
Abstract
Objective
Single nucleotide polymorphisms (SNPs) of pentraxin 3 (PTX3) are associated with various outcomes of lung infections. This study aimed to analyze the relationship between PTX3 polymorphisms and the severity of community-acquired pneumonia (CAP).
Methods
This is a retrospective case-control study comprising 43 patients with severe CAP (SCAP) and 97 patients with non-severe CAP. Three SNPs in the PTX3 gene (rs2305619, rs3816527, and rs1840680) from peripheral blood samples were genotyped by real-time polymerase chain reaction. The association between each SNP and the CAP severity was analyzed by logistic regression analysis.
Results
We found that the rs1840680 polymorphism was significantly associated with CAP clinical severity. However, no such association was observed for the genotypes and allele frequencies of rs2305619 or rs3816527. The PTX3 rs1840680 AG genotype was an independent factor for a lower risk of SCAP after multivariate logistic regression analysis. Male sex and coronary heart disease were associated with an increased risk of SCAP.
Conclusions
The PTX3 rs1840680 AG genotype was found to be associated with a lower risk of SCAP, and may serve as a potential protective biomarker to help clinical judgment and management.
Keywords
Introduction
Community-acquired pneumonia (CAP) remains a leading cause of hospitalization and death worldwide.1,2 The short-term mortality (in-hospital mortality or 30-day mortality) rate of hospitalized patients with CAP ranges from 4% to 18%, whereas that of patients with severe CAP (SCAP) reaches 50%.3–6 Timely and accurate evaluation of illness severity, as well as appropriate determination of care intensity and location, is conducive to lethality reduction. 7 However, patients with SCAP usually present with diverse clinical manifestations, such as acute respiratory distress syndrome, septic shock, disseminated intravascular coagulation, and multiple organ dysfunction. 8 There are several factors that contribute to the deterioration of CAP: different pathogen species, variable virulence in microorganisms, older age of host, immune status, and comorbidities. 9 In addition, how the host’s genetic background affects the immune response influences the severity and outcome of CAP.10,11 Studies on polymorphisms of genes that encode inflammatory and anti-inflammatory mediators involved in the pathogenesis of pneumonia have demonstrated contradictory results. For example, a meta-analysis showed that the interleukin 6 (IL6) rs1800795-C allele was associated with excessive systemic inflammatory responses, whereas the IL10 rs1800896-A allele played a protective role in the development of severe inflammation, compared with the other common polymorphisms of proinflammatory cytokines. 12 On the contrary, a multi-centered prospective observational study identified a protective role of heterozygosity for the tumor necrosis factor receptor superfamily member 1B (TNFRSF1B) + 676 single nucleotide polymorphism (SNP) in CAP outcome, whereas no associations were found for the polymorphisms of TNF, lymphotoxin alpha (LTA), IL-6, and IL1 receptor antagonist (IL1RN) in the same population. 10 Taking into account that infectious diseases are related to pathogen-induced immune dysregulation, finding immunological traits presented as host genetic variants can be a way to establish a prediction index in patients suffering from CAP and its complications.
