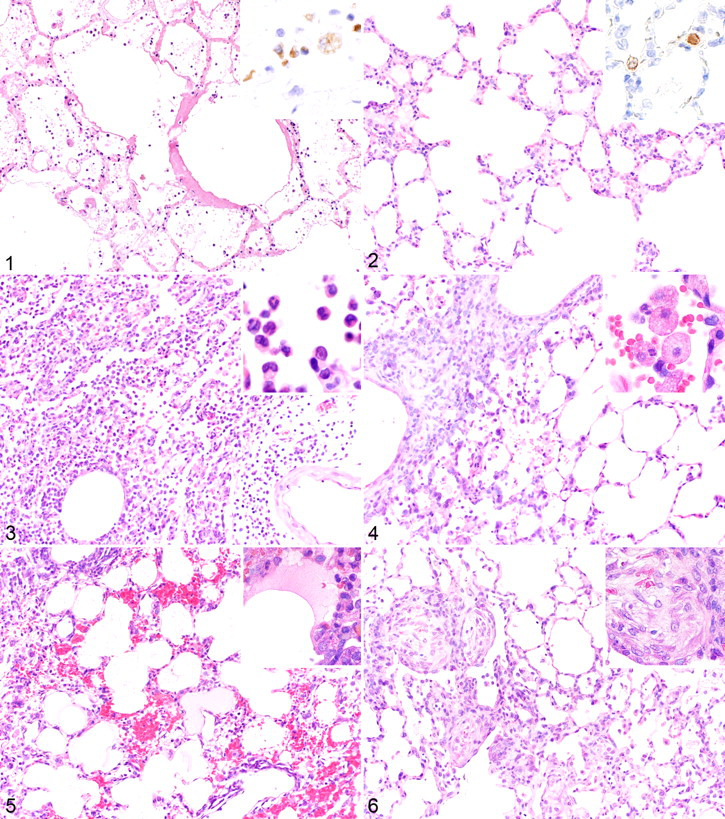
In some cases, coinfections by Aspergillus sp, Mucor sp, Pseudomonas aeruginosa, Klebsiella sp, methicillin-resistant Staphylococcus aureus, α-hemolytic Streptococcus sp, and cytomegalovirus have been reported. 6,19 In situ hybridization and immunohistochemistry studies have identified viral nucleic acid and/or antigen in epithelial cells from bronchi, bronchioles, trachea, and alveoli and in multinucleated cells and alveolar macrophages. 16,17,47,58,66,75 Investigation of the cytokine and chemokine profiles of SARS patients has suggested that severe SARS-related lung injury might be attributed to an excessive host immune response—particularly, the deregulation of proinflammatory cytokine production. 48,50,68,77
Wild Animals and Domestic Animals in the Market
The susceptibility of wild animals, civets (Paguma larvata), raccoon dogs (Nyctereutes procyonoides), and Chinese ferret badgers (Melogale moschata) that were involved in a market-related SARS outbreak were assessed for their susceptibility to SARS-CoV. 46 SARS-CoV-like virus was identified in swabs taken from Himalayan palm civets (P larvata) and raccoon-dogs (N procyonoides) using reverse transcriptase polymerase chain reaction (RT-PCR). Both these species, along with the Chinese ferret badger, had neutralizing antibodies to animal coronavirus, and the Himalayan palm civet serum samples were positive for anti-SARS-CoV antibodies on Western blots. 46 The discovery of coronaviruses genomically related to SARS-CoV isolated from humans (> 99% homologous at the nucleotide level) in masked palm civets raises the possibility that civets play a role in the transmission of SARS-CoV to the human population. 71 After intratracheal and intranasal inoculation with human SARS-CoV, civets became lethargic and less aggressive, with a raised body temperature and leucopenia. Histopathologic examination showed lesions of interstitial pneumonia by days 13 to 35 postinoculation. Civets were equally susceptible to SARS-CoV isolated from other civets and from humans.
Ferrets (Mustela furo) and domestic cats (Felis domesticus) were also susceptible to SARS-CoV after intratracheal inoculation. 37 Interestingly, noninoculated ferrets and cats housed with inoculated animals became infected with SARS-CoV. Both species showed evidence of virus excretion on RT-PCR of pharyngeal swabs 2 days after infection, peaking 6 to 8 days after infection. Cats had seroconverted by day 28. However, the ferrets were lethargic, developed conjunctivitis, and died 16 to 21 days after infection. On pathologic examination, these ferrets showed marked hepatic lipidosis and emaciation. These data obtained from cats and ferrets suggest that the virus is efficiently transmitted to animals cohabiting with infected individuals.
