
Abstract
Objective
Oxidative stress caused by the pro-inflammatory cytokine interleukin (IL)-1β has been widely investigated for cancer risk. In this study, we focused on the role of IL-1β rs1143634 polymorphism to reveal its impact on cancer development.
Methods
Related studies with fixed inclusion criteria were selected from electronic databases to May 2021. This meta-analysis was performed with odds ratios and 95% confidence intervals. Heterogeneity, publication bias and sensitivity analyses were also conducted. Trial sequential analysis (TSA) and in-silico gene expression analysis were performed.
Results
Forty-four case–control studies involving 18,645 patients with cancer and 22,882 controls were included. We observed a significant association of this single nucleotide polymorphism with overall cancer risk in the codominant model 3 (1.13-fold), recessive model (1.14-fold) and allelic model (1.08-fold). Subgroup analysis revealed that rs1143634 elevated the risk of gastric cancer, breast cancer and multiple myeloma. In addition, Asian and mixed populations and hospital-based controls had a significantly higher risk of cancer development. TSA confirmed our findings.
Conclusion
Our meta-analysis revealed that the presence of IL-1β rs1143634 polymorphism increases the risk of cancer development. Among polymorphism carriers, the Asian population has a higher risk than other ethnic populations.
This meta-analysis was registered retrospectively at INPLASY (https://inplasy.com/, INPLASY2021100044).
Introduction
Currently, cancer is a leading cause of death worldwide. Oxidative stress induced by chronic inflammation plays a vital role in cancer development. Although inflammation is necessary for the immune system to protect the body against foreign infections, the overstimulation of inflammatory cytokines has been identified to be responsible for cancer progression.1–6 Cancer cells often increase the release of cytokines that stimulate the activation of multiple genes involved in cellular migration, proliferation and survival. These cytokines help establish a favorable microenvironment for neoplastic initiation and DNA damage.7,8 Interleukin-1 (IL-1) is a pro-inflammatory cytokine that exerts a wide range of biological actions, and several case–control studies have shown that IL-1 polymorphisms are significantly associated with different cancers . 9
The IL-1β gene encodes IL-1β, which is one of the most potent pro-inflammatory cytokines that initiates and amplifies both acute and chronic inflammation and is involved in various cellular actions, such as proliferation, differentiation and apoptosis. Upon stimulation, blood monocytes and tissue macrophages produce IL-1β proprotein, which is cleaved and activated by caspase 1.10–15 According to genome-wide association studies, patients with three common characteristic polymorphisms of this gene, including rs16944, rs1143627 and rs1143634, are highly susceptible to cancer development.16–18
rs1143634, also known as +3954C>T, is a silent coding sequence polymorphism located in exon 5 of chromosome 2. This single nucleotide polymorphism (SNP) showed a significant association with increased IL-1β release from lipopolysaccharide-induced cells in previous in vitro studies.19–21 Silent SNPs tend to produce truncated proteins that remain inactive or degrade faster than active proteins. This occurs when a silent SNP inactivates the splicing site and causes premature termination of mRNA transcription. 22 In the case of rs1143634, the presence of this polymorphism increases active IL-1β rather than inactive protein.19–21 Excess IL-1β concentrations facilitate a suitable environment for cancer development by increasing the rate of uncontrolled cellular proliferation and differentiation and interfering with apoptosis. Over the past decades, several individual case–control studies on IL-1β rs1143634 polymorphism and cancer susceptibility have been conducted in different ethnic groups. Although some studies reported a significant link between this variant and different cancers, others failed to establish any significant association. 9 In this meta-analysis, we summarized previous studies to investigate the connection between IL-1β rs1143634 polymorphism and cancers and provide comprehensive outcomes.
