
Abstract
Growth factors are low molecular peptides active in the stimulation of cell proliferation and in the regulation of embryonic development and cellular differentiation. Significant progress has been made in developing effective strategies to treat human malignancies with new chemical compounds based on a rationale directed against various components of signaling pathways. Many of these drugs target a growth factor receptor—for instance, in the form of monoclonal antibodies or inhibitors of tyrosine kinases, such as monoclonal antibodies against epidermal growth factor receptors used in treating certain types of breast cancer. Imatinib mesylate [Gleevec]) is an excellent example of mediators of signal transduction, such as tyrosine kinases. Growth factors proper are used to ameliorate various and sometimes fatal side effects of cytotoxic and/or myelosuppressive chemotherapy. Basic characteristics of several growth families are discussed with therapeutic modalities based on growth factor activity or, more often, inhibition of such activity.
Keywords
Growth factors are low molecular peptides active not only as promoters of cell proliferation but also as regulators of embryonic development and cellular differentiation throughout the life of an organism. Unlike hormones, which are released by specialized endocrine organs and target specific organs by binding to specific cellular or nuclear receptors, growth factors are produced by virtually all cells, and they target specific receptors expressed on numerous cells. High-affinity binding of growth factors to specific cell membrane receptors (and, in a few instances, nuclear receptors) initiates activation of numerous and complex signal transduction pathways leading to gene transcription and the modulation of cellular functions. Because of their role as major regulators of cell growth and proliferation, growth factors, through activation of signaling pathways mediators of the various signal transduction pathways, are also important players as promoters of tumor growth. It then follows that molecules acting as antagonists or inhibitors of growth factors and signal transduction mediators could be utilized as specific biological chemotherapeutic agents.
Although numerous biological activities of growth factors were first identified in live animals and cell lines derived from animal tissues, their role in specific tumor growth and progression has been better characterized in human tumors. This, with the need to develop more effective strategies to treat human malignancies, led to the development and clinical use of new chemical compounds based on a rationale directed against various components of signaling pathways. Many of these drugs target a growth factor receptor (eg, in the form of monoclonal antibodies) or are antagonists of mediators of signal transduction, such as tyrosine kinases. Growth factor receptors do not need their usual ligand or a ligand at all to be activated. For example, a ligand produced by a cell leads to autocrine stimulation of cell membrane receptors expressed by the same cell. Intracrine stimulation of even deficient receptors by activation of intracellular components of the receptor itself or more downstream mediators of signal transduction is also a potent effector of growth factor activity. 203,233 It is for these reasons that the drugs in clinical use today were developed toward the more downstream molecules and that they exhibit a great degree of specificity and, so far, apparently less toxicity. Growth factors proper (or cytokines) are used to ameliorate various and sometimes fatal side effects of cytotoxic and/or myelosuppressive chemotherapy where stimulation of hematopoietic or epidermal cell proliferation is desirable.
The roles of growth factors and their signal transduction systems in individual tumor types in domestic animal species have been studied less and are therefore not as well understood in such roles as those in rodent and human systems. The aim of this review is (1) to compare biological activities of several major families of growth factors, with special emphasis on their activity and expression patterns in tumors, (2) to discuss their application, and (3) to review the use of specific inhibitors of growth factors and their signal transduction pathways in clinical oncology.
Epidermal Growth Factor Family
Epidermal growth factor (EGF), a small polypeptide of molecular weight about 6 kDa, was serendipitously discovered by Stanley Cohen, who was attempting to extract nerve growth factor from salivary glands of male mice (Table 1). His preparation stimulated premature opening of eyelids in newborn mice. 41 Upon its isolation, the stimulating polypeptide was named EGF. EGF has been the most studied and best understood growth factor and serves with its receptor system as a paradigm of a polypeptide growth factor and its signaling system. 26 EGF is expressed almost exclusively in normal adult tissue. Transforming growth factor α (TGFα) is its analog expressed mostly but not exclusively in embryonic tissues (where it participates in their prenatal development) and in a variety of tumors (Table 1). 52 It has a 30–40% amino acid sequence homology to EGF, and it binds to the EGF receptor (EGF-R) with the same affinity. TGFα induces an almost identical range of activities as EGF. 90,158 In addition to the small 6-kDa form, several larger forms of TGFα are biologically active either in their extracellular form or in their transmembrane form. 55 Other members of the EGF family have properties similar to but also distinct from EGF and are not discussed here for lack of space. All the mature peptides show conservation of 6 cysteine residues that form 3 intramolecular disulfide bonds, which then fold the proteins into 3 loops. It is this arrangement of cysteines that defines a peptide as a member of the EGF family. 