Because of speculation that the virus originated in domesticated animals, pigs and chickens were used for experimental infection with SARS-CoV; 66 however, the results of such studies showed that domesticated animals were not amplifying hosts for SARS-CoV.
In 2005, horseshoe bats (Rhinolophus sinicus) were identified as a natural reservoir for a group of coronaviruses distantly related to SARS-CoV. The genome sequence of this SARS-like coronavirus is approximately 88 to 92% homologous at the nucleotide level with that of the SARS-CoV. 26
Study of SARS in Animal Models
Soon after experiments were performed in monkeys according to Koch postulates, 11,25 cats, ferrets, mice, pigs, chickens, hamsters, guinea pigs, and rats were experimentally infected with SARS-CoV in an attempt to develop animal models in which disease pathogenesis and preventative strategies could be evaluated. 33,34,37,56,66,67 Although these infected animals lacked severe acute pulmonary illness (in contrast to what is typical of SARS), they showed virologic and serologic evidence of SARS-CoV infection, yet some had mild symptoms and showed evidence of virus shedding. Note, however, that the research groups used different viral isolates to infect these animals (as in the monkey experiments described in the following sections); therefore, it is not clear whether one animal model is better than another for SARS-CoV infection. Nevertheless, these studies still demonstrated the diverse range of mammalian species susceptible to experimental infection by SARS-CoV, and these animals may be useful models for acute SARS-CoV replication and associated pathology and for insights into the early events in SARS-CoV infection.
Experimental SARS-CoV Infections in Monkeys
A nonhuman primate model would be a useful research tool to study the pathogenesis of SARS and for the development of vaccines and other therapeutic agents. After successfully demonstrating SARS-CoV infection and disease in cynomolgus monkeys (Macaca fascicularis), 11,25 some research groups investigated the susceptibility of other Old World monkeys, such as rhesus monkeys (Macaca mulatta), 39,51,57 African Green monkeys (Cercopithecus atheiops or Chlorocebus sabeus), 39 the New World monkey, and the common marmoset (Callithrix jacchus). 15 In most of these experiments, the clinical manifestations of SARS presented as mild respiratory symptoms at worst or were not observed at all. To date, there is no monkey model of lethal SARS infection.
However, these monkey experiments demonstrated some discrepancy regarding clinical manifestations and pathologic changes (see Table 1 ). Some researchers concluded that the macaque model was of limited use for the study of SARS pathogenesis and for the evaluation of new therapies, 27,36,39,42,57 but others reached different conclusions. 11,25,51 These differences of opinion may be due to the strains of SARS-CoV and/or the subspecies of monkey used in these studies. In a comparative study of 3 species of Old World monkey experimentally infected with SARS-CoV, the virus replicated to a higher titer and for a longer time in the respiratory tract of African Green monkeys than in that of rhesus and cynomolgus monkeys. 39 However, all these Old World monkeys are known to be susceptible to SARS-CoV, and all showed seroconversion by day 28 postinfection. In the lungs of infected monkeys, a strong innate immune response characterized by activation of the genes for various proinflammatory cytokines and chemokines—including IL-6, IL-8, and IP-10—was induced during the early phase of SARS-CoV infection. 7 Phosphorylation of signal transducer and activator of transcription 1 and production of IFN-β were also seen in the lungs. These results indicate that the activation of this antiviral pathway at the site of viral replication might be critical to develop protection against SARS-CoV infection.