Materials and methods
Literature search strategy
Multiple authorized electronic databases (PubMed, Google Scholar, CNKI, Web of Science and EMBASE) were comprehensively searched for related literature using specific key terms up to May 2021. The selected key terms included cancer, interleukin-1 beta, rs1143634 (+3954C>T), IL-1β polymorphism and cancer, link between IL-1β rs1143634 and carcinogenesis and IL-1β polymorphism and cancer development in various ethnic populations. Additional studies were extracted from the references and citations of the selected studies and the ‘similar studies’ option of the respected websites. We selected published studies without restricting available languages.
Publication screening
The eligibility of the publications was determined based on the previously selected key terms, and the overall selection process was completed using a protocol designed by the authors. The authors (SJ and MAA) selected the eligible studies containing the related data and organized the extracted data for the meta-analysis by comprehensive screening. The overall study selection protocol was designed as a PRISMA flow diagram 23 using Review Manager (RevMan), Version 5.4 (The Cochrane Collaboration, 2020). The overall process was revised through final screening by another author (MSI).
This meta-analysis was retrospectively registered at INPLASY (https://inplasy.com/, INPLASY2021100044). Because no patients or controls were directly involved in this meta-analysis, patient consent and ethical approval were not necessary.
Inclusion and exclusion criteria
The main inclusion criteria of the selected studies were that they must contain comparative genotypic information and detailed data regarding IL-1β rs1143634 (+3954C>T) polymorphism in both patients with cancer and control populations. If the selected studies contained genotypic data on other SNPs, we only extracted the IL-1β rs1143634 (+3954C>T) data to include in this meta-analysis. We excluded studies without IL-1β rs1143634 genotypic data in patients with cancer as they were not eligible for this study. Publications containing incomplete genotypic data on rs1143634 were also excluded. Studies lacking control population data and those with incomplete information were avoided for further comparison in this meta-analysis.
Extraction and quality assessment of data
The study ID, publication year, country and ethnic background of the study population, cancer type, control type, genotypic method, sample and control size, clinical histories and basic characteristics, genotypic data for the selected SNP, Hardy–Weinberg equilibrium (HWE) p-value and Newcastle–Ottawa Scale (NOS) score were collected from each selected study by the authors. 23 Two authors (SJ and MAA) screened and processed the data using a previously designed protocol, and another author (MSI) reviewed the organized data by conducting the final screening.
Statistical analysis
We performed statistical analysis by comparing the frequency of IL-1β rs1143634 polymorphism among patients with different cancers and control populations to determine the connection between IL-1β rs1143634 variants and cancer development susceptibility. The meta-analysis used hospital-based (HB) and population-based (PB) control populations as the control arms and patients with various cancers carrying the IL-1β rs1143634 polymorphism as the experimental arm. We used Review Manager (RevMan 5.4) to perform the overall statistical data analysis. Estimation of cancer susceptibility was pooled as odds ratios (ORs) with 95% confidence intervals (CIs). Based on heterogeneity, both the fixed-effect model and the random-effects model were used (Q-test). If heterogeneity was significant (p-value <0.10), a random-effect model was applied, and when heterogeneity was not significant, the fixed-effect model (Mantel–Haenszel) was applied.
The Begg & Mazumdar test and Egger’s regression test were carried out to estimate publication biases. Sensitivity analysis was also performed to assess the reliability of the results by excluding individual studies one at a time. Ethnicity-based sub-group analyses (White, Asian, African and mixed) were conducted to analyze the role of IL-1β rs1143634 in patients with cancer among different ethnic populations. Cancer types with less than two studies were sub-grouped into ‘other cancers’ for further subgroup analysis.
We applied seven common genetic models, including the association-allele model (AM: T vs. C), codominant model 1 (COD1: TC vs. CC), codominant model 2 (COD2: TT vs. CC), codominant model 3 (COD3: TT vs. TC), dominant model (DM: TT+ TC vs. CC), recessive model (RM: TT vs. TC + CC) and over-dominant model (OD: TC vs. TT+ CC). TT, TC and CC indicate normal homozygotes, heterozygotes and mutant homozygotes, respectively.