As a member of the pentraxin superfamily, pentraxin 3 (PTX3) is an acute-phase inflammatory mediator that plays an important role in the innate immunity of the lung. 13 PTX3 is produced by numerous cells, including mononuclear phagocytes, dendritic cells, vascular smooth muscle cells, alveolar epithelial cells, endothelial cells, and fibroblasts, at sites of inflammation in response to various inflammatory stimuli such as TNF-α and IL-1β, (nuclear factor kappa B) NF-κB, bacterial lipopolysaccharides, and galactomannan of Aspergillus fumigatus conidia.14–16 It can activate and regulate the complement system via complement component 1q (C1q) and factor H, interact with microbial moieties, and regulate inflammation.15,17 PTX3-deficient mice showed a defective ability to clear pathogens, as well as accelerated death kinetics.18,19 The plasma and alveolar lavage fluid levels of PTX3 have been reported to be related to the clinical severity of CAP.20–22
PTX3 is encoded by the PTX3 gene in humans. Located on chromosome 3 band q25, it consists of three exons and two introns, spanning approximately 7 kb. 23 PTX3 polymorphisms have been disclosed as important risk factors across different infectious diseases.24,25 Cunha et al. 24 found that among 22 PTX3 SNPs, only rs2305619, rs3816527, and rs1840680 remained significantly associated with invasive aspergillosis in the recipients with hematopoietic stem-cell transplantation (HSCT). Chiarini et al. 25 also showed that the haplotypes of rs2305619, rs3816527, and rs1840680 were associated with lung colonization by Pseudomonas aeruginosa (PA) in patients with cystic fibrosis. Despite the known association of PTX3 levels with CAP severity and the disease-relevance of the SNPs in the PTX3 gene, no data are available on the association between PTX3 polymorphisms and the clinical severity of CAP. The aim of our study is to investigate the association between three candidate PTX3 variants (rs3816527, rs2305619, and rs1840680) and the clinical severity of CAP.
Patients and methods
Editorial policies and ethical considerations
This retrospective case-control study adhered to the tenets of the Declaration of Helsinki and corresponded to the STROBE checklist for case-control studies. 26 It was approved by the Ethics Committee of Sun Yat-sen Memorial Hospital, Sun Yat-sen University (No: SYSEC-KY-KS-2020-005) on 8 January 2020. Written informed consent was obtained from each patient or their legal representative before entering the study.
Study design and patient recruitment
We searched the electronic medical records for patients admitted to the Department of Respiratory and Critical Care Medicine of Sun Yat-sen Memorial Hospital between 1 January 2018 and 31 December 2019. Adult patients (aged > 18 years) with CAP were enrolled and divided into a SCAP group and non-severe community-acquired pneumonia (NSCAP) group based on their conditions upon admission as the case and control groups, respectively. The severity of pneumonia was in accordance with the criteria established by the American College of Chest Physicians in 2007. 27 Patients with SCAP needed to meet at least one major criterion (invasive mechanical ventilation or septic shock with the need for vasopressors) or at least three minor criteria (respiratory rate ≥30 breaths/minute, oxygenation index [PaO2/FiO2] ratio ≤250, multi-lobe infiltrates, hypothermia [core temperature <36°C], leukopenia [white blood cell count <4000 cells/mm3], thrombocytopenia [platelet count <100,000 cells/mm3], hypotension requiring aggressive fluid resuscitation, confusion/disorientation, and uremia [blood urea nitrogen ≥20 mg/dL]). 27 The exclusion criteria were as follows: the presence of any malignancy, active tuberculosis, pulmonary cystic fibrosis, pulmonary embolism, pregnancy, HSCT, solid organ transplantation, or other immunosuppressive conditions. Additionally, admitted patients in our hospital are routinely inquired regarding whether discarded biological specimens can be used in medical research work. Patients who did not provide informed consent were excluded from this study.
Demographics and clinical parameters
We reviewed the medical records of all recruited patients and extracted data regarding demographics and clinical parameters, including age, sexuality, length of hospital stay, comorbidities, pneumonia severity index (PSI) scores, and 30-day mortality.
DNA extraction and genotyping
We obtained samples by isolating total blood mononuclear cells from whole blood samples (about 2 mL) taken from each participant’s stored specimens using the erythrocyte lysate (Sango Biotech, Shanghai, China) according to the manufacturer's instructions. We extracted total DNA with a DNA extraction kit (Generay Biotech, Shanghai, China). Three SNPs (rs2305619, rs3816527, and rs1840680) of PTX3 were candidates based on their associations and functional consequences with pulmonary infectious diseases in previous studies.24,28–30 We conducted real-time polymerase chain reaction (qPCR) in triplicate using the TaKaRa Premix Ex Taq II Kit (TaKaRa Bio Inc., Dalian, China) according to the manufacturer’s instructions. A melting curve analysis according to the fluorescent color of the probes was used to define the genotyping of the SNPs, which was analyzed with CFX Manager software 1.6 (Bio-Rad, Hercules, CA, USA).