Summary of Published Reports of Experimental Infection of Monkeys to Determine Their Susceptibility to Severe Acute Respiratory Syndrome Coronavirus a
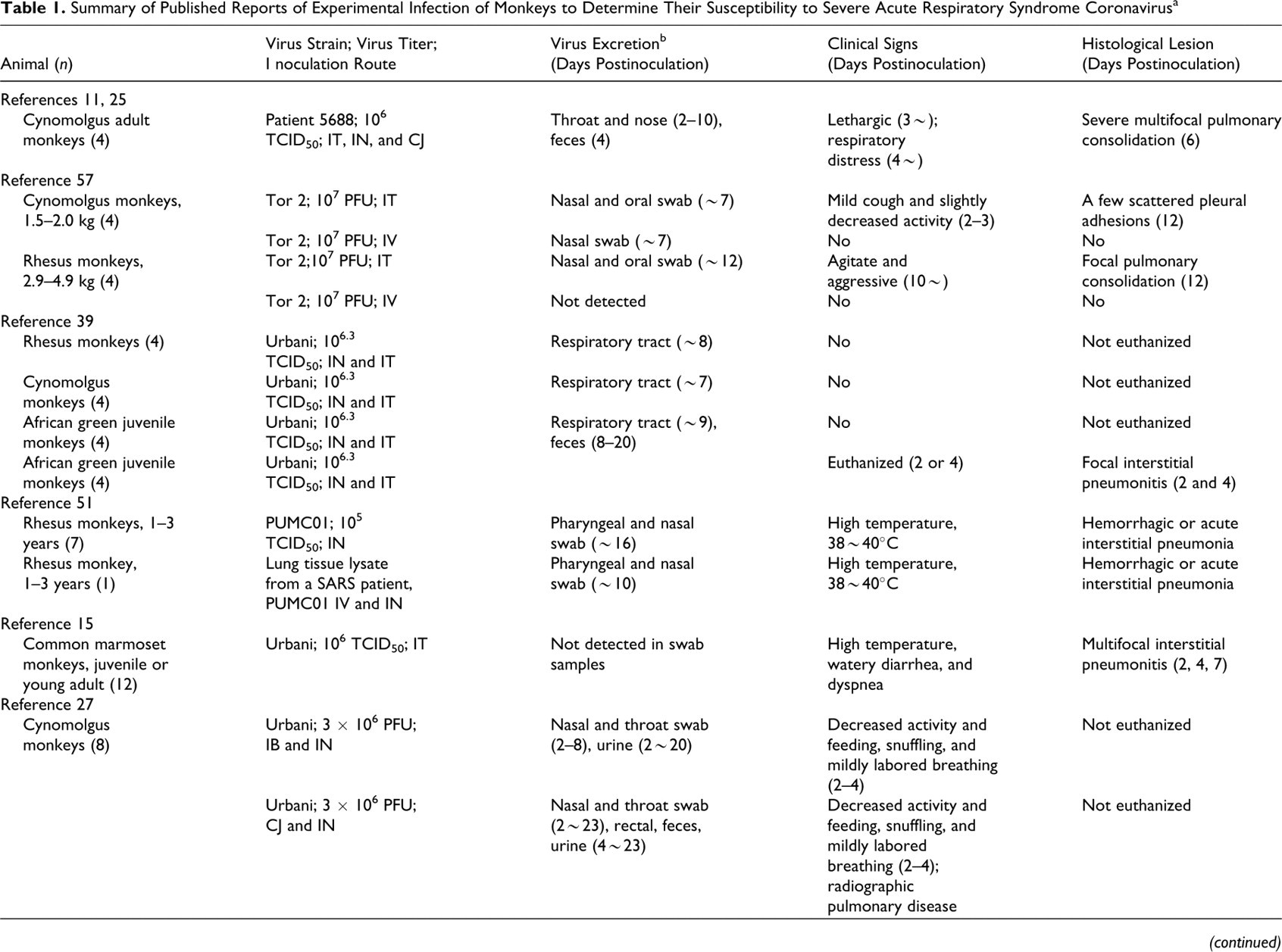
a CJ, conjunctival infection; IT, intratracheal inoculation; IN, intranasal inoculation; IV, intravenous inoculation; IB, intrabronchial inoculation; PFU, plaque-forming unit.
b Based on reverse transcriptase polymerase chain reaction or virus isolation.
c Two monkeys were subsequently inoculated 25 days after the first intranasal inoculation.
d Two monkeys were subsequently inoculated 35 days after the first intragastric inoculation.
e One monkey showed virus excretion.