Trial sequential analysis (TSA)
TSA was performed to reduce the random error risk. We first determined the required information size (RIS) and defined the monitoring boundaries by setting the following criteria: 1) 95% CI with a p-value <0.05, 2) 20% relative risk reduction, 3) 80% statistical power and 4) 5% type I error. We used TSA software (version 0.9.5.10 beta) 24 for conducting TSA. The statistical summary (Z values) was plotted on the Z-curve, which showed the TSA boundary. If the cumulative Z-curve crossed the TSA boundary or RIS, this meta-analysis was considered to have achieved a reasonable and sufficient degree of evidence, confirming that no additional studies are required.
In silico gene expression analysis
To evaluate the overall impact of rs1143634 polymorphism on the IL-1β gene expression level, we conducted an important in silico gene expression analysis termed expression quantitative trait loci (eQTL) analysis through the GTEx portal website (http://www.gtexportal.org/). Two skin samples from the GTEx database were analyzed, including sun-exposed skin samples and non-sun exposed skin samples (suprapubic). Sun-exposed skin samples were taken from the lower leg, and non-sun-exposed skin samples were taken from the suprapubic area. The skin samples were obtained as slices with the subcutaneous fat removed, avoiding pubic hair in the suprapubic region.
Results
Selection of the individual studies
Figure 1 outlines the complete study selection process in this meta-analysis. Forty-four studies17,25–67 were selected from 970 primary studies acquired from the searched databases following the eligibility criteria. Comprehensive screening of the titles, abstracts and full texts for each study was conducted to include or exclude the studies. The quality of the studies was determined using the NOS quality assessment score, and low-quality studies (score <6) were excluded. Among the 44 studies, there were 18 on gastric cancer (GC), 8 on lung cancer (NSCLC), 7 on breast cancer (BC), 4 on colorectal cancer (CRC), 3 on prostate cancer (PCa) and 4 on other cancers.

PRISMA flow diagram for study selection.
Study characteristics
The basic demographic information of the 44 selected case–control studies involving 18,645 patients with cancer and 22,882 controls is summarized in Table 1. Among them, 16 studies were from the Asian population, 23 studies were from the White population, 2 studies were from the African population, and the other 5 studies were from mixed populations. One association study recruited both African and White populations for the IL-1β rs1143634 polymorphism. Most studies reported the HWE p-value.

SMA: Sequenom MassARRAY; RT-PCR: reverse transcription-polymerase chain reaction; SSP: sequence-specific amplification; RFLP: restriction fragment length polymorphism; ALM-ASA: adapter-ligation mediated allele-specific amplification; HB: hospital-based; PB: population-based; HWE: Hardy–Weinberg Equilibrium; NOS: Newcastle–Ottawa Scale; GC: gastric cancer; CRC: colorectal cancer; BC: breast cancer; LC: lung cancer; NSCLC: non-small cell lung cancer; SQLC: squamous cell lung cancer; MM: multiple myeloma; PCa: prostate cancer; PNT: pancreatic neuroendocrine tumor; NPC: nasopharyngeal cancer; OS: osteosarcoma, ND: not defined.
Association of IL-1β rs1143634 with cancers
The overall meta-analysis of the total study population showed a significantly elevated risk in patients with cancer carrying the IL-1β rs1143634 variant in three different genetic models, including COD3 (TT vs. TC: OR = 1.13, 95% CI = 1.02–1.25, p = 0.016), RM (TT vs. TC + CC: OR = 1.14, 95% CI = 1.04–1.25, p = 0.006) and AM (T vs. C: OR = 1.08, 95% CI = 1.0–1.17, p = 0.039). According to the ethnicity-based subgroup analysis, the White population did not show any significant link between rs1143634 and cancer risk. In contrast, the Asian population showed a significantly increased risk of cancer among the variant carriers in several genetic models, including COD1 (TC vs. CC: OR = 1.54, 95% CI = 1.12–2.11, p = 0.008), DM (TT + TC vs. CC: OR = 1.54, 95% CI = 1.14–2.09, p = 0.005), OD (TC vs. TT+CC: OR = 1.48, 95% CI = 1.07–2.03, p = 0.017) and AM (T vs. C: OR = 1.50, 95% CI = 1.15–1.95, p = 0.003). In the African population, there was no significant association between cancer risk and the rs1143634 variant. Other studies with mixed populations showed a significant risk in COD2 (TT vs. CC: OR = 1.22, 95% CI = 1.07–1.40, p = 0.004), COD3 (TT vs. TC: OR = 1.22, 95% CI = 1.06–1.40, p = 0.006), RM (TT vs. TC + CC: OR = 1.22, 95% CI = 1.07–1.39, p = 0.004) and AM (T vs. C: OR = 1.05, 95% CI = 1.0–1.11, p = 0.050) genetic models.
