
Abstract
Objectives
To investigate the influence of cyclosporin A (CsA) pre-treatment and etomidate (ETO) post-treatment on lung injury induced by limb ischemia-reperfusion (I/R) in rats.
Methods
Rats were randomly divided into five groups: sham, I/R, I/R+CsA, I/R+ETO, and I/R+CsA+ETO. Limb I/R lung injury was established by bilateral clamping of the femoral arteries for 2 hours. Following reperfusion for 3 hours, blood gas analysis was performed. Pathological changes were assessed using immunohistochemistry. The apoptosis index (AI) and wet/dry weight ratio (W/D) were calculated. Levels of Fas protein and FasL mRNA were assessed by western blotting and RT-PCR, respectively. Tumor necrosis factor (TNF)-α and interleukin (IL)-1β were detected by ELISA.
Results
I/R resulted in decreased PaO2 but increased AI, W/D, Fas, FasL mRNA, TNF-α and IL-1β. Scattered punctate apoptosis and necrosis were observed by immunohistochemistry. Compared with the I/R group, the I/R+ETO and I/R+CsA groups showed increased SpO2, decreased AI, W/D, Fas, FasL mRNA, TNF-α and IL-1β, and decreased numbers of apoptotic and necrotic cells. Combined treatment with CsA+ETO resulted in more dramatic changes in these parameters.
Conclusions
ETO post-treatment and CsA pretreatment reduced lung injury induced by limb I/R in rats. The mechanism may be related to synergistic inhibition of Fas/FasL signaling.
Lung injury induced by limb ischemia-reperfusion (LILIR) is of high clinical interest. In-depth studies of ischemia-reperfusion (I/R) have found that in addition to tissues directly affected by I/R, distant organs may also be damaged. 1 Some studies showed that etomidate (ETO) can reduce injury associated with brain ischemia-reperfusion by down-regulating Fas/FasL.2,3 Other studies showed that cyclosporin A (CsA) can reduce injury associated with myocardial ischemia-reperfusion by reducing the expression of FasL on the cell surface. 4 The membrane surface molecules Fas and its ligand FasL have a profound impact on the mechanism of apoptosis. 5 Our previous studies found that limb I/R could induce kidney and liver injuries in rats.6,7 Based on these results, we wondered whether ETO post-treatment and CsA pre-treatment would affect Fas/FasL signaling during limb I/R in rats. Clinically, almost all patients undergoing lower limb surgery experience injuries caused by limb I/R. Therefore, it is necessary to better understand the mechanisms of limb I/R injuries in animal models. The aim of study was to investigate the synergistic effects of ETO post-treatment and CsA pre-treatment on limb I/R injuries in rats.
Materials and methods
Randomization
A random number table method was used to select animals and assign them to treatment groups.
Time and place
Time: 2019. Setting: Central Hospital Affiliated to Shenyang Medical College
Materials
Experimental methods
Ethics
The animal study was approved by the local Ethics Board of the Central Hospital of Shenyang Medical College (Shenyang City, China).
Blood gas analysis
Blood (3 mL) was obtained from the carotid artery following 3 hours of limb reperfusion. Blood gas analysis was conducted immediately using a Gem Premier 3000 Blood Gas Analyzer (LeiCheng Trading Co., Ltd., Nanjing City, China) and PaO2 was recorded.
Biochemical indexes and wet/dry weigh ratio (W/D)
Levels of tumor necrosis factor (TNF)-α and interleukin (IL)-1β were assessed using an ELISA kit (Abcam Co., Cambridge, UK). Following limb reperfusion for 3 hours, tissue from the upper pole (1 cm3) of the left lung was used to produce 10% lung tissue homogenate. For ELISA, lung tissue homogenates (100 µL/well) were added, then plates were sealed and incubated at 37°C in the dark for 2 hours. The plates were washed, a biotinylated antibody (100 µL/well) was added, and the plates were re-sealed and incubated at room temperature for 1 hour. The plates were washed again and a horseradish peroxidase-conjugated secondary antibody (100 µL/well) was added and incubated at room temperature for 20 minutes. The plates were washed a final time, developer (100 µL/well) was added, and the plates were incubated in the dark for 20 minutes. The reaction was terminated with stop solution (50 µL/well), the plates were gently agitated, and then the absorbance at 450 nm was measured. Absorbance values from blank wells were subtracted from the average values for experimental wells. Standard curves were constructed and used to calculate TNF-α and IL-1β concentrations.