Development of Vaccine and Therapeutic Agents to SARS-CoV Using Nonhuman Primate Models
Nonhuman primate models for the development of a SARS vaccine (using macaques) 5,12,35 and other therapeutic agents have been reported. 52,64,72 An adenoviral-based vaccine was shown to induce SARS-CoV-specific T-cell and virus neutralizing antibody responses in a rhesus monkey model. 12 Another study assessed, in African Green monkeys, an existing live attenuated human parainfluenza virus type 3 vaccine candidate as a vector for the expression SARS-CoV S protein. 5 After mucosal immunization via the respiratory tract, these animals produced SARS-CoV-neutralizing antibodies. The vaccine efficacy was assessed with African Green monkeys via challenge experiments, in which the immunized monkeys did not shed SARS-CoV after the initial challenge. Cynomolgus monkeys were used to evaluate the immunogenicity and protective efficacy of a purified inactivated SARS vaccine, 52 which induced high levels of neutralizing antibodies and prevented the replication of SARS-CoV within the lymph nodes, spleen, lung, and kidney. The recombinant nucleocapsid protein of SARS-CoV has been evaluated in BALB/c mice and monkeys in immunogenicity studies, 35 and the immunodominant B-cell epitopes and functional cytotoxic T-lymphocyte epitopes of this protein were identified in both species. Rhesus monkey models have been used to develop therapeutic agents for SARS-CoV infection using short interfering RNA inhibitors or equine anti-SARS-CoV F(ab′) antibody fragments. 29,64 Such approaches provided relief from elevated body temperature, diminished the number of SARS-CoV-antigen-positive cells (identified by immunohistochemistry), and reduced acute respiratory damage in these monkeys, which validated this animal model for the evaluation of effective treatments against SARS-CoV infection.
Experimental Infection of Small Experimental Animals
Rodents have been used for the development of vaccines and antiviral agents against SARS-CoV. 13,14,63,74 In murine models, intranasal inoculation with human isolates resulted in little histopathologic evidence of disease, despite the relatively high viral concentrations found in lung tissues. However, the reductions in viral titer seen in these models shows the effectiveness of vaccines and neutralizing antibodies. 67 To study the mechanisms underlying the clearance of SARS-CoV, C57BL/6 (B6), beige, CD1–/–, and RAG1–/– mice were experimentally infected with the virus. 13 Surprisingly, all the mice cleared the virus, suggesting that NK cells and adaptive cellular immunity do not play a role in clearance. Young BALB/c mice also developed asymptomatic or mild respiratory tract infections after intranasal inoculation with human viral isolates. However, adult BALB/c mice (12–14 months old) developed symptomatic pneumonia with severe weight loss, which correlated with the age-related susceptibility to lethal SARS infections in humans. 54
Syrian golden hamsters developed acute interstitial pneumonia and consolidation after intranasal administration of 103 TCID50 of SARS-CoV. 56 This consolidation, in association with virus replication, was most extensive on day 7 postinfection. Following low-titer SARS-CoV infection, hamsters showed pulmonary lesions, such as inflammatory infiltration with mononuclear cells in the alveolar septa and peribronchial pneumonic consolidations, along with consistently high levels of viral replication. As small outbred animals, hamsters were thought to be a better model than that of mice for evaluating the therapeutic effects of different treatment regimens. As such, immunotherapy using monoclonal antibodies against SARS-CoV was evaluated in this hamster model. 55 Postexposure monoclonal antibody therapy was also shown to be successful in this animal model, making use of virus replication in the respiratory tract and associated pulmonary pathologic findings.
New Animal Models for SARS Using Rodents-Adapted SARS-CoV
Roberts et al developed and characterized a mouse-adapted strain, MA15, from a human strain called Urbani isolate. 53 This virus was lethal for young female BALB/c mice (6–8 weeks old) after intranasal inoculation. Compared with the original Urbani isolate, the MA15 strain has 6 coding mutations that might be responsible for the adaptation and increased virulence of this strain. With another SARS-CoV isolate (Frankfurt 1), the virus was serially passaged 10 times in 4-week-old F344 rats, resulting in the development of a strain that replicated more efficiently than the original virus. 41 Furthermore, adult F344 rats (6 months old) showed more severe acute lung injury than that of young rats after infection with this new F-ratX strain. 41 An increase in the production of proinflammatory cytokines—particularly, IL-6—was seen in adult rats compared with young rats. These proinflammatory cytokines may be produced by leukocytes and macrophages that infiltrate the injured lungs, leading to enhanced inflammatory reactions. Analysis of the nucleotide sequence encoding the relevant S1 domains of the passaged virus identified a missense mutation in the receptor-binding domain. We found that this mutation is responsible for the more efficient viral replication seen in these rats. 41 DAD was the major pathologic finding in this rat model. During the early phase of infection (3–7 days after inoculation with the rat-adapted SARS-CoV), there was evidence of extensive edema in the lungs; severe inflammatory infiltration by neutrophils, macrophages, and lymphocytes; and hyaline membrane formation (Figs. 2–6). By day 14 postinfection, intra-alveolar fibrosis was present.