In the sub-group analysis of different cancer types, patients with IL-1β rs1143634 polymorphism showed a significant risk of GC in COD1 (TC vs. CC: OR = 1.25, 95% CI = 1.00–1.56, p = 0.048), DM (TT + TC vs. CC: OR = 1.25, 95% CI = 1.01–1.56, p = 0.039), OD (TC vs. TT+CC: OR = 1.25, 95% CI = 1.00–1.55, p = 0.045) and AM (T vs. C: OR = 1.21, 95% CI = 1.00–1.46, p = 0.044) models. For BC, patients with this polymorphism showed a significantly increased cancer risk in two genetic models, including COD2 (TT vs. CC: OR = 1.31, 95% CI = 1.03–1.67, p = 0.029) and RM (TT vs. TC + CC: OR = 1.35, 95% CI = 1.08–1.67, p = 0.008). The carriers of IL-1β rs1143634 polymorphism showed a significant risk for multiple myeloma (MM) in the RM model (TT vs. TC + CC: OR = 2.64, 95% CI = 1.25–5.57, p = 0.011). No significant association was found for the other types of cancers. Sub-group analysis of HB control populations showed a significantly increased risk in two genetic models: DM (TT + TC vs. CC: OR = 1.18, 95% CI = 1.00–1.40, p = 0.049) and AM (T vs. C: OR = 1.17, 95% CI = 1.02–1.35, p = 0.030). PB controls with rs1143634 polymorphism did not show any association with cancer risk. The overall findings were summarized in Table 2 and Figure 2.
Meta-analysis of the association between IL-1β rs1143634 polymorphisms and cancer susceptibility.

OR: odds ratio, 95% CI: 95% confidence interval, COD1: codominant model 1, COD2: codominant model 2, COD3: codominant model 3, DM: dominant model, RM: recessive model, OD: over-dominant model, AM: allelic model; GC: gastric cancer; BC: breast cancer; LC: lung cancer; CRC: colorectal cancer; PCa: prostate cancer; MM: multiple myeloma; HB: hospital-based; PB: population-based. Bold values indicate statistically significant differences (p < 0.05).

Forest plots describing the association between IL-1β rs1143634 (+3954C>T) polymorphism and cancer susceptibility.
Heterogeneity
The Q-test was performed to determine the degree of heterogeneity (Table 2). Heterogeneity was significant in maximum subgroup analysis models (p-value <0.1), and random-effect models were applied for these analyses. All subgroup analyses showed significant heterogeneity (p-value < 0.1), except the subgroup analysis with the mixed population and patients with PCa. The overall analysis with the total study population did not show significant heterogeneity in COD3 (I2 = 19.52) and RM (I2 = 22.46) genetic models.
Sensitivity and publication bias analyses
To confirm the reliability of the outcomes, we performed a sensitivity analysis by the sequential omission of each study. The influence of each included study on the final outcome of this meta-analysis was analyzed, and none of the studies interfered with the pooled ORs. The sensitivity analysis revealed the stability and robustness of this meta-analysis (Table 3).
Sensitivity analysis of the meta-analysis.