Statistical analysis
Statistical analysis of the data was performed using SPSS statistical software package version 25.0 (SPSS Inc., Armonk, NY: IBM Corp, USA). Demographics and clinical characteristics of the SCAP and NSCAP groups were compared using a t-test for continuous variables and a chi-square test or rank sum test for categorical variables. Hardy–Weinberg equilibrium of the three allele distributions of the PTX3 polymorphisms was evaluated using the chi-square test. Univariate logistic regression was used to estimate the associations between the PTX3 allele and genotype distributions and SCAP risk. Multivariate logistic regression was performed to identify independent risk factors associated with SCAP. Variables associated with SCAP risk at P < 0.1 in the univariate analyses were considered for entry in the multivariate analysis. The results are presented using odds ratios (ORs) with 95% confidence intervals (CIs). The threshold for statistical significance was P < 0.05.
Results
Patient characteristics
One hundred and fifty-three eligible patients with CAP were invited to participate in this study. In total, 140 patients with CAP were recruited (43 patients with SCAP and 97 patients with NSCAP), and the response rate was 93.5%. The average age of the study participants was 61.21 ± 16.22 years. The demographics and clinical characteristics of the study groups were compared, and male sex, diabetes, and coronary heart disease were more prevalent in the SCAP group than in the NSCAP group (P < 0.05; Table 1). The length of hospital stay was longer in the SCAP group than in the NSCAP group (median days 19 vs. 9, P < 0.001). The 30-day mortality was 27.9% in the SCAP group, whereas no deaths occurred during that follow-up period in the NSCAP group. There were no significant differences in age, hypertension status, or chronic COPD status between the SCAP group and the NSCAP group (all P > 0.05).