The mouse-passaged Frankfurt 1 isolate of SARS-CoV (F-musX) induced severe respiratory illness in adult mice (6 months old) but not in young mice (4–8 weeks old). 43 The moribund adult mice showed severe pulmonary edema and DAD accompanied by virus replication. These mice also showed excessive acute proinflammatory responses in the lungs after SARS-CoV infection, leading to severe pulmonary edema and DAD.
Pathogenesis of SARS
Human lungs infected with SARS-CoV show evidence of DAD characterized by desquamation of epithelial cells, fibrin and collagen deposition in the alveolar space, hyperplasia of type II pneumocytes, increased infiltration of the interstitium by mononuclear cells, and, in some cases, the presence of multinucleated syncytial cells. 6,10,50 The detection of antigen-positive cells in human tissues is usually related to the duration of illness. In fatal cases of short duration, viral antigens were detected in pneumocytes and, occasionally, macrophages within the intra-alveolar necrotic debris in the alveolar septal walls and, rarely, in bronchial epithelial cells.
In animals infected with SARS-CoV (monkeys, mice, and hamsters), tracheal and bronchial epithelial cells, type 1 pneumocytes, and some scattered macrophages were positive for viral antigens within 5 days of inoculation. 15,39,51,54,56,57 The first step of viral infection requires the binding of viral attachment proteins to specific cell surface receptors and, in some cases, coreceptors. The SARS-CoV spike protein (S protein) mediates infection of receptor-bearing cells. The sites of viral replication in humans and animals coincide with the presence of cells expressing angiotensin-converting enzyme 2 (ACE2), which is the functional receptor for SARS-CoV S protein to which it binds with high affinity. 30 –32
Focal interstitial mononuclear infiltration and edema were detected in the lungs of these animals during the early stages of infection and so may give clues to the early events of SARS infection. After elimination of the virus, the changes associated with alveolar damage were accompanied by proliferation of fibroblasts within the inflammatory foci. The number and size of these foci decreased over time, but a few persisted in the lungs of some animals for several weeks after infection. 54
Ferrets and marmosets both develop moderately severe hepatitis after inoculation with SARS-CoV, and in B6 mice, intranasal SARS-CoV infection spreads to the central nervous system (predominantly, the hippocampus) without any associated histopathologic changes. 13
In vivo studies using laboratory animals suggested that 3 factors—expression of the virus receptor (ACE2), proteases, and host immune response—may be important features of the pathogenesis of SARS-CoV infection.
Role of ACE2 in the Pathogenesis of SARS
Several reports supported the suggestion that ACE2 is a physiologically relevant receptor during infection. Tissue expression of the receptor correlates with the localization of the virus in infected individuals and animals. 22,47,61,62 In addition, the efficiency of infection is correlated with the ability of the ACE2 within each species to support viral replication. A study of ACE2 knockout mice indicated that the enzyme is a crucial receptor for SARS-CoV in vivo 24 and that SARS-CoV and the S protein both reduce ACE2 expression. The cellular distribution of ACE2 therefore contributes to the pathogenesis of the disease. Several groups have established transgenic mice that express human ACE2 in the airways and in other epithelial tissues, under the regulation of a global promoter or as driven by the mouse ACE2 promoter. 40,65,73 They found that the lungs and brains of these mice were the major sites of SARS-CoV replication and that extensive neural infection was probably the cause of death. 45,76 However, these sites of virus receptor expression and viral replication are not sufficient to account for the lethal acute pulmonary lesions seen in adult human cases of SARS-CoV infection.