COD1: codominant model 1, COD2: codominant model 2, COD3: codominant model 3, DM: dominant model, RM: recessive model, OD: over-dominant model, AM: allelic model.
Publication bias was tested using Egger’s test and Begg & Mazumdar’s test. The funnel plots are shown in Figure 3, and the bias parameters are presented in Table 4. The bias analysis was conducted for overall studies, and no visual asymmetry was found for COD2, COD3 and RM, indicating the absence of publication bias. The rest of the analysis model showed potential publication biases (p-value <0.05).

Funnel plots indicating publication bias.
Outcome of publication bias analysis.
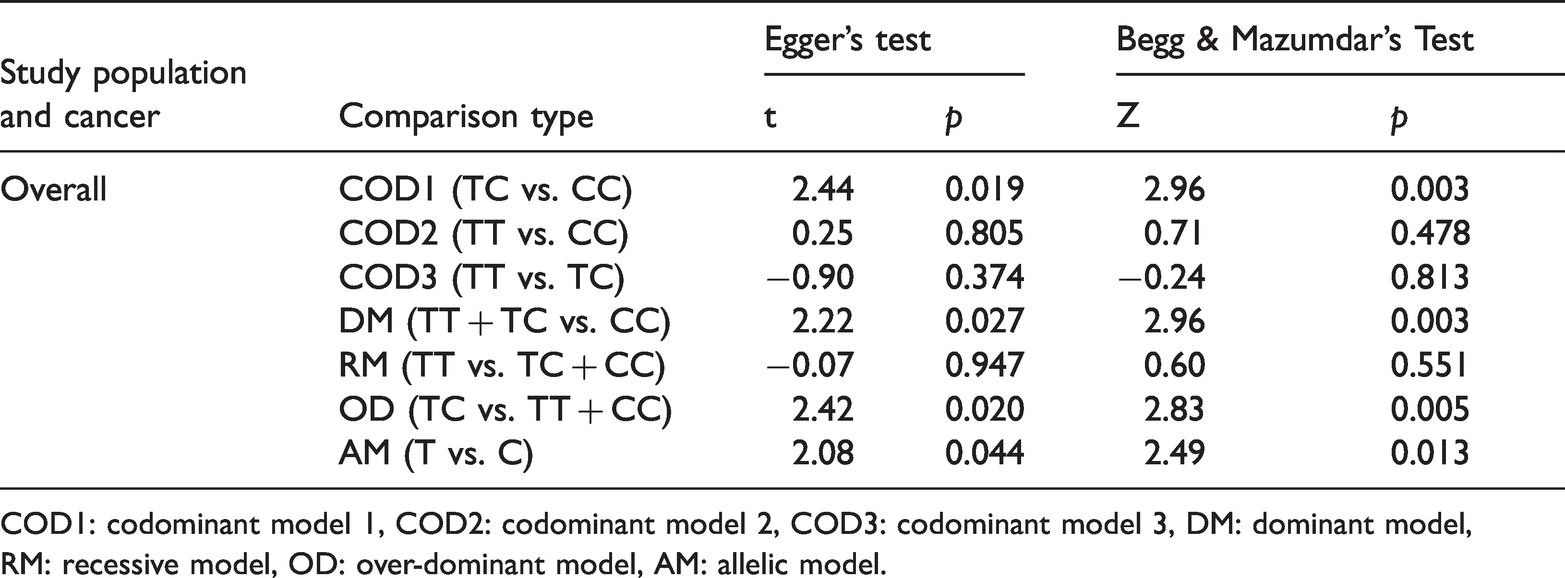
COD1: codominant model 1, COD2: codominant model 2, COD3: codominant model 3, DM: dominant model, RM: recessive model, OD: over-dominant model, AM: allelic model.
TSA results
TSA plots revealed that the cumulative Z-curve for rs1143634 crossed conventional and/or trial sequential monitoring boundaries and achieved the RIS in the overall analysis, GC, BC and HB controls, demonstrating that an adequate level of evidence was achieved, and no further studies are required to confirm the results of the present meta-analysis (Figure 4 A–F). For the Asian subgroup of overall cancer, the Z-curve surpassed the trial sequential monitoring boundary but failed to attain the RIS.