Characteristics of the SCAP and NSCAP groups.
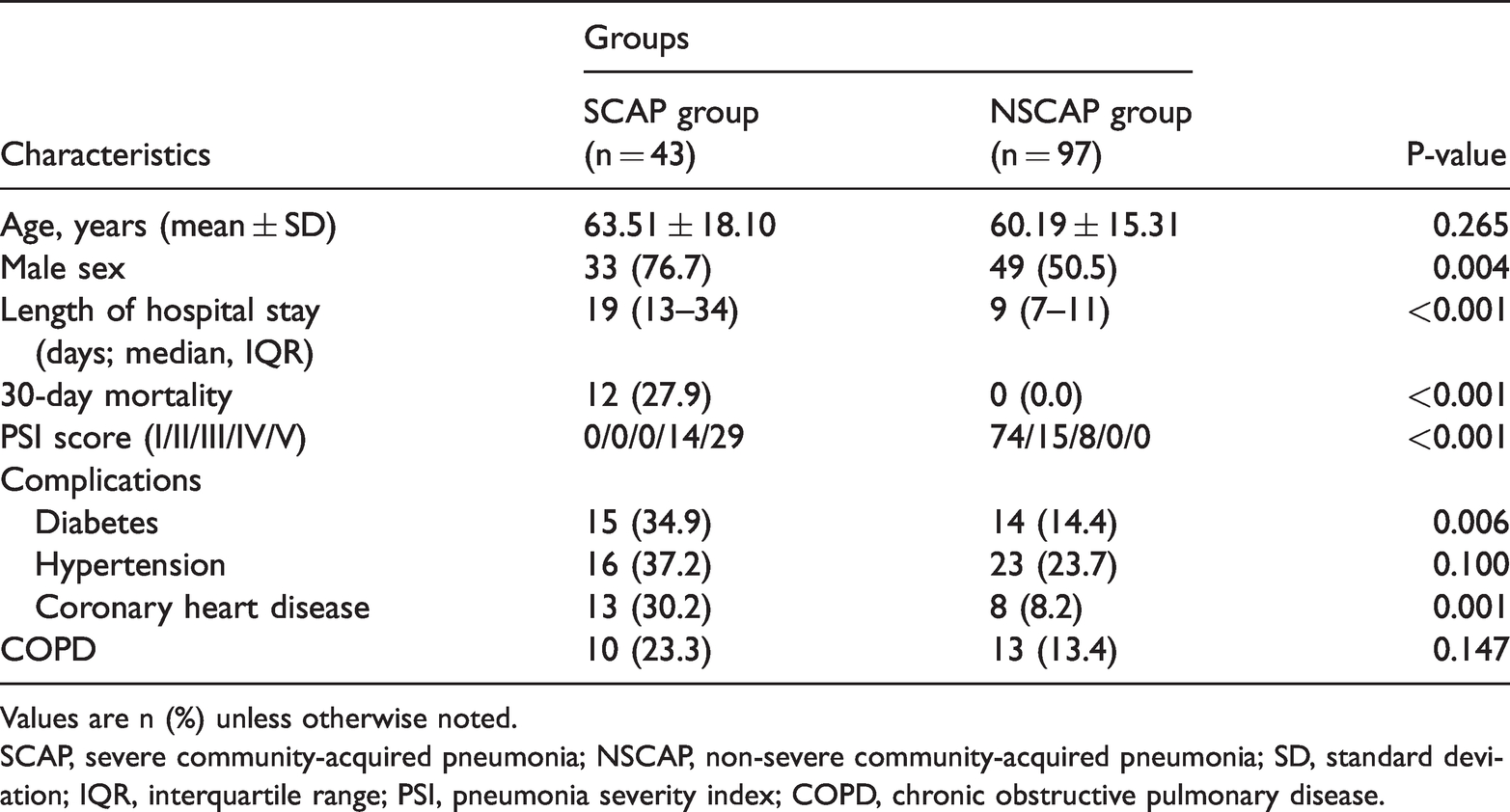
Values are n (%) unless otherwise noted.
SCAP, severe community-acquired pneumonia; NSCAP, non-severe community-acquired pneumonia; SD, standard deviation; IQR, interquartile range; PSI, pneumonia severity index; COPD, chronic obstructive pulmonary disease.
Associations between PTX3 SNPs and CAP severity
To assess the population genetic stability of the current study, Hardy–Weinberg equilibrium analysis was conducted. All allele frequencies of the three SNPs followed the Hardy–Weinberg equilibrium for all participants. We then investigated the relationships between the abovementioned genetic variants and CAP severity. We observed that the allele (Table 2) and genotype (Table 3) distributions of rs2305619 and rs3816527 were not significantly different between the SCAP and NSCAP groups (all P > 0.05) by univariate logistic regression, even after adjustment for age and sex (all P > 0.05). Additionally, no significant difference was found in the rs1840680 allele distribution between the SCAP and NSCAP groups (P > 0.05). Intriguingly, the genotype frequencies of rs1840680 differed significantly between the SCAP and NSCAP groups (P = 0.034; Table 3). Furthermore, the frequency of the rs1840680 AG heterozygosity was lower in the SCAP group than in the NSCAP group (23.3% vs. 46.4%; Table 3). In accordance with this result, AG heterozygosity was significantly associated with a decreased risk of SCAP compared with AA and GG homozygosity (OR = 0.350, 95% CI = 0.155–0.789, P = 0.011).
Distribution of PTX3 gene polymorphism alleles in the SCAP and NSCAP groups.

PTX3, pentraxin 3; SCAP, severe community-acquired pneumonia; NSCAP, non-severe community-acquired pneumonia; SNP, single nucleotide polymorphism; OR, odds ratio; CI: confidence interval.
Distribution of PTX3 gene polymorphism genotypes in the NSCAP and SCAP groups.