For calculation of W/D, the lower pole tissue of the right lung (1 cm3) was obtained and washed with 4°C normal saline. Residual blood was removed and surface water was dried with filter paper. The wet weight (W) was measured, then the lung tissue was dried at 80°C for 48 hours. The dry weight (D) was measured and then W/D was calculated.
Immunohistochemistry
The streptavidin–biotin complex (SABC) method was used and the experimental steps were carried out according to the manufacturer’s instructions. 3,3′-Diaminobenzidine (DAB) was used to stain samples and positive expression was classified as brown cytoplasm. A sample of the middle pole lung tissue (1 cm3) was obtained from the right lung. The sample was dewaxed, hydrated, washed twice with phosphate-buffered saline (PBS) for 5 minutes, then incubated at room temperature in PBS containing 3% H2O2 for 5 to 10 minutes. The sample was washed with distilled water three times then antigen retrieval was performed. The sample was washed with PBS for 5 minutes, then normal sealing solution was added by dripping and incubated at room temperature for 20 minutes. Excess liquid was removed. Primary antibody (1:100 dilution, Abcam) was added and incubated at room temperature for 1 hour. After washing with PBS three times for 2 minutes, biotinylated secondary antibody (1:100 dilution, Abcam) was added and incubated at 20 to 37°C for 20 minutes. After washing with PBS three times for 2 minutes, SABC reagent was added and incubated at 20 to 37°C for 20 minutes, then the plates were washed with PBS four times for 5 minutes. DAB color was developed using a DAB color development kit.
After washing with distilled water, hematoxylin was added and the samples were set aside for 2 minutes. The samples were then treated with hydrochloric acid and alcohol, dehydrated, sealed and examined under the microscope. Brown-stained cells were classified as positive. Five visual fields were randomly selected from each section, and 100 cells were counted from each visual field. The ratio of brown-stained cells to these 100 cells (RBS) was calculated.
Hoechst33258 staining
A sample of right upper lung tissue (1 cm3) was obtained, embedded and sectioned routinely. The sections were washed with PBS, stained for 5 minutes with Hoechst33258 staining solution (0.5 mL, AAT Bioquest Inc, CA, USA), then washed again with PBS. The sections were placed on slides, anti-quenching sealing solution was added, and the slide was covered. The slides were observed by fluorescence microscopy under 200× magnification. Using a BX-41 microscope (Olympus Corporation, Tokyo, Japan), normal nuclei can be observed as round and dark blue. By contrast, the chromatin of apoptotic cells is pyknotic, and their nuclei are dense or fragmented with a bright white color. Cells with dense or white-bright nuclei were counted. Five visual fields were randomly selected from each section, and 100 cells were counted from each visual field. The ratio of apoptotic cells among these 100 cells was taken as the apoptotic index (AI).
Fas protein expression
Western blotting was used to detect Fas protein expression in lung tissue. Lung tissue (1 cm3) was obtained from the lower pole of the right lung. Residual blood was washed with 4°C normal saline, then the samples were dried with filter paper and 10% tissue homogenate was prepared. Total protein was extracted by centrifugation and protein concentrations were assessed using a DYY-6B electrophoresis instrument (Liuyi Instrument Factory, Beijing, China). Following SDS-PAGE, electrophoretic transfer to polyvinylidene difluoride membranes, and blocking, rabbit anti-rat Fas polyclonal antibody (Abcam) and rabbit anti-rat β-actin polyclonal antibody (Abcam) were added and incubated at 4°C overnight. Alkaline phosphatase conjugated secondary antibody (Sigma) was added, incubated at room temperature for 2 hours, and then substrate was applied for color scanning. The blots were imaged using the Scion Image Analysis System (Apple, Cupertino, CA, USA). Fas expression was calculated by densitometry by comparison with expression of β-actin.