Trial sequential analysis for IL-1β rs1143634 in allele models. (a) Overall, (b) White, (c) Asian, (d) Gastric cancer, (e) Breast cancer and (f) Hospital-based controls. The blue line represents the cumulative Z curve, and the red lines indicate the futility boundaries.
IL-1β gene expression
The eQTL analysis from GTEx revealed that the mutant allele of IL-1β rs1143634 leads to increased IL-1β mRNA expression in the colon (p = 0.00095) and skin (p = 0.0032) (Figure 5A & B).
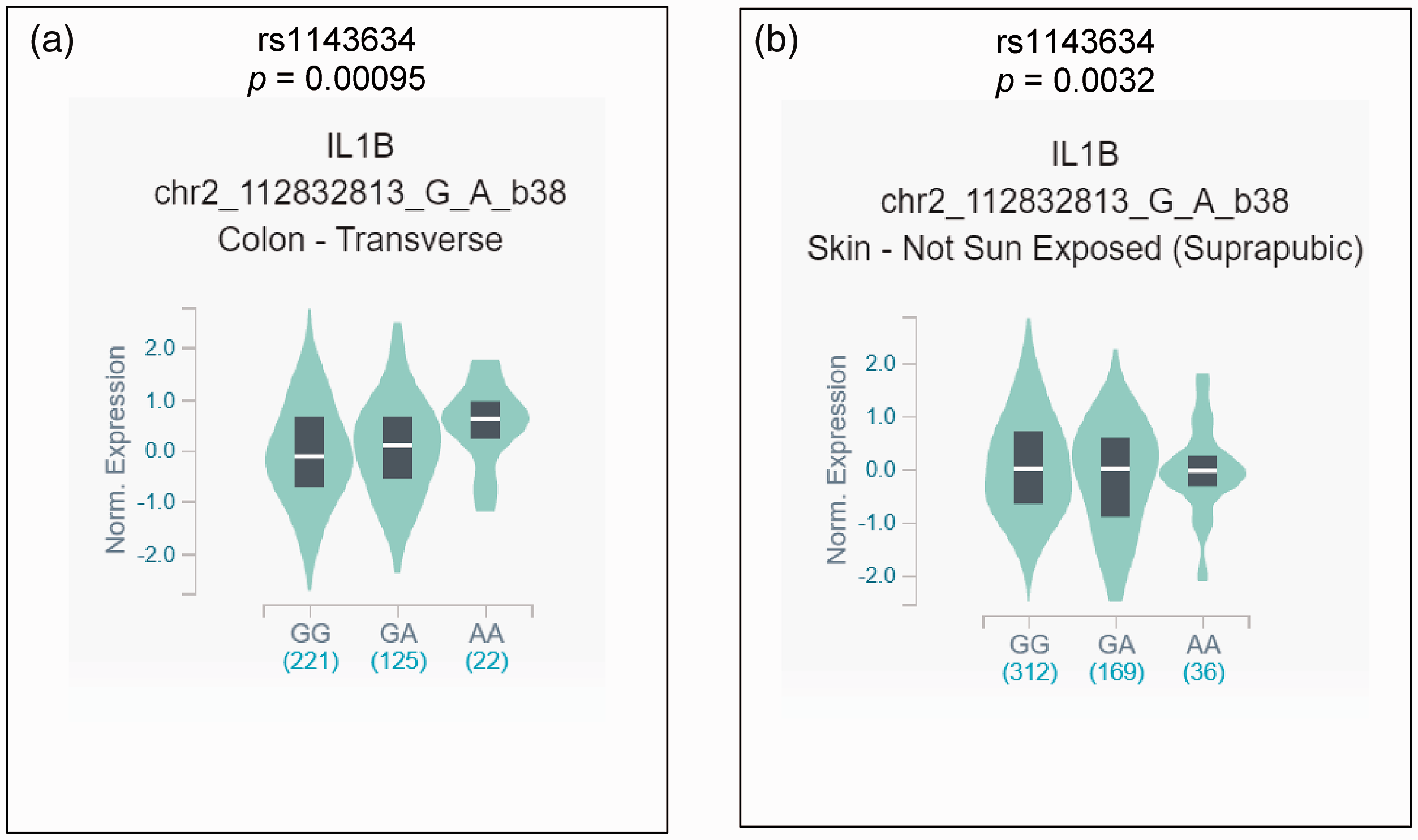
In silico expression analysis of IL-1β in relation to different variants of the rs1143634 polymorphism. (a) There was a significant difference in the expression of IL-1β mRNA in colon tissues depending on the three genotypes, and the variant allele showed higher expression. (b) There was a significant difference in the expression of IL-1β mRNA in the non-sun exposed skin samples, and the variant allele showed lower expression. The values in the brackets represent the frequency of different genotype carriers.