52 EGF-like repeats are found in many extracellular and cell surface proteins. 41,173 The members of the EGF family bind to a family of glycosylated 170-kDa cell surface erbB-type receptors. 41,42 These receptors have tyrosine kinase activity localized in the intracytoplasmic domain. Upon ligand binding, these receptors form homo- and heterodimers. This leads to activation of tyrosine kinase domains and autophosphorylation of specific tyrosine residues in the cytoplasmic portion of the receptor. 22,222,223 The transmembranous and intracytoplasmic portions are homologous with c-erbB oncogene, one of the first discovered oncogenes. 61 Overproduction of wild type EGF-Rs in some human breast cancers results in cell proliferation, even at low levels of EGF. Monoclonal antibodies to EGF-Rs are used therapeutically in these patients (see below). A constitutively active mutant of HER2, a molecule closely related to the EGF-R, enables uncontrolled proliferation of cancer cells, even in the absence of a ligand. A 185-kDa transmembrane glycoprotein is the product of the HER2/neu gene, 220 and it is overexpressed in up to 30% of human breast cancer cases 15 and in many canine 6,77 and feline mammary carcinomas. 168 A point mutation in the neu gene identified in peripheral nerve sheath tumors of several species of domestic animals raises the question of whether mutations in the neu gene can serve as a useful diagnostic marker. 238
Growth Factors: Their Receptors and Occurrence in Tumors a
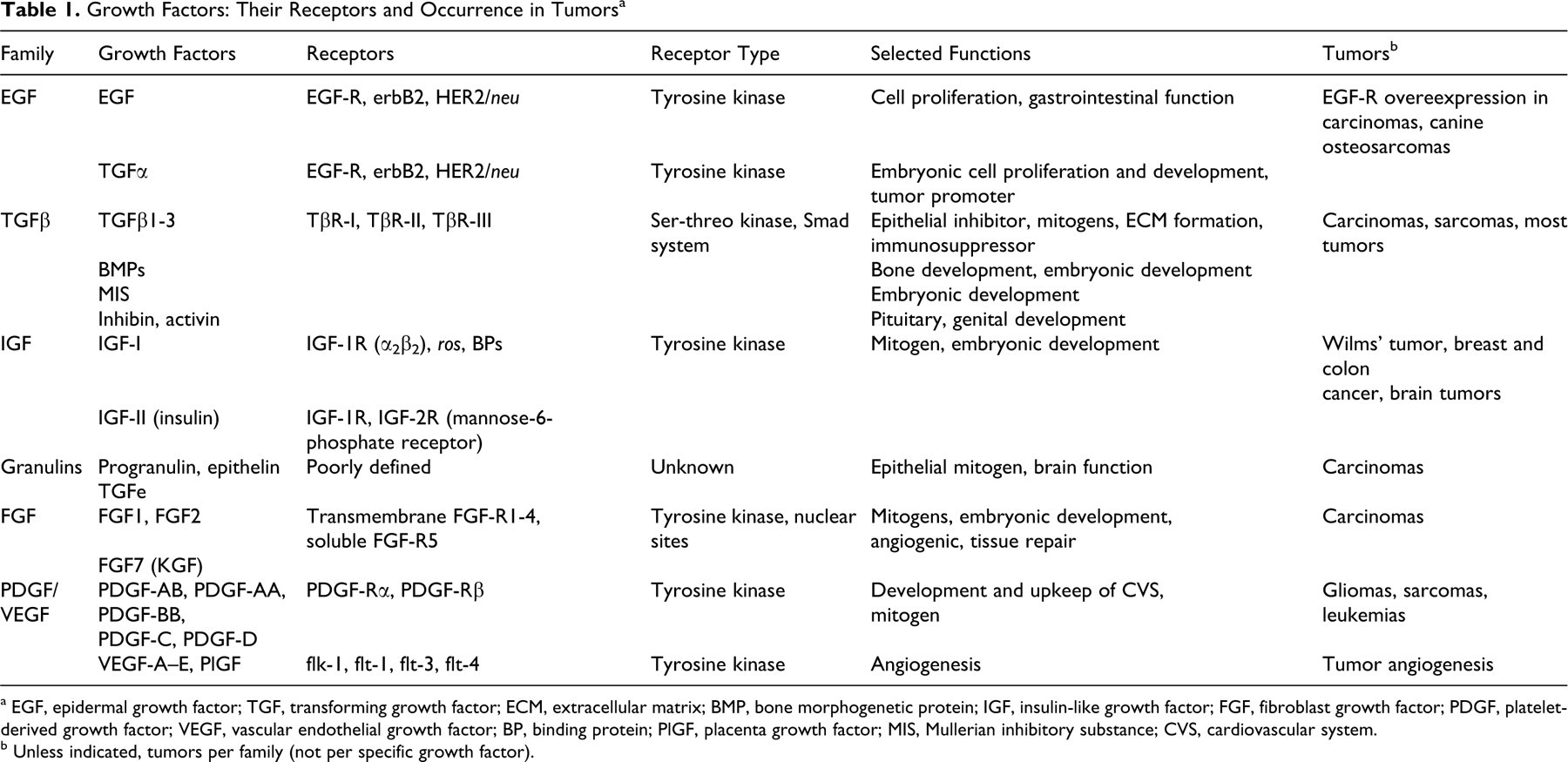
a EGF, epidermal growth factor; TGF, transforming growth factor; ECM, extracellular matrix; BMP, bone morphogenetic protein; IGF, insulin-like growth factor; FGF, fibroblast growth factor; PDGF, platelet-derived growth factor; VEGF, vascular endothelial growth factor; BP, binding protein; PlGF, placenta growth factor; MIS, Mullerian inhibitory substance; CVS, cardiovascular system.
b Unless indicated, tumors per family (not per specific growth factor).
Both EGF and TGFα are mitogens for epithelial and mesenchymal cells; they promote angiogenesis 225 and bone resorption in vitro, 112 as well as precocious eyelid opening in newborn mice, just to name a few activities (Table 1). Although TGFα is primarily expressed in embryonic and tumor tissues, it is synthesized in normal cells. For example, TGFα, synthesized by keratinocytes, stimulates keratinocyte migration. It is also overexpressed in psoriatic epidermis. TGFα is produced by respiratory and gastrointestinal (GI) epithelial cells, macrophages, and pituitary cells (where it stimulates release of prolactin). TGFα is expressed at higher level in the GI tract than EGF is. 39 TGFα synthesis increases in acute injury, and TGFα contributes to repair of injury. It stimulates proliferation of the gastric mucosa and production of gastric mucin, and it inhibits gastric acid secretion. 194,231 It is overproduced in patients with Ménétrier disease, a premalignant gastric condition. 269 Japanese patients with gastric carcinoma have a poor prognosis if their tumors have high expression of EGF, TGFα, or EGF-R. 269 TGFα may act as a tumor promoter in colonic and pancreatic cancer. 82,93
Unlike EGF, TGFα is rhythmically secreted by the hypothalamic suprachiasmatic nucleus, and it plays a role in the circadian rhythm. Its level is high in nocturnal rodents during the day and low at night. 137
Transgenic mice expressing TGFα develop epithelial hyperplasia and metaplasia of the pancreas and GI tract and, after long latency, carcinomas of the mammary gland and liver but exhibit only minor skin aberrations. 118,161,219 Knockout TGFα mice (otherwise healthy and fertile) show only minor hair follicle and ocular abnormalities, 153,155 although later studies indicated a wider range of abnormalities, such as ependymal disruption 252 and GI problems. 65 These findings indicate that redundancy within the EGF family (and the presence of other, unrelated growth factors) allows substitution of one of the other EGF family members when TGFα is missing. EGF-R null mice not only have severely disorganized hair follicles with impaired differentiation but also develop systemic inflammatory disease leading to early death. 94
The overexpression of EGF-Rs and EGF-R-related receptors in many carcinomas indicates that TGFα (or other members of the EGF family) plays a major role in their proliferation. For example, overexpression of erbB-2 was found in spontaneously occurring canine mammary carcinomas 63 and osteosarcomas 72 and in experimentally induced adenocarcinomas of the esophagus. 131 Recent advances in the development and clinical use of monoclonal antibodies as therapeutic agents against specific human malignancies, especially against human colon and breast cancer expressing high levels of EGF-Rs, support this notion.
Amgen’s Vectibix (panitumumab; Thousand Oaks, CA), a monoclonal antibody binding to human EGF-R, is used as a single agent for the treatment of patients with metastatic colorectal cancer expressing EGF-Rs (Table 2). It is interesting (in view of the predominance of skin problems in TGFα transgenic mice) that many patients develop dermatological problems, such as dermatitis acneiform, pruritus, erythema, rash, skin exfoliation, paronychia, dry skin, and skin fissures. This is related to Vectibix’s blockade of EGF binding and subsequent inhibition of EGF-R-mediated signaling pathways. 4
Biologicals Targeted at Growth Factors or Their Receptors a
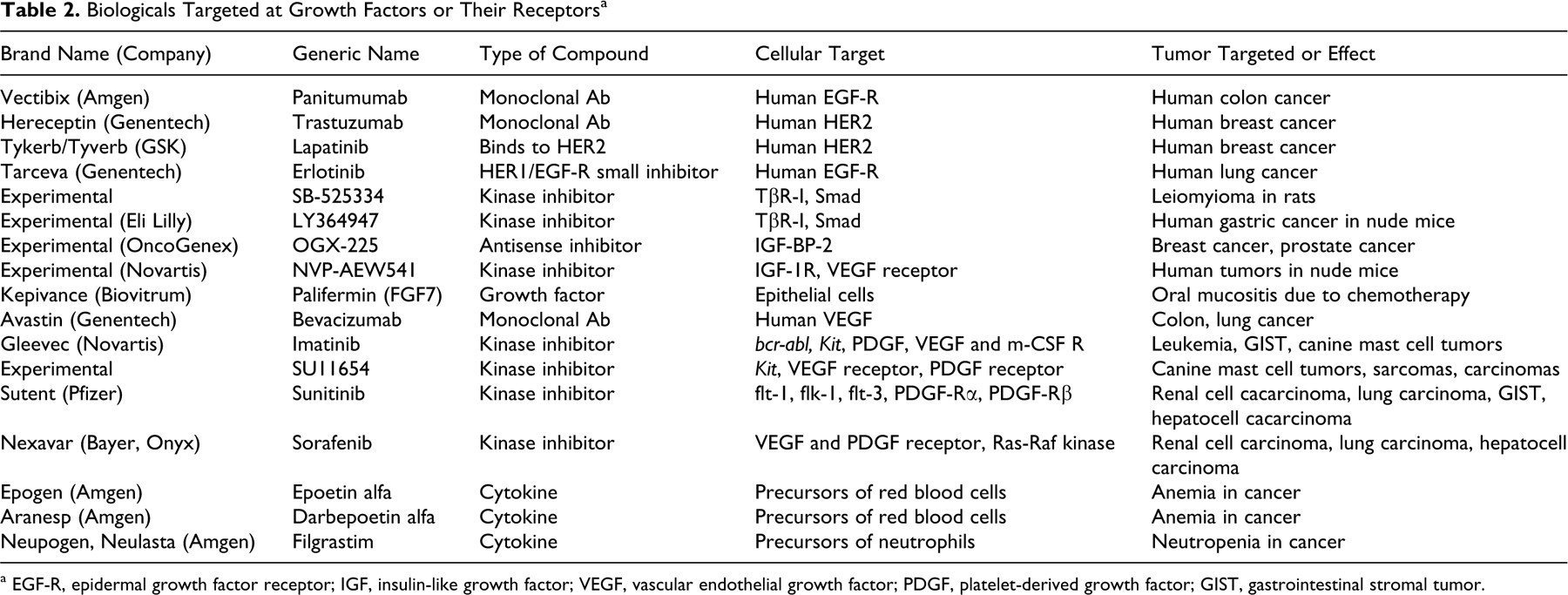
a EGF-R, epidermal growth factor receptor; IGF, insulin-like growth factor; VEGF, vascular endothelial growth factor; PDGF, platelet-derived growth factor; GIST, gastrointestinal stromal tumor.