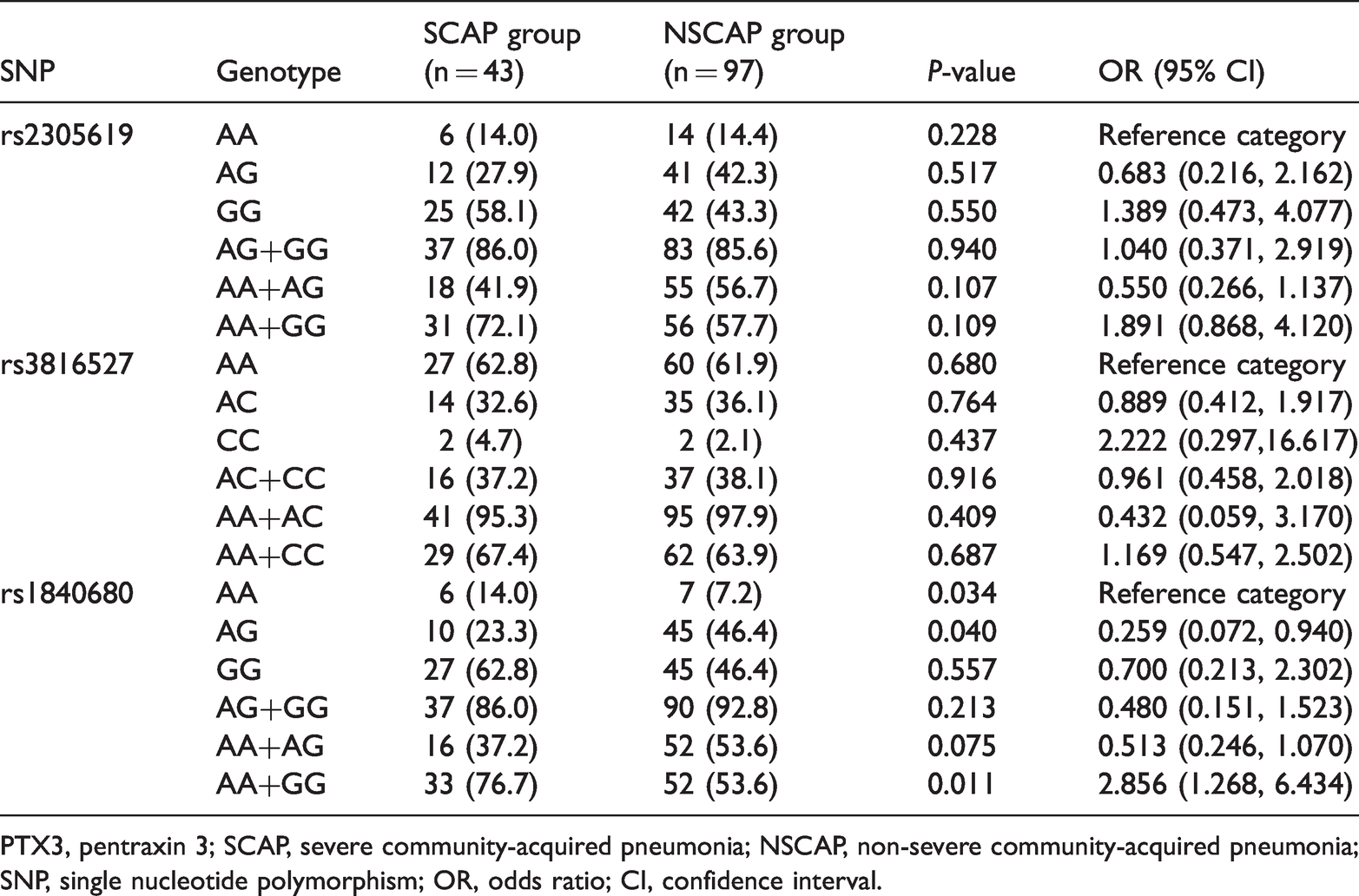
PTX3, pentraxin 3; SCAP, severe community-acquired pneumonia; NSCAP, non-severe community-acquired pneumonia; SNP, single nucleotide polymorphism; OR, odds ratio; CI, confidence interval.