Discussion
Previous reports suggest that cancer and inflammatory cytokines are closely related. For example, the elevated expression of IL-1β in most human cancer types indicates its crucial impact on carcinogenesis. In addition, the increased level of IL-1β restricts improvement in many cancer cases.35,68,69 Because some previous studies provided controversial reports, we attempted to collect and analyze all evidence to understand the role of IL-1β rs1143634 in different cancers.
IL-1β inhibits gastric acid secretion and potentiates chronic inflammation in GC, worsening the disease.70,71 As IL-1β rs1143634 polymorphism increases the production of active IL-1β, this SNP is thought to play a critical role in GC. Zhang et al. showed that the heterozygote model of IL-1β +3954C>T was related to a significantly increased risk of GC. 61 Another study also demonstrated that C>T genotype carriers showed a significantly increased risk of this cancer. 58 Wen et al. reported an elevated risk and added that environmental factors potentiated the chance of cancer development. 57 Similarly, Sakuma et al. provided evidence that polymorphism carriers in the Japanese population might have an increased risk of GC development in the corpus. 45 In contrast, a number of studies showed that the IL-1β rs1143634 variant is not associated with an elevated risk of GC.27,28,40,47,60,67 Moreover, El-Omar et al. stated that this variant might have a protective effect against GC, although the result was statistically non-significant. 17 Persson et al. conducted both PB and HB case–control studies but did not find any connection. 42
IL-1β binds to the estrogen receptor of BC cells and activates transcription. Pooja et al. reported that variant alleles of IL-1β rs1143634 elevated the risk of BC 43 , whereas other previous studies did not find any significant correlations.32,33,63 Two case–control studies reported that this variant might not be related to lung cancer,26,54 whereas Kiyohara et al. and Ter-Minassian et al. indicated that smokers who are mutant T allele carriers of rs1143634 might have a higher risk of lung cancer.34,53 This SNP did not show any association with NSCLC and small cell lung cancer in men.29,35,41 For CRC, only polymorphism carriers among the Chinese Han population showed an increased risk, 56 but other studies reported a negative association.44,64 Patients with MM carrying CT and TT alleles of the IL-1β +3954C>T polymorphism exhibited improved survival rates and survival conditions compared with the CC allele carriers. 25 However, another study in patients with MM did not find any possible association with this polymorphism. 62 Excess IL-1β potentiates inflammation caused by oxidative stress in cancerous pancreatic beta cells, leading to cell destruction and restricted insulin release. It also controls adhesion, invasion and chemoresistance by triggering various signaling pathways, such as nuclear factor kappa B and extracellular signal-regulated kinase.72–75 Cigrovsk Berkovic et al. found a possible association between rs1143634 variant and pancreatic neuroectoderm tumors, although the results were not statistically significant. 66 This SNP did not show a significant association with PCa,38,59 and prior studies31,65 reported no risk for other cancer types. This polymorphism was found to have varied relationships with malignancies in different ethnic populations.9,39
In this meta-analysis, the IL-1β rs1143634 variant showed a significantly elevated association with cancers in three genetic models, COD3 (1.13-fold), RM (1.14-fold) and AM (1.08-fold). The Asian population showed a significantly enhanced risk of cancer in various genetic models, such as COD1 (1.54-fold), DM (1.54-fold), OD (1.48-fold) and AM (1.50-fold). African and White populations did not show any connection between the IL-1β rs1143634 variant and cancer susceptibility. Populations with other mixed ethnicities were significantly associated with cancer risk in COD2 (1.22-fold), COD3 (1.22-fold), RM (1.22-fold) and AM (1.05-fold) models.