The ACE2 and angiotensin II type 2 receptors are both known to protect against severe acute lung injury in animal models, 20 whereas ACE, angiotensin I, and AT1 receptors may induce edema, leading to acute lung injury. The renin–angiotensin system has an important role in maintaining blood pressure homeostasis, and ACE2, a homologue of ACE, negatively regulates this system. Animal experiments have shown that binding of the SARS-CoV S protein to ACE2 downregulates its expression, resulting in the blockade of the renin–angiotensin pathway and thus promoting disease pathogenesis by inducing edema and impaired lung function. 24 Loss of ACE2 expression in acute lung injury could lead to leaky pulmonary blood vessels through AT1a receptor stimulation. 20
Role of Proteases in the Pathogenesis of SARS
In the study of SARS-CoV infection in cultured cells, Simmons and coworkers found that SARS-CoV takes a receptor-mediated endosomal pathway for cell invasion. 59 They first observed that cells with ACE2 expressing SARS-CoV S protein undergo syncytial formation (cell–cell fusion) when treated with trypsin but not in a low-pH environment. They showed that SARS-CoV infection was blocked by treatment with endosome- or lysosome-tropic agents, such as NH4Cl or bafilomycine, which inhibits acidification of the endosome. 59 These data suggest that SARS-CoV enters cells via the endosomal pathway and that a low-pH environment in the endosome is critical for virus invasion. They also postulated that certain proteases would activate the fusion activity of the S protein in a low-pH environment in the endosome. Subsequently, they reported that cathepsin L, which is active at low pH, plays a critical role for SARS-CoV entry into cells via the endosomal pathway. 60 Bosch et al reported that the treatment of S protein expressing cells with either trypsin or cathepsin L induced cell-cell fusion. 4 They further showed that purified SARS-CoV S protein was cleaved by either trypsin or cathepsin L. These results collectively suggest that SARS-CoV enters cells via endosomes in a protease-dependent fashion and that cleavage of the S protein is critical. 4,60 We found that cells infected with SARS-CoV developed cell–cell fusion after treatment with not only trypsin but other proteases, such as chymotrypsin and elastase. 38 All these proteases with fusion-inducing activity potentiated SARS-CoV entry from the cell surface; these proteases facilitated infection of SARS-CoV attached to the cell surface receptor ACE2, even when infection via the endosome was blocked by endosome-tropic agents. Most interesting, the infection facilitated by these proteases from the cell surface was 100-fold more efficient than infection via endosomal pathway. 38 These findings suggest that one of these proteases—namely, elastase—might be involved in the exacerbation of SARS pathogenesis, given that elastase is one of the major proteases induced in lung inflammation. We therefore induced elastase in the lung of mice with low-virulent bacterial infection or with lipopolysaccharide administration and then infected them with mouse-adapted SARS-CoV. 1,21 This resulted in an exacerbation of pneumonia caused by SARS-CoV infection, and most of the mice died within a week or so with DAD, a form of damage often associated with SARS patients. From these observations, we suggested that this protease is a key factor in the development of severe lung damage after SARS-CoV infection in mice. 1,21 These studies also opened the possibility of using a protease inhibitor as a therapeutic tool for SARS.
Excessive Inflammatory Reactions to SARS-CoV
To develop a mouse model for SARS, BALB/c and C57BL6 mice were infected with human isolates of SARS-CoV. 13,14,74 When 4- to 8-week-old BALB/c and C57BL6 mice were anesthetized and intranasally inoculated with SARS-CoV, the virus replicated in the lungs but produced no clinical signs of disease and was eliminated from the lungs by days 5 to 7 postinoculation. Roberts et al reported that old BALB/c mice (12–14 months) developed signs of clinical illness (weight loss, ruffled fur, and mild dehydration) coupled with histopathologic evidence of pulmonary disease following infection by human isolates of SARS-CoV. 54 This age-related increase in morbidity seen in these BALB/c mice is similar to observations in human cases during the 2003 SARS outbreak. It is now known that advanced age is independently associated with higher mortality in SARS patients. 3,9 A comparative study of the gene expression profiles in pulmonary samples from young and old mice infected with a human isolate, the Urbani strain, suggested that in older mice, the infection induces an exacerbated host immune response to the virus. 2 In young mice, during peak viral replication, genes related to cellular development, cell growth, and cell cycle were downregulated, and these transcripts returned to basal levels as virus was cleared. However, immune response and cell-to-cell signaling genes were upregulated during virus infection and even after viral clearance in old mice.
Concluding Remarks
Comparative studies of human cases and the various animal models of SARS suggest that pathogen and host factors are both important in the pathogenesis of the disease. Most experimental animals infected with SARS-CoV did not present with the severe pulmonary illness seen in the majority of adult human cases of SARS. Several groups have recently developed rodent-adapted strains of SARS-CoV that cause mild to fatal acute respiratory disease. 43,53 These inexpensive and relatively reproducible animal models have potential for their use in testing vaccines and antiviral agents for the treatment of SARS.
Footnotes
Acknowledgements
We acknowledge the research funding from the Ministry of Health, Labour, and Welfare, Japan (grant in aid for research on emerging and reemerging infectious diseases) and a grant from the Ministry of Education, Culture, Sports Science and Technology (No. 19041073). We also thank our colleagues within the institute but, especially, Drs S. Matsuyama, Y. Ami, and T. Sata for their valuable discussion. We also thank Ms M. Fujino, Ms Y. Sato, and Ms A. Harashima for their technical assistance.
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
The authors declared that they received no financial support for their research and/or authorship of this article.