FasL mRNA expression
RT-PCR was used to detect the expression of FasL mRNA in lung tissue. Lung tissue (1 cm3) from the lower pole of the left lung was used to prepare 10% lung tissue homogenate. Total RNA was extracted using Trizol (Sigma, St. Louis, MO, USA), reverse transcribed into cDNA, and then amplified by PCR (Eppendorf, Hamburg, Germany). The sequences of primers were as follows: FasL mRNA (395 bp, Bao Bioengineering Co., Ltd. Dalian City, China) upstream primer 5′-ACGACACGGACAGC-3′, downstream primer 5′-GATCACGCGAGGAGCAC-3′; and β-actin (198 bp): upstream primer 5′-CATCTCGGATGCACAG-3′, downstream primer 5′-GGACCCGTAGCACG-3′. The reaction conditions for FasL mRNA RT-PCR were: 94°C pre-denaturation for 2 minutes; 35 cycles of 95°C for 45 s, 57°C 45 s, 72°C for 60 s; 72°C extension for 5 minutes. The reaction conditions for β-actin were: pre-denaturation at 94°C for 2 minutes, 30 cycles of 94°C for 40 s, 58°C for 40 s, 72°C for 60 s; 72°C extension for 5 minutes. The PCR products were analyzed by 2% agarose gel electrophoresis and ethidium bromide staining. The expression level of FasL mRNA was assessed based on the relative densitometry of the FasL and β-actin PCR products using the Scion Image Analysis System.
Statistical methods
SPSS version 13.0 software (SPSS Inc., Chicago, IL, USA) was used for all statistical analyses. Data were expressed as means ± standard deviations (x ± s). Differences between groups were assessed using the independent sample t-test, single factor analysis of variance and Fisher’s least significant difference test. Correlational analyses were also performed. Values of P<0.05 were considered statistically significant.
Results
Analysis of the number of experimental animals
All 150 rats survived the procedures and results were available for analysis.
TNF-α, IL-1β, and PaO2
Compared with the sham group, levels of TNF-α and IL-1β were increased and PaO
Comparison of TNF-α, IL-1β, W/D and PaO2 in each group (n = 30, x + s).

aP < 0.05 compared with sham group; bP < 0.05 and cP < 0.05 compared with I/R group; dbP < 0.05 and dcP < 0.05 compared with I/R+CsA group and I/R+ETO groups, respectively.
Immunohistochemistry
Under the light microscope, the lung cells of the sham group rats were orderly, and few leukocytes were observed infiltrating into the alveoli. In the I/R group, many inflammatory cells infiltrated into the alveoli. An increased number of necrotic cells with brownish yellow staining were observed and occasionally aggregated. In the I/R+CsA and the I/R+ETO groups, changes were similar: necrotic foci with dense punctate staining were observed, and inflammatory cells were decreased. In the I/R+CsA+ETO group, necrotic cells were occasionally seen, and the number of inflammatory cells decreased in the alveoli and pulmonary interstitium was reduced (Figure 1 and Table 2).