Genentech’s Herceptin (trastuzumab; South San Francisco, CA) is an antibody that specifically targets HER2 (also referred to as HER2/neu), an EGF-R-related protein present on the cells of some breast cancers (Table 2). About 30% of breast cancers express very high levels of HER2, and Herceptin appears to be effective in this group of women. The level of HER2 within a tumor is first determined using immunohistochemistry. Herceptin is utilized for adjuvant treatment of HER2-overexpressing node-positive or node-negative (ER/PR-negative or with one high-risk feature) breast cancer in combination with standard chemotherapy. Herceptin alone is also used to treat women whose metastatic breast cancers have high levels of HER2/neu and that become resistant to standard hormone therapy and chemotherapy drugs. The combination of Herceptin with paclitaxel, a mitotic inhibitor originally sold as Taxol, is also used for the first-line treatment of HER2-overexpressing metastatic breast cancer. 80
Tykerb/Tyverb (lapatinib) is an oral medication from GlaxoSmithKline (Alhambra, CA) that targets HER2 differently than Herceptin (Table 2). It inhibits receptor autophosphorylation and activation by binding to the adenosine triphosphate–binding pocket of the EGFR/HER2 protein kinase domain. 176 It can be effective in women with advanced or metastatic breast cancer disease resistant to Herceptin but who have high levels of HER2 and who have failed treatment with an anthracycline, a taxane, and Herceptin. 84
Tarceva (erlotinib), also manufactured by Genentech, is a HER1/EGFR small inhibitor used in combination with paclitaxel for the treatment of patients with locally advanced or metastatic non–small cell lung cancer after failure of standard chemotherapy regimen. It is also used in combination with conventional chemotherapy for the treatment of advanced pancreatic cancer (Table 2). 80
It remains to be seen whether similar monoclonal antibodies and receptor inhibitors aimed at components of growth factors signaling pathways could be developed to treat or at least control certain fairly common animal malignancies, such as mammary tumors in dogs.
Transforming Growth Factor β Family
Transforming growth factor β (TGFβ) is a large family consisting of numerous differentiation and development modulating proteins that can be classified into at least 4 groups (Table 1): The Mullerian inhibitory substance (MIS) group regulates Mullerian duct regression in male embryos. The inhibin/activin group includes inhibin (which blocks follicle-stimulating hormone release by pituitary cells) and activin (which stimulates follicle-stimulating hormone release by pituitary cells). They are important in early development of the genital tract. Vg-related proteins regulate primarily embryonic development and cell differentiation. This group includes bone morphogenetic proteins (BMPs; which regulate bone development), dorsalin (which regulates neural tube differentiation), growth differentiation factors, decapentaplegic transcript (which regulates dorsal-ventral patterning in Drosophila), and VgI gene (inducer of mesoderm development from ectoderm in Xenopus). TGFβ family contains 3 isoforms: TGFβ1–3.
All TGFβ-related proteins originated from a common ancestral gene, and they share homologous positioning of least 7 cysteine residues forming a cysteine knot.
159
Each protein is encoded as a large precursor and (with the exception of MIS) is processed to a C-terminal monomer of 100 to 134 amino acids.
81
Active TGFβ is released from latent TGFβ by activation with thrombospondin-1.
226
In general these proteins are not just mitogens: They play crucial roles in embryonic development, morphogenesis, bifunctional regulation of cell proliferation, immune system, tissue repair, and hematopoiesis, to name at least some processes.
159,164,186,211
Because this family is so large in number and so complex in its functions, I discuss only TGFβ1–3 (primarily TGFβ1) and BMPs. The TGFβ1-3 isoforms are 60–80% homologous to each other, and there is an even greater interspecies sequence homology for each isoform. They are homodimers with about 25 kDa and several disulfide bonds. 134,180 TGFβ1 is primarily expressed in endothelial, hematopoietic, and connective tissues cells. 159 TGFβ1 stimulates the proliferation of mesenchymal cells, 37,210 but it is a potent inhibitor of epithelial and endothelial cell proliferation 37,99,243,253 and a potent immunosuppressant. 164 Because it is a promoter of apoptosis and it participates in cell cycle arrest in G1 phase, 202 it acts as a tumor suppressor. 186 TGFβ1 is also a strong fibrogenic agent: 212 It induces the production of collagen and other components of extracellular matrix; it downregulates the expression of matrix metalloproteinases; and it upregulates the synthesis of protease inhibitors. 180 Because of these abilities TGFβ1 promotes stromal proliferation and thus cancer growth. TGFβ1 is the most common and most powerful isoform (and the most studied as well). TGFβ2 is synthesized at higher levels in epithelial cells and neurons, and TGFβ3 is produced mostly in mesenchymal cells. 159
TGFβ1 null mice develop postnatal diffuse and lethal inflammation. This condition became an excellent model for autoimmune diseases, and it underlies the important role of selective immunosuppression in healthy organisms. 7,146 The fulminant autoinflammatory disease also precludes the use of TGFβ1 inhibitors to counteract TGFβ1-induced immunosuppression in cancer patients where a resulting widespread inflammation would have devastating consequences. However, mice with only one copy of the gene for TGFβ1 are normal unless they encounter carcinogenic stimuli, and then they develop lung and liver tumors. 244 Similarly, mice expressing only one copy of the gene for Smad4 or Smad3 (mediators of TGFβ1 signal transduction) develop colon cancer under certain conditions at a higher rate than that of normal mice. 213,242,278
Overexpression of TGFβ1 in transgenic mice leads to widespread organ fibrosis, including pancreatitis, hepatic fibrosis, and impaired lung development leading to fetal lethality owing to a block in branching morphogenesis. 141
The TGFβ receptor and signal transduction system is complex and differs in many ways from receptor systems of other growth factors. 138,186 Most likely, this has evolved because of the multifunctionality of these growth factors where a specific cell function (eg, cell proliferation), depending on conditions, is either stimulated or inhibited by TGFβ. 211 Three classes of TGFβ receptors have been identified: TβR-I, TβR-II, and TβR-III. 34,75,149,150 TβR-III is the most abundant receptor; it is a cell surface proteoglycan called β-glycan. 151 It binds and concentrates TGFβ near the cell surface and transfers it to TβR-I and TβR-II, which are signaling receptors. TβR-I and TβR-II possess an extracellular binding domain, a transmembrane domain, and a cytoplasmic serine-threonine protein kinase domain (rather than a tyrosine kinase domain found in the intracytoplasmic domain of the majority of growth factor receptors). Phosphorylation of the serine-threonine residues leads to phosphorylation and activation of 3 related groups of downstream transcription factors called Smads. Smads are evolutionarily very conserved in their amino acid sequence among a variety of species. 138,187 Smad2 and Smad3 propagate the TGFβ-initiated signal to the nucleus where they associate with Smad4. This step is required for the formation of transcriptional complexes. 186 Although experimental evidence supports the notion that these signal transduction pathways would provide many potential targets for biological chemotherapy, translation into clinical practice has so far not been successful. During carcinogenesis, a long multistep process, most transformed and malignant cells become resistant to TGFβ-induced growth inhibition. For example, this happens in practically all human pancreatic and most human colon cancers. 88,258 Most often this is due to a mutation or loss in expression in one or more genes encoding for components of the TGFβ signaling cascade. Advanced tumors often secrete increased amounts of TGFβ, which greatly contributes to immunosuppression seen in a majority of cancer patients and to metastatic spread as TGFβ gains the ability to stimulate proliferation of cancer and stromal cells alike. 186 In the case of metastases, TGFβ is particularly effective as it stimulates production of extracellular matrix proteins (eg, collagens and fibronectin) and inhibits the production of matrix metalloproteinases (eg, collagenases), which enable the spread of metastatic cells. 19 High levels of serum TGFβ1 were observed in patients with persistent and spreading lung cancer. 51 During human colon cancer progression, TGFβ1 becomes a stimulator of growth and invasion. 109,186 TβR-II mutations lead to inactivation of TGFβ-induced growth inhibition 186,259 and have been found in many human cancers. 157,174 Smad4 mutations are implicated in pathogenesis of pancreatic and colonic cancers. 91,248 Smad3 expression in certain cancer cell lines can be manipulated both ways: to suppress tumor growth in nude mice or to promote aggressive tumor growth. 213 SB-525334, an inhibitor of TGFβ-induced serine-threonine kinase and Smad phosphorylation, inhibited the incidence and growth of leiomyomas in Eker rats (which have a defect in one of the tumor suppressor genes) but increased their incidence of renal carcinomas (Table 2). 140 Studies like this call for caution when developing antitumor strategies targeting the TGFβ system.