In a multivariate logistic regression analysis accounting for sex, coronary heart disease, diabetes, and PTX3 rs1840680, we further determined that the PTX3 rs1840680 AG genotype was an independent factor for a lower risk of SCAP compared with the PTX3 rs1840680 AA or GG genotypes (OR = 0.386, 95% CI = 0.161–0.929; P = 0.034; Table 4). However, male sex (OR = 4.223, 95% CI = 1.678–10.629; P = 0.002) and coronary heart disease (OR = 4.614, 95% CI = 1.502–14.175; P = 0.008) were independently associated with an increased risk of SCAP in this model.
Multivariate logistic regression analysis for independent risk factors associated with SCAP.

SCAP, severe community-acquired pneumonia; OR, odds ratio; CI, confidence interval; PTX3, pentraxin 3.
Discussion
The current study aimed to investigate the relationship between genetic variation in pentraxin-3 and clinical severity of CAP in a Chinese population by analyzing three SNPs (rs3816527, rs2305619, and rs1840680) in subjects with severe CAP and control subjects. We observed a significantly lower frequency of the rs1840680 AG heterozygosity in the PTX3 gene of the patients with SCAP when compared with those with NSCAP, indicating a potential protective role of this polymorphism in the progression of CAP. To our knowledge, this is the first report studying the association between PTX3 polymorphisms and the clinical severity of patients with CAP in a Chinese population.
According to previously published studies, the variants of the PTX3 rs1840680 gene relate to the pathogenesis of pulmonary infections by diverse microorganisms among different ethnicities. Early research conducted by Chiarini et al. 25 revealed a lower incidence of the rs1840680 AG genotype in participants with PA compared with that in the controls (44.0% vs. 48.1%, P = 0.059), as well as a lower frequency of the “G-A-G” haplotype (SNPs position: rs2305619, rs3816527, and rs1840680) in the cases than in the controls (42.7% vs. 58.7%; OR = 0.52, 95% CI = 0.26–1.07; P = 0.012) by analyzing linkage disequilibrium. In another study, He et al. 30 found a significant association between the rs1840680 AA homozygote and the susceptibility of invasive pulmonary aspergillosis in a Chinese COPD population, but a lower incidence of the rs1840680 AG genotype was still seen in the case group than in the control group (13.89% vs. 40.15%, P = 0.000044). The aforementioned findings, together with our study, suggest a potential protective effect of the PTX3 rs1840680 polymorphism in the context of various pulmonary infections.
As a soluble pattern recognition receptor (PRR), PTX3 can recognize microorganisms invading the lower respiratory tract, activate the classical complement pathway, facilitate opsonization, and enhance pathogen clearance by alveola macrophages and neutrophils in the early stage of inflammation. 15 The primary structure of human PTX3 is highly conserved during evolution, with an N-terminal domain encoded by the first two exons and a C-terminal pentraxin-like domain encoded by the third exon. 31 Thus, PTX3 gene variations associated with protein structure or expression levels may inevitably influence the susceptibility to lung infections caused by different pathogens. In a murine model with PTX3 gene deficiency, increases in both airway colonization and mortality for the ptx3−/− mice infected with PA were observed. 19 Additionally, defective resistance to lung infection by Aspergillus fumigatus and a massive inflammatory response were also observed in this model. 19 However, a dual effect of PTX3 was shown in PTX3 transgenic mice with Klebsiella pneumonia pulmonary infection. 32 In transgenic mice given a high inoculum, faster lethality was associated with overt PTX3 expression, increasing nitrate levels in plasma, diminished TNF-α production and neutrophil influx, and greater dissemination of bacteria to blood at 20 hours after infection. 32 In contrast, transgenic PTX3 expression conferred protection to mice given a lower inoculum, with enhanced TNF-α production, greater neutrophil influx into the lung, and phagocytosis of bacteria by migrated neutrophils. 32 Moreover, Koh et al. 33 demonstrated up-regulated PTX3 expression in the lungs of mice with pneumococcal infection and overexpression of PTX3 that enhanced the pneumococcus-related cytokine response at the location of infection. However, cytokine expression was inhibited in the murine model with PTX3 gene knockdown, indicating that PTX3 deficiency ameliorated acute inflammation by pneumococcal toxin pneumolysin. 33 These findings emphasize the mediator effect of PTX3 in the innate immune response to pulmonary infections. This is consistent with the previous clinical observation that persisting high levels of circulating PTX3 were related to the severity of sepsis and long-term mortality. 34