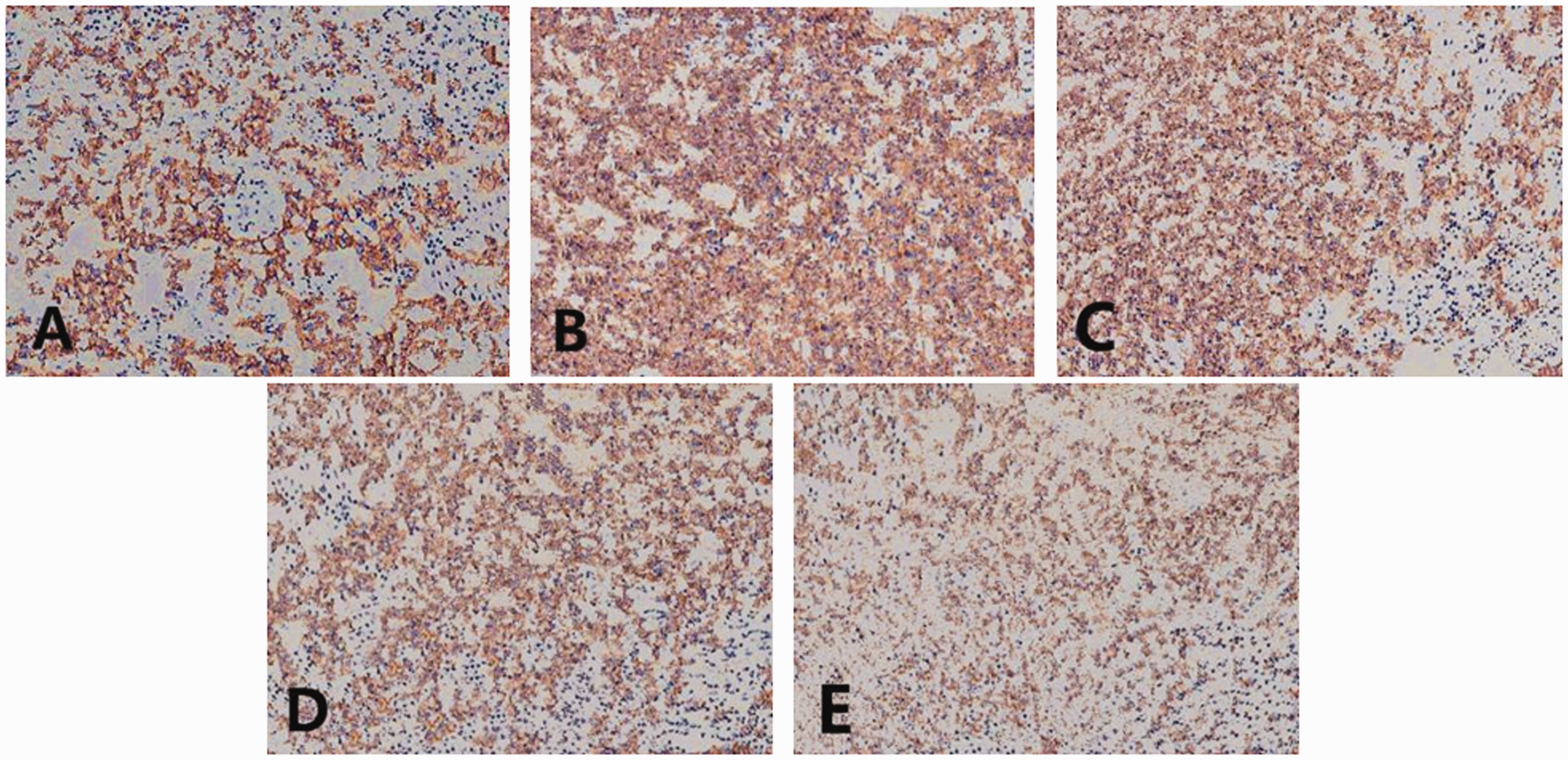
Morphological structures of lung immunohistochemical sections in each group by light microscopy (200×). A: Sham group; B: I/R group; C: I/R+CsA group; D: I/R+ETO group; E: I/R+CsA+ETO group.
Comparison of AI and RBS in each group (n = 30, x + s).

aP < 0.05 compared with sham group; bP < 0.05 and cP < 0.05 compared with I/R group; dbP < 0.05 and dcP < 0.05 compared with I/R+CsA and I/R+ETO groups, respectively.
Hoechst33258 staining and AI
Compared with the sham group, the AI was increased in the I/R group (P = 0.021). Compared with the I/R group, the AI was decreased in the I/R+CsA and I/R+ETO groups (P = 0.034). Compared with the I/R+CsA and I/R+ETO groups, the AI was further decreased in the I/R+CsA+ETO group (P = 0.019) (Figure 2 and Table 2).

Comparison of Hoechst33258 staining in the lung tissues of each group (200×).