Recently, the success in inhibition of TβR-1 with a small molecule inhibitor (LY364947) incorporated into nanocarriers delivered into the tumor during experimental therapy in nude mice carrying human gastric carcinoma shows a new creative approach that deserves further exploration (Table 2). 124
BMPs are involved in the development of the musculoskeletal system and in the differentiation of its components, such as bone, tendon, and cartilage (Table 1). 256 BMP7 null mice die shortly after birth with numerous developmental bone and cartilage defects, polycystic kidney, and malformed eyes. 116 Both BMP7 and BMP2 are excellent inducers of bone formation in vivo, and the hope is that they would provide an effective treatment of poorly healing fractures. 83,114 BMP2 accelerated the repair of delayed union or nonunion bone healing in dogs. 169 Recombinant human BMP7 has been approved for clinical use in people in nonunion fractures in Europe and in Australia. 79 However, the potential of BMPs in treatment of pathological fractures occurring in metastatic disease either in people or animals has not been utilized as of now.
Insulin-like Growth Factors
IGF-I and IGF-II are the only 2 known members of insulin-like growth (IGF) family (Table 1). Because they are closely related to insulin, all 3 are sometimes grouped together. 209 They evolved from an ancestral insulin-like gene by duplication. 132 IGFs are primarily synthesized in the liver. They retain C peptide (which is cleaved off in insulin) and have an extended carboxy terminus. 235 I discuss mostly IGF-I because the biological role of IGF-II is unclear even after many years of intensive research. IGF-I is a small peptide (7 kDa) with 3 disulfide bonds. Hormones regulate the production of IGF-I. The majority of IGF-I is synthesized in the liver under the control of growth hormone. 167 Parathyroid hormone regulates the production of IGF-I in bone. 268 Sex steroids regulate the activity of IGF-I. 166
Whereas insulin is present in blood at picomolar concentrations and has a rapid clearance, IGF-I circulates at nanomolar concentrations. It can bind to any of the 6 types of IGF-binding proteins (IGF-BPs). 122,170 These binding proteins are synthesized in the liver, and they modulate IGF activity by limiting IGF-I access to tissues and receptors. IGF-BP-3 binds more than 95% of serum IGF, and so it is the main regulator of bioavailability of IGF-I. IGF-I upregulates the level of IGF-BP-3. 10,14 The affinity can be reduced by protease cleavage, phosphorylation of the binding protein, or binding of the binding protein to cell surfaces. This would then enhance availability of IGFs. IGF-BP-2 is the second-most abundant IGF binding protein in serum. 263
IGF-I is involved in many functions in many organs. It is a mitogen for epithelial and mesenchymal cells, and it acts as an antiapoptotic agent. 121 It is important in embryonic development and in renewing epithelial cell populations of organs (liver, breast, colon, prostate). It exhibits autocrine activity in several normal cell types (eg, smooth muscle cells in the media of blood vessels, striated muscle cells) and cancer cells. 2,117 It also stimulates collagen synthesis, induces oligodendrocyte development, and supports growth of astrocytoma and meningioma cells in vitro. 195 IGF-I can also act like as a weak insulin, especially in vitro. IGF-BP-1 level is inversely related to insulin level. IGF-BP-1 inhibits the hypoglycemic action of IGF-I, which is about 6% of the hypoglycemic action of insulin. IGF-I has the same metabolic effects as insulin, but it has to be used at higher levels to achieve the same effect.