Variants in PTX3 rs1840680 are reportedly associated with circulating PTX3 levels.30,35,36 Carmo et al. 35 found that in 524 patients with chronic hepatitis, the mean PTX3 plasma level of the rs1840680 AG genotype was significantly lower than that of the AA genotype (1.62 ng/mL vs. 2.80 ng/mL, P = 0.008), but slightly higher than that of the GG genotype (1.62 ng/mL vs. 1.55 ng/mL). Barbati et al. 36 found a similar result where the AG genotype of PTX3 rs1840680 was associated with lower PTX3 levels compared with the AA genotype, but associated with higher PTX3 levels compared with the GG genotype in patients with or without acute myocardial infarction (2.54 ng/mL vs. 2.60 ng/mL vs. 2.10 ng/mL, P = 0.06; 1.49 ng/mL vs. 1.61 ng/mL vs. 1.47 ng/mL, P = 0.009). Taking into consideration that severe CAP and its complications are generally caused by excessive release of inflammatory cytokines and an imbalance between inflammatory and anti-inflammatory reactions,37,38 it is intriguing that a supposed “defective” production of PTX3 could be one factor responsible for disease severity. Further validation of the association between the genetic variant and PTX3 expression in a real-world scenario is needed.
As previously mentioned, a multi-center, prospective observational study suggests a protective role of heterozygosity at the TNFRSF1B + 676 SNP in the outcome of CAP. 10 Actually, molecular heterosis is common in humans and may occur in up to 50% of all gene associations. 39 Molecular heterosis is attributed to the phenomenon of overdominance, where the heterozygote phenotype is superior to that of homozygotes. 40 Heterozygosity may facilitate the dynamic balance between pro-inflammatory and anti-inflammatory factors, thus providing better defense against exposure to a wider range of causative agents and exogenous/endogenous pathogens. 40 Therefore, the protective effect for heterozygotes at the PTX3 gene may be attributed to this phenomenon. In terms of translational application, further investigation of the molecular mechanisms underlying PTX3 gene polymorphisms in patients with CAP may help clinicians to identify patients who are prone to developing SCAP.
This study has some clear limitations. First, the sample size was relatively small because our study was conducted at a single medical center. Second, all participants were from the southern part of China, so there may have been selection bias. It is necessary to conduct external validation in other ethnicities. To overcome these limitations, we suggest that further population-based studies with larger sample sizes be conducted to establish the role of PTX3 gene polymorphisms in patients with CAP.
Conclusions
Our findings demonstrate that the PTX3 rs1840680 AG genotype was associated with a protective effect against severe disease in patients with CAP. Our results highlight the important impact of genetic variation on the progression of CAP and may add a potential biomarker to assist in evaluating the severity of CAP. More studies are necessary to elucidate the mechanisms concerning the role of PTX3 gene polymorphisms in patients with CAP.
Footnotes
Author contributions
Funding acquisition: SJ. Conceptualization and design: QZ, LH, and SJ. Clinical data acquisition: QZ, BH, SB, YX, YD, ZW, TT, and LH. Data analysis and interpretation: QZ, TT, and BH. Original manuscript writing: QZ, TT, and BH. All authors were involved in manuscript review and approval.
Declaration of conflicting interest
The authors declare that there is no conflict of interest.
Funding
This work was supported in part by the National Natural Science Foundation of China (81670022, 82071804), Natural Science Foundation of Guangdong Province (2017A030313681), and Guangzhou Municipal Science and Technology Project (201704020123).