We also performed a subgroup analysis with different cancer types. IL-1β rs1143634 polymorphism showed a significant risk association with GC in COD1, DM, OD and AM (1.25-, 1.25-, 1.25-, and 1.21-fold, respectively), BC in COD2 and RM (1.31- and 1.35-fold, respectively) and MM in RM (2.64-fold) models. We found no significant association for the other types of cancers. The selection of controls slightly affected the outcomes according to our sub-group analysis with HB and PB control populations. The analysis with HB controls showed a significantly increased risk in DM (1.18-fold) and AM (1.17-fold) models. No risk association was revealed for the IL-1β rs1143634 variant with cancers in the case of PB controls. The presence of variant alleles of IL-1β rs1143634 might increase the risk of various cancers.
The analysis of sensitivity confirmed the stability and robustness of the present meta-analysis. Moreover, TSA demonstrated that the cumulative Z-curve for the rs1143634 SNP crossed the conventional monitoring boundaries and achieved the RIS, demonstrating that adequate evidence was achieved for this meta-analysis and that no additional studies are needed to verify the results. However, in the Asian population, the Z-curve surpassed the trial sequential monitoring boundary but failed to attain the RIS. Furthermore, we conducted IL-1β gene expression analysis through eQTL, which revealed that mutant alleles of IL-1β rs1143634 lead to increased IL-1β mRNA expression in both colon tissues.
Our meta-analysis had some limitations that could not be avoided. Some of the subgroup analyses showed significant heterogeneity based on the Q-test analysis. Visual asymmetry, Egger’s and Begg & Mazumdar’s tests reported the presence of possible publication bias in a few models. Finally, because of missing information, we failed to provide additional data for individuals, such as their age, sex or disease duration, that may have enriched the quality of the investigation.
Conclusions
Our meta-analysis revealed that the presence of IL-1β rs1143634 variant might elevate the cancer risk in the overall population. Among rs1143634 polymorphism carriers, the Asian population has a greater risk than other ethnic populations. Further studies with larger sample sizes, specific ethnicities and unbiased populations with detailed individual information should be conducted to confirm our findings.
Supplemental Material
sj-pdf-1-imr-10.1177_03000605211060144 - Supplemental material for Role of IL-1β rs1143634 (+3954C>T) polymorphism in cancer risk: an updated meta-analysis and trial sequential analysis
Supplemental material, sj-pdf-1-imr-10.1177_03000605211060144 for Role of IL-1β rs1143634 (+3954C>T) polymorphism in cancer risk: an updated meta-analysis and trial sequential analysis by Sarah Jafrin, Md. Abdul Aziz and Mohammad Safiqul Islam in Journal of International Medical Research
Footnotes
Author contributions
Mohammad Safiqul Islam: conceptualization, supervision, data analysis, software and writing-reviewing and editing; Sarah Jafrin and Md. Abdul Aziz: literature search; Sarah Jafrin: writing-original draft preparation, methodology; Md. Abdul Aziz: writing-reviewing and editing. The final version of the manuscript has been reviewed and approved by all authors.
Declaration of conflicting interest
The authors declare that there is no conflict of interest.
Funding
This research received no specific grant from any funding agency in the public, commercial or not-for-profit sectors.