IGF-I (also IGF-II) has been found in several human tumors: Wilms' tumors, breast and colon cancer, 251 brain tumors, 266 and in cell cultures derived from astrocytomas and meningiomas. 195 The plasma IGF-I level is thus positively related to cancer risk. 190,266 Population studies in people indicate a relationship between circulating levels of IGF-I and IGF-BP-3 and the risk of some common cancers (breast, colon, prostate). IGF-BP-3 was found to have opposite effects on different tumor cells; it potentiates IGF mitogenic effect in some 43 but not in other breast cancer cell lines. 208 Increased levels of another IGF binding protein, IGF-BP-2, have been found in common types of human cancers, such as those of the ovary, colon, lung, and prostate. 40,66,125,142,148 IGF-BP-2 activates the phosphatidylinositol 3-kinase/Akt pathway and regulates PTEN suppressor gene. 148,190 So et al have linked increasing levels of IGF-BP-2 with malignant transformation of human breast epithelial cells, and they found low levels in benign breast tissue. 232 The IGF-BP-2 inhibitor OGX-225 inhibited the expression of IGF-BP-2 in certain cancer cells, with inhibition of cancer cell proliferation (Table 2). These results make OGX-225 an attractive candidate as a therapeutic option in IGF-BP-2-expressing breast cancer. 232
Similarly, IGF and the components of the IGF system have been found overexpressed in animal tumors as well. Normal testis expresses IGF-I, IGF-II, IGF receptors, and IGF binding proteins. 28,275 The expression of the IGF system in canine testicular tumors depends on the tumor type. Whereas low expression of IGF-I and IGF-BP-1 was observed in Sertoli cell tumors and seminomas, the expression of IGF-BP-4 and IGF receptors was upregulated in Leydig cell and mixed tumors. 191 The increase in IGF-I synthesis in canine mammary tumors is linked with the production of growth hormone, and its effects are likely influenced by local levels of estrogen. 171,200 Transgenic mice overexpressing IGF-I developed mammary tumors after a period of increased ductal proliferation. These mice were also more susceptible to development of such tumors after exposure to a mammary carcinogen. 54 It is likely that the IGF-I system contributes to invasiveness and metastatic spread in human and canine osteosarcomas. Cell lines derived from human and canine osteosarcomas overexpress IGF-I receptors and synthesize IGF-BP-4 and IGF-BP-5. The addition of IGF-I stimulated tumor cell proliferation and the invasion of these cells through Matrigel. Some of these canine cell lines were tumorigenic and metastatic in athymic mice. 154
IGF-I receptor is a tyrosine kinase that bears partial homology with insulin receptor and with the extracellular portion of EGF-R. It also has homology with c-ros oncogene. The mature receptor is a heterotetramer α2β2 of 350 kDa 33,145 and specializes in cell growth activities. The related insulin receptor transduces various metabolic pathways. 255,257 Although IGF-II exhibits its mitogenic activity through IGF-I receptor, it binds to its exclusive IGF-2R (which functions as a receptor for mannose-6-phosphate). 209
Inhibitors of IGF-I receptor and its downstream signaling pathways are currently being tested as agents in cancer therapy. 189 Of these, NVP-AEW541, a selective inhibitor of the IGF-I receptor kinase manufactured by Novartis (Basel, Switzerland), appears to be the most promising agent. It inhibited the soft agar growth of several human tumor cell lines, and it significantly impaired growth of human tumors implanted into nude mice (Table 2). 78,172,245 Interestingly, this inhibitor is also apoptotic and antiangiogenic. It acts through the downregulation of mRNA for vascular endothelial growth factor. 172,245 In combination with an inhibitor of the EGF-R, NVP-AEW541 leads to synergistic carcinoma growth inhibition. 22,47 In another study, NVP-AEW541 was shown to have a synergistic effect with Herceptin in inhibition of the growth of human breast cancer cells. 68
An exhaustive review by Samani et al provides additional information on the often-conflicting data and evidence on the relationship between cancer and levels of components of the IGF system (this includes IGFs, IGF binding proteins, IGF receptors, and signaling pathways). This review also describes roles for the IGF system in tumor progression and metastasis either in transgenic mice or in rodents with implanted malignant tumors. 218
Granulins
Granulins are a promising though far-less-studied group of growth promoting proteins (Table 1). Originally described as transforming growth factor e 92,185 and epithelins 229 for their presence in epithelial cells and their autocrine ability to stimulate their proliferation, they were also identified in granules of leukocytes and therefore named granulins. 11 These proteins are widely expressed and play a role in tissue repair, host defense, carcinogenesis, and brain function. 9,45,96,98,152,276 They were originally identified as small (6 kDa) peptides rich in cysteines and disulfide bonds. 11,229 The presence of many cysteine residues makes production of recombinant granulins more difficult. 250 They are ubiquitous and expressed in plants, nematodes, fish, and mammals. 12 In mammals they are synthesized as larger proteins called progranulins, granulin/epithelin precursor, PC-derived growth factor, or acrogranin. 8,17,18,193 This larger form is expressed constitutively in epithelial cells covering skin and lining the GI and reproductive tracts, neurons in the brain, and immune cells. 50 Progranulin also modulates embryonic development of epithelial cells. 56 Increases in progranulin levels in transformed cell lines lead to progression into highly aggressive tumor cell lines, where progranulin stimulates cell proliferation and invasion. 96,97,162
The granulin receptor has evaded full characterization in part because of its presence in low numbers in cell membranes 46,184 and because of an unknown mode of signal transduction and the lack of widely used transgenic model. These two last obstacles also impede progress in development of drugs targeting components of granulin signal transduction pathways. More recently, Cheung et al found that granulin stimulates proliferation of hepatocellular carcinoma cells and formation of metastases. 35 This group has shown that an antibody to granulin inhibited the growth of hepatocellular xenografts in nude mice. 107 These findings should further stimulate research into the potential of antibodies to granulin in treatment of hepatocellular carcinoma and perhaps other malignancies in humans and animals.
Fibroblast Growth Factors
Fibroblast growth factors (FGFs) form a large group of potent mitogens for fibroblasts and epithelial cells and modulators of embryonic development of the mesoderm (Table 1). They are also strong promoters of angiogenesis. Brain, pituitary, retina, eye, and cartilage are good sources of FGFs. 87 The FGF system consists of at least 23 growth factors, 4 high-affinity transmembrane receptors with tyrosine kinase activity (FGFRs 1-4), and 1 soluble receptor (FGFR-5). 32,179,183 The extracellular receptor portion contains 3 immunoglobulin domains. Alternative splicing generates variants of FGFRs, which enhances interaction of FGFs with their receptors. 274 Acidic FGF (FGF1) and basic FGF (FGF2) were the first characterized members of this family of growth factors. 1,87 FGF1 and FGF2 have high affinity for heparin, so they exhibit strong binding to proteoglycan components of the extracellular matrix, where they are often sequestered. During tissue injury, they are released from their extracellular storage sites and are thus activated. These 2 FGFs do not have signal sequence peptide and are not glycosylated. However, they might be released from apoptotic cells and by an alternative protein secretory pathway from intracellular pools. Such release can be triggered by extracellular vesicle shedding, heat shock, injuries, and even normal mechanical stress (eg, plasma membrane distortion during cell movement). 86,228,246 FGF2 release from cells requires the availability of extracellular heparin sulfate proteoglycan in the immediate vicinity of cells. 273 FGFs 11–14 stay intracellular, whereas the remaining FGFs are translated with a signal sequence and glycosylated (ie, they are secreted from cells in the usual way).
Upon receptor binding, the extracellular or intracellular forms of FGFs can translocate to the nucleus, where they bind to nuclear receptors and more directly activate gene transcription. The translocation of intracellular FGFs to the nucleus is one form of intracrine stimulation. 203 FGF2 and FGF3 exist in high molecular weight (HMW) forms that contain a nuclear localization signal. 133,197 Several studies have demonstrated that HMW FGF2 promotes metastatic spread. 178,249 Theoretically, such nuclear receptors should represent good targets for the development of specific antagonists to be used in cancer treatment and in the therapy of other diseases. So far, therapeutic use of FGFs has been limited to keratinocyte growth factor (FGF7).
FGF7, produced exclusively by mesenchymal cells, is a potent mitogen for epithelial cells 70 and, with closely related FGF10, binds to FGF receptor 2 (FGFR2-IIIb) expressed on epithelial cells. 113 FGF7 and its receptor FGFR2-IIIB are expressed in limbal stroma and corneal epithelium, respectively, and thus likely participate in corneal epithelium renewal. 36 Studies on mice transgenic for FGF7 show increased expression of FGF7 early in the embryonic development. This expression results in squamous cell carcinoma of the cornea later in life. However, overexpression of FGF7 in adult transgenic mice leads only to corneal epithelial hyperplasia.
Biovitrum's Kepivance (Stockholm, Sweden; palifermin) is a recombinant form of human FGF7 (Table 2). Kepivance reduces the incidence and duration of severe oral mucositis by helping protect existing epithelial cells that line the mouth and throat from the damage caused by chemotherapy and radiation and by stimulating the growth and development of new epithelial cells. 4 Only 2 growth factors so far have been found to be effective agents of healing of chronic wounds in a clinical setting: FGF7 214 and platelet-derived growth factor. 215
The genes for FGF3, FGF4, and FGF19 are located on human chromosome 11q13.3. This locus is amplified in many carcinomas of head and neck, breast, upper GI tract, and bladder. 128,217,271,272 FGF3 and FGF4 have been characterized as potent transforming proteins. 271 FGF3 was originally identified as an oncogene in murine mammary carcinoma, where it is a target of murine mammary tumor virus (MMTV) integration. 32 Its overexpression was observed in human non–small cell lung carcinoma (with EGF-R). 240 The genes for FGF6 and FGF23 are located on human chromosome 12p13.32. 129 FGF6 is similar to FGF4, and it is involved in malignant transformation. 156 FGF23 has a causative role in tumor osteomalacia. 227 Overexpression of FGF7, FGF8, and FGF17 has been documented in human cancer. 32
FGF receptors have also been implicated in human carcinogenesis. FGFR-1 was originally identified as a chimeric fusion protein with zinger protein in human acute myelogenous leukemia. 267 It is also overexpressed in animal mammary tumors. 261 Mutations in one of the immunoglobulin regions of FGFR-2 and FGFR-3 were identified in human GI and bladder carcinomas and are thought to play a role carcinogenesis. 32,115 Although the FGFR2 IIIC alternative splice variant of the FGFR-2 is primarily expressed in normal mesenchymal cells, its expression is switched on in malignant epithelial (ie, carcinoma) cells, where it drives malignant transformation. 30,32 Progressive deletions of the carboxyl-terminal sequences introduced into alternative splice variants of the FGFR-2 receptor promote its transforming activity through alterations in receptor recycling and stability, thus inducing persistent signaling of the receptor. 31
As members of the so-called stem cell signaling network, FGFs, with the BMP/TGFβ, Wnt, and Notch signaling networks, are key players in embryonic development, tissue repair, and carcinogenesis (eg, of GI tumors 130 ). Perhaps derived from their role in the embryonic development of organs from mesoderm, FGFs participate in developmental epithelial-to-mesenchymal and mesenchymal-to-epithelial transitions, processes instrumental during tumor progression. Specifically, carcinoma invasiveness and metastatic spread are accompanied by these transitions. 30,32,130
Platelet-Derived Growth Factor Family
The members of platelet-derived growth factor (PDGF) family, which includes several PDGF variants and vascular endothelial growth factors (VEGFs), are encoded by genes for 4 PDGF chains (PDGF-A–D) 76,103 and 6 VEGF chains (VEGF-A–E and placenta growth factor). 44,69 The mature growth factors are dimers of disulfide-linked polypeptide chains and are essential for proper embryonic development (primarily of vasculature) and for angiogenesis and vasculature maintenance during the lifetime of an organism. PDGF/VEGF growth factors are a part of superfamily of proteins containing cysteine knots. 165
Angiogenesis
Angiogenesis is a commonly used term that actually describes two similar but nevertheless distinct events: Vasculogenesis means the formation of new blood vessels de novo from differentiation of endothelial cells from mesodermal precursors. It takes place only during embryonic development and leads to formation of a primary vascular plexus. 24 Angiogenesis occurs throughout the prenatal and postnatal life, after vasculogenesis has taken place. 24 New vessels are generated from preexisting vessels during numerous physiological processes (embryonic development, wound repair) and pathological processes (tumorigenesis, diabetic retinopathy). 25 Most growth factors stimulate angiogenesis.
FGFs are potent angiogenic agents, at least in the laboratory. However, during embryonic development, they are involved in mesoderm development rather than vasculogenesis and angiogenesis. That seems to be the domain of VEGF, probably the most specific angiogenic growth factor. After binding to one of its receptors (see below), activation of signaling pathways leads to endothelial cell survival, proliferation, migration, and vascular permeability, all conditions necessary for successful blood vessel formation. 196
PDGF
PDGFs play important roles in repair mechanism in the vascular system, although their physiological roles in the adult are less understood than their participation in various pathological diseases and processes (Table 1). 5
PDGF was purified originally from human platelet α granules, where it is stored (hence its name) as the heterodimer AB. 239 PDGF is expressed and synthesized by endothelial cells, placental trophoblasts, smooth muscle cells, macrophages, and sarcoma cells. PDGF is a 29- to 33-kDa glycoprotein, usually composed of 2 chains (at least in humans), either A or B, which are joined by disulfide bridges. 120,201 The heterogeneity of chain A, thought to be genetic owing to proteolysis during purification or differential glycosylation during synthesis, is responsible for variability in molecular size. Chain A mRNA is expressed mainly in intimal and smooth muscle cells, and the homodimer PDGF-AA is secreted by tumor cell lines. 101
Chain B has molecular weight of about 16 kDa and is encoded by what is known as the c-sis oncogene. 60,260 The designation sis reflects the original identification of this oncogene in a sarcoma of simian origin. It is predominantly expressed in endothelial cells and macrophages in atherosclerotic plaques, but it was also found in mesotheliomas and other tumors. 135,147 Other variants of PDGF include PDGF-C and PDGF-D, identified mostly in tumors. 100
PDGF binds to 2 types of receptors with 5 extracellular immunoglobulin loops and an intracellular tyrosine kinase domain. 5 Their structure is similar to structures of oncogenes and cell membrane receptors for other ligands, such as c-fms, c-kit, and flt3 (a VEGF receptor). Their activation leads to induction of several oncogenes, including c-fos and c-myc. PDGF receptor α (PDGF-Rα) binds homodimer PDGF-AA and monomers PDGF-A, PDGF-B, and PDGF-C. PDGF receptor β (PDGF-Rβ) binds homodimer PDGF-BB and monomers PDFG-B and PDFG-D. The heterodimer PDGF-AB binds to both receptors. 100 Ligand binding induces receptor dimerization and autophosphorylation of tyrosine residues. 100,102 PDGF-B (in PDGF-B+/- mice) and PDGF-Rβ signaling participate in angiogenesis in many organs during embryonic development. PDGF receptors have been studied extensively for their paracrine and autocrine, or rather intracrine, activity; that is, PDGF remains sequestered intracellularly and activates the internal domain of the PDGF receptor. It is for the intracrine activity and activation of oncogenes such as c-ras and c-myc that PDGF receptors may play a crucial role in transformation. 100
PDGF-AB form is present in platelets and is required by fibroblast, smooth muscle, and glial cells for their optimal growth. Its role in wound healing and as a chemotactic agent for fibroblasts and smooth muscle cells, neutrophils, and macrophages has been well documented. It is also involved in blood vessel repair and atherosclerosis, pulmonary fibrosis, and glomerulonephritis. 144,214,239
Experiments with transgenic mice uncovered new pathways for PDGFs. Studies with PDGF-A and PDGF-Rα null mice led to the conclusion that PDGF-A is epithelium derived and is involved in proliferation and spreading of adjacent PDGF-Rα positive mesenchymal cells, and that the cooperation with other growth factor systems (eg, FGFs, BMPs) and signaling molecules (eg, hedgehogs and Wnts) 130 is essential for proper organogenesis of most organ systems. 5 Investigations using PDGF-B and PDGF-Rβ knockout mice showed the importance of PDGF-B and its signaling system in the development of the cardiovascular system. 5
Homodimers PDGF-AA and PDGF-BB are important as mitogens for normal and neoplastic mesenchymal tissues. For example, exogenous PDGF-BB has a modest effect on healing of equine superficial digital flexor tendons. 95 PDGF-BB promotes sprouting of new blood vessels 13 and is mitogenic for smooth muscle cells and pericytes. 64 Relatively high levels of PDGF-BB (as judged from overexpression of the c-sis oncogene) were found in canine osteosarcomas. 135,147 McCarthy et al found that PDGF may act indirectly in osteosarcomas through stimulation of osteoprotegerin, a regulator of osteoclastogenesis, at least in human and mouse osteosarcoma and osteoblastic cell lines. 163 Analysis by Donnem et al indicates correlation between (1) high expression of PDGF-B and PDGF-C and PDGF receptor α and (2) poor prognosis in patients with non–small cell lung cancer, whereas the presence of PDGF-A, PDGF-B, and PDGF-D and PDGF-Rα in tumor stroma (but not in tumor cells) correlated with good prognosis in other patients with the same type of lung cancer. 59 Such complex and at times conflicting results show that we have a long way to go toward a full understanding of the role of PDGFs and their receptors (and, by extension, other growth factors and their receptors) in tumor growth and progression.
Autocrine stimulation involving the PDGF-B/PDGF-Rβ system has been shown to occur during tumorigenesis of gliomas, 48,254 perhaps by activation with TGFβ signaling system (Smad 2/3/4). 21 Upregulation of PDGF-Rα has been documented in human gliomas 21 and canine olidendrogliomas 57 as well, and the level of PDGF-Rα correlated well with the degree of malignancy. 71,105 In dermatofibrosarcoma protuberans, translocations of the PDGF-B gene have been reported. 230 Whereas there is good evidence for autocrine PDGF stimulation in glioblastomas and sarcomas, 5 only the paracrine mode of stimulation has been documented in carcinomas. 103
It is likely that at least in some tumors, there are interactions between the PDGF signaling system and other oncoproteins. For example, bovine papillomavirus E5 oncoprotein (a DNA-type oncoprotein) interacts with PDGF-Rβ in bovine urinary bladder tumors. That the phosphorylated (ie, activated) receptor indeed colocalizes and interacts with E5 in these tumors was shown by confocal microscopy and immunoprecipitation. 20 Katayama et al showed the presence of PDGF and PDGF-Rβ in feline vaccine-associated sarcoma and that they play a role in the proliferation of these cells. The tyrosine kinase inhibitor imatinib mesylate (see below) inhibited PDGF-BB-induced autophosphorylation of the PDGF receptor in the sarcoma cells and thus shows potential as a specific chemotherapeutic agent in this common feline tumor. 126 Other inhibitors or antagonists of PDGF signaling are under development in animal studies or clinical trials in people. 5 Many other tyrosine kinase inhibitors, recently developed and marketed as effective chemotherapeutic agents, target other growth factors and their receptors but demonstrate cross-activity against PDGF.
For the last several years, recombinant human PDGF-BB or becaplermin (Regranex, manufactured by Chiron and formulated by Ethicon) has been used to facilitate healing of difficult chronic wounds in people with severe diabetes. 215,234 However, some rather limited data suggest that long-term application of PDGF might have led to cancer at or near the site of application in a small group of patients who were administered many tubes of Regranex. 3,119 This would represent an example of promotion of already overstimulated cells by a mitogen.
Vascular Endothelial Growth Factors
Vascular endothelial growth factor (VEGF), a distant relative of PDGF, is usually considered an exclusive angiogenic growth factor (Table 1). At least 6 VEGF types have been described: VEGF-A–E and placenta growth factor (PlGF). 69,136 VEGF-A is the most common and most extensively studied isoform, and because it is usually described as VEGF, I use the term here as well. Two groups of VEGF isoforms, VEGFs and PlGF, are the result of alternative splicing of a specific gene. 136 These isoforms differ in their molecular weight and affinity for binding to cell surface heparan sulfate. This binding is important in regulation of bioavailability and activity of individual VEGFs. 205 Plasmin and metalloproteinase-3 regulate VEGF activity by generation of VEGF proteolytic fragments. 108,143 VEGFs are glycosylated homodimers with sequence homology (20%) to A and B chains of PDGF. They contain a core of regularly spaced cysteine residues (cysteine knot motif).
Three VEGF receptors—VEGRF-1 (flt-1), VEGFR-2 (KDR/flk-1), and VEGFR-3 (flt-4)—have been described. They belong to a subfamily of tyrosine kinase receptors within the PDGF receptor class and are characterized by 7 extracellular immunoglobulin-like domains. They are expressed in endothelial cells from the stage of blood island formation onward, throughout embryogenesis. All VEGF isoforms can bind to VEGFR-1 and VEGFR-2. VEGFR-1 and VEGFR-2 are localized to vascular and lymphatic endothelial cells in adult tissues. VEGFR-3 is present mostly in the lymphatic endothelium. However, VEGFR-2 is the main VEGF receptor in endothelial cells. 181,241 VEGF activates endothelial cells, which then secrete matrix-degrading proteinases. This event is necessary for the proper formation of normal blood vessels, but it is also a necessary step during cancer invasion and spread of metastases. 196,236
Although targeting VEGF (or its receptor) would affect both normal cells and cancer cells, it could suppress the proliferation of cancer cells more profoundly because of faster growth of blood vessels in tumors, at least in theory. Moreover, studies on transgenic animals indicate that VEGF acts not only as an angiogenic growth factor but also as an autocrine agent, directly stimulating the proliferation of primary tumor cells and the growth of metastatic lesions. 110,224 It has been known that overexpression of c-myc oncogene in human breast cancer is associated with metastatic disease. 53 In contrast, mammary tumors in MMTV-myc transgenic mice do not metastasize and, in addition, are poorly vascularized. Only when VEGF is expressed in MMTV-myc mice does metastatic disease ensue, characterized by high degree of vascularity and increased synthesis of extracellular matrix proteins, such as type I collagen, fibronection, and tenascin C. 23 This is supported by Kato et al, who observed an increase in plasma and serum VEGF in dogs with mammary tumors. 127 Qiu et al also found VEGF overexpression in spontaneous canine malignant mammary tumors and even more so in their lymph node metastases. 198 Interestingly, the same group showed expression of PTEN to be lower in malignant mammary tumors than in either benign tumors or normal mammary gland and even lower in lymph nodes metastases. 199 PTEN protein is a product of a suppressor gene involved in phosphatase regulation; that is, PTEN counteracts phosphorylation events induced by growth factors, including VEGF. Unfortunately, it was not clear from the above studies whether there was a correlation between low PTEN and high VEGF in individual tumors or metastases. Reddy et al showed that VEGF is capable of inducing differentiation of bone marrow cells transplanted into nude mice carrying Ewing’s sarcoma into vascular smooth muscle cells/pericytes. 206 In another study, the growth of human Ewing’s sarcoma carried in cell culture and in mice was suppressed by tyrosine kinase receptor (including flk-1 and flt-1) inhibitors SU6668 and SU5416 or with anti-VEGF antibody (bevacizumab, see below), presumably as a result of suppression of angiogenesis. 49
VEGF has been identified in many other types of canine neoplasms besides mammary tumors. Intracranial tumors—including malignant glioblastomas, oligodendrogliomas, and benign meningiomas 58,192,216 —express VEGF, as do vascular tumors, 247,270 lymphomas, 265 mastocytomas, 204 and melanomas. 262
It should not be too surprising then that monoclonal antibodies to VEGF or the components of the VEGF signal transduction system would be effective chemotherapeutic agents. Avastin (bevacizumab), manufactured by Genentech, represents an excellent example (Table 2). This compound is a monoclonal anti-VEGF antibody that has been used in combination with classic chemotherapeutic drugs to treat patients with metastatic carcinoma of the colon or rectum; patients with unresectable, locally advanced, recurrent or metastatic nonsquamous non–small cell lung cancer; and patients who have not received chemotherapy for metastatic HER2-negative breast cancer. 80 Active immunization against xenogeneic VEGF (ie, against recombinant human VEGF) in dogs with spontaneous cancer led to decreases in plasma levels of VEGF and decreased tumor microvessel density with an overall 30% tumor response (ie, decrease in tumor volume). Because the dogs exhibited a strong antibody response to xenogeneic, but not canine, VEGF, it is likely that that immune tolerance to canine VEGF was not effectively broken. 123 Caunt et al showed that an antibody to neuropilin-2—a coreceptor for VEGF-C and an isoform expressed mostly in lymphatic endothelial cells—reduces distant mouse mammary carcinoma metastases by blocking VEGF-C binding to its receptor and inhibits its effect on migration of lymphatic endothelial cells. 29 However, Stockmann et al observed that, paradoxically, deletion of VEGF, at least from inflammatory white cells infiltrating tumors, results in accelerated tumor progression. At this point, it is not clear whether this effect is due to deletion of VEGF itself or whether inflammatory cells lacking VEGF are less capable of containing tumor cells. 237
Antiangiogenic Agents
Because tumors require blood vessels for their growth and invasion, targeting tumor angiogenesis has been the holy grail of specific biological chemotherapy. Folkman was the first scientist to develop specific cancer treatment (at least in mice) based on inhibition of angiogenesis within tumors. He used several collagen fragments, endostatin and angiostatin, in his studies on mice bearing tumors. 16,182 These peptides act at least in part through inhibition of VEGF action. 74 There are many angiogenesis inhibitors in clinical use or at least in clinical trials for cancer therapy. Folkman classified them into several groups. 73 The group of so-called total antiangiogenic drugs includes anti-VEGF antibodies (see above), endostatin, and angiostatin. Other agents are only partially antiangiogenic, and they include drugs used for other purposes, such as Herceptin (used to treat women with HER2-positive breast cancer) and Celebrex (celecoxib; used to treat arthritis). Direct angiogenesis inhibitors such as endostatin act to block endothelial cells from responding to endothelial mitogens, whereas indirect agents block a tumor oncogene or a receptor. Avastin, which targets VEGF receptor, belongs to the latter category. Many antiangiogenic agents (eg, interferon β, thrombospondin, angiostatin, and endostatin) are produced endogenously. 73 It is likely that in the future, antiangiogenic drugs will be combined with other anticancer agents to enhance their anticancer effectiveness (see below). The success of the combination of Avastin with traditional chemotherapy in treating cancer is quite promising in this type of approach. 111 More recently, a combination therapy exploiting inhibition of endothelial cells and pericytes by inhibition of signaling of VEGF and PDGF receptors shows potential, at least in mice. 139
Targeting Signal Transduction
Gleevec (imatinib) has been manufactured by Novartis as the first specific biological chemotherapeutic agent (Table 2). 177 It targets bcr-abl fusion protein involved in the development of Philadelphia chromosome–positive chronic myeloid leukemia in people. 27,62 Abl, the product of an oncogene, is a tyrosine kinase requiring cytokine stimulation of a receptor for its activation. Activation of abl leads to dimerization and autophosphorylation of the protein in normal cells. The bcr protein contains a dimerization motif but without kinase activity, and its function is unknown. The Philadelphia chromosome, a marker of human chronic myeloid leukemia, is a result of translocation between chromosome 9 (containing the ABL gene) and chromosome 22 (containing the BCR gene). This translocation leads to production of a novel fusion product containing the dimerization domain of bcr and the kinase domain of abl. Consequently, the fusion protein forms a tetramer in the absence of cytokine, resulting in a constitutively active tyrosine kinase and uncontrolled cell division. Imatinib blocks the activity of the chimeric bcr-abl protein. Because this protein is synthesized predominantly in leukemic cells, normal cells are much less affected, and recipients of imatinib suffer from fewer side effects. Imatinib also blocks the activity of the abnormal Kit protein, the product of the mutated gene in metastatic Kit-positive GI stromal tumor (GIST). In addition, imatinib inhibits the activity of tyrosine kinases associated with PDGF, VEGF, and macrophage colony-stimulating factor (m-CSF) receptors. 106,160,221 A clinical trial evaluating its efficacy in women with persistent ovarian cancer expressing at least one target of imatinib mesylate (Kit, PDGFR-α, or PDGFR-β) is in progress. 221 Hiraga and Nakamura suggested that imatinib mesylate reduces osteoclast activity through inhibition of m-CSF receptor-induced phosphorylation of c-FMS and suppresses the growth of bone metastases in a nude mice model. 106 Whether these results will be applicable to women with metastatic breast carcinoma or to dogs suffering from the same tumor type remains to be seen.
Similar treatment modalities blocking tyrosine kinases have been under investigations in veterinary oncology as well. Kit tyrosine kinase has been implicated in the growth of aggressive mast cell tumors in dogs. Several Kit and other tyrosine kinase inhibitors, including imatinib and SU11654, when given as a single agent or in combination, led to cell cycle arrest and apoptosis of malignant mast cells, sarcomas, and carcinomas. 85,150
Numerous other tyrosine kinase inhibitors are being currently tested on human patients in multicenter phase III clinical trials. To enumerate and describe them all would constitute another extensive review. Just like imatinib, these agents target multiple tyrosine kinase inhibitors and are explored in therapy of several types of malignancies. For example, sunitinib (Sutent, manufactured by Pfizer, New York, NY; Table 2) blocks receptor tyrosine kinases of the split-kinase domain family, such as flt-1, flk-1, flt-3, PDGF-Rα, and PDGF-Rβ. Sunitinib is being evaluated for the treatment of metastatic renal cell carcinoma, 38,104 non–small cell lung cancer, 264 GIST, 207 breast cancer, 175 and hepatocellular carcinoma. 89 Another compound is sorafenib (Nexavar, manufactured by Bayer, Leverkusen, Germany; Table 2), an inhibitor of VEGF and PDGF receptor and the Ras-Raf kinase pathway. It has been found effective in patients with renal cell carcinoma, 38,104 lung cancer, 264 and hepatocellular carcinoma. 89 Side effects associated with this type of therapy are the consequence of the biological activity of these agents. Thromboembolism and hemorrhage have been documented with the use of sunitinib and sorafenib. 67 Sunitinib is also thought to lead to hypertension and renal dysfunction, 277 neovascularization of epiphyseal growth plate, and impaired corpora lutea formation in monkeys. 188
Hematopoietic Cytokines
Several hematopoietic cytokines have been used in human oncology to treat the often deadly side effects of cancer chemotherapy, such as anemia and neutropenia (Table 2).
Epogen (epoetin alfa) and Aranesp (darbepoetin alfa) are recombinant proteins manufactured by Amgen that work in a similar way as the body’s natural erythropoietin—a glycoprotein produced by the kidneys that circulates through the bloodstream to bone marrow, where it stimulates red blood cell production. These drugs are in extensive clinical use in oncology to treat anemia in cancer patients. Interestingly, progression or recurrence of breast, non–small cell lung, head and neck, lymphoid, and cervical cancers in patients receiving high-dose and long-term therapy with recombinant erythropoietin has been reported. In addition, patients have experienced renal failure and cardiovascular and thromboembolic events, which at times have been fatal. 4
Neupogen (filgrastim) is another recombinant cytokine produced by Amgen. It stimulates the production of neutrophils during myelosuppressive chemotherapy-induced neutropenia. However, severe, even fatal, side effects of Neupogen call for caution when administered. Splenic rupture and sickle cell crises have been reported in patients receiving Neupogen; some cases have been fatal. Acute respiratory distress syndrome and allergic reactions have also been reported. 4
Neulasta, also from Amgen is a longer-lasting form of Neupogen. Neulasta requires only one injection per chemotherapy cycle, whereas Neupogen may require daily injections for up to 14 days following chemotherapy. Splenic rupture (including fatal cases), acute respiratory distress syndrome, sickle cell crises, and allergic reactions have been reported. 4
Postscriptum
This article was not meant as, and is not, a comprehensive review of growth factor involvement in carcinogeneis. Because the topic is so vast, with thousands and thousands of exciting studies entered into databases, it was a real challenge what to choose. I am afraid that the inclusion of topics and studies was guided by my personal preferences and research interests. Anyway, the review is supposed to spur interest and perhaps stimulate more effort and research in veterinary cancer medicine, both basic and applied.