Comparison of Fas protein expression
Compared with the sham group, Fas protein expression was increased in the I/R group (P = 0.025). Compared with the I/R group, Fas expression was decreased in the I/R+CsA and I/R+ETO groups (P<0.033). Compared with the I/R+CsA and I/R+ETO groups, Fas expression in the I/R+CsA+ETO group was further decreased (P = 0.017) (Table 3 and Figure 3).
Comparison of relative amount of Fas and FasL mRNA in each group (n = 30, x + s).

aP < 0.05 compared with sham group; bP < 0.05 and cP < 0.05 compared with I/R group; dbP < 0.05 and dcP < 0.05 compared with I/R+CsA and I/R+ETO groups, respectively.
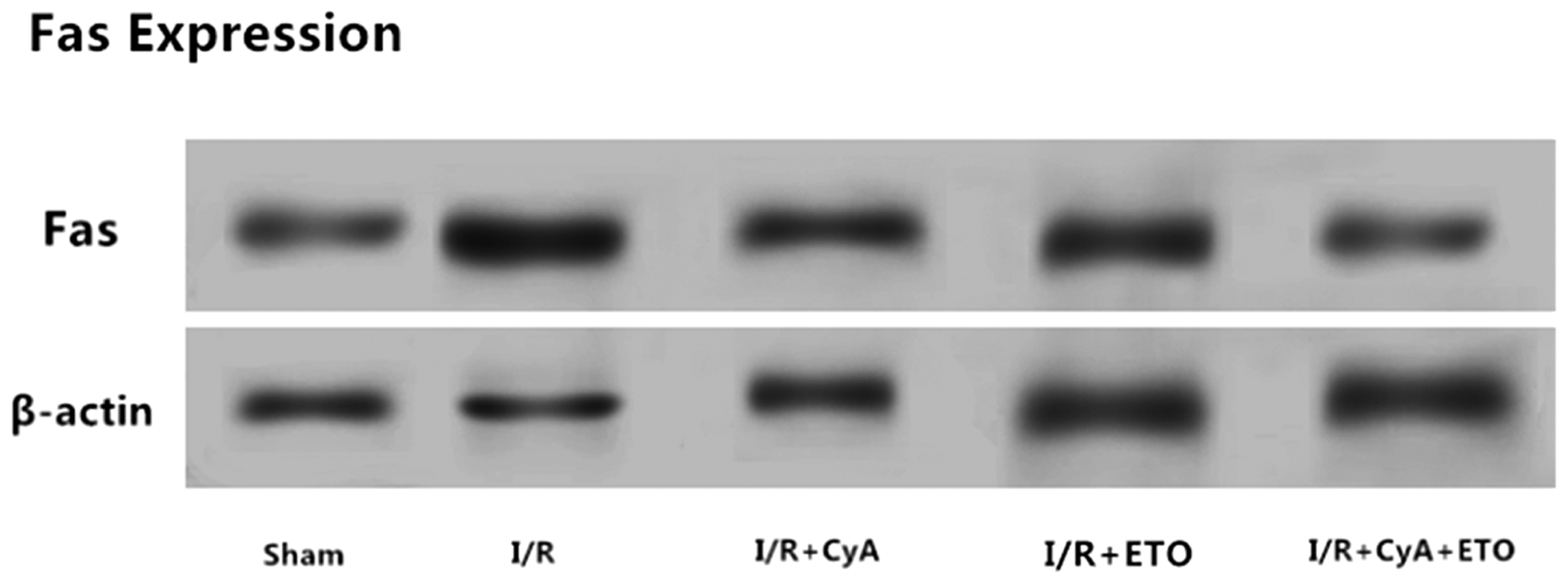
Comparison of Fas protein expression in the lung tissues of each group.
Comparison of FasL mRNA
Compared with the sham group, FasL mRNA levels in the I/R group were increased (P = 0.024). Compared with the I/R group, FasL mRNA levels in the I/R+CsA and I/R+ETO groups were decreased (P = 0.013). Compared with the I/R+CsA and I/R+ETO groups, FasL mRNA levels in the I/R+CsA+ETO group were further decreased (P = 0.021) (Table 3 and Figure 4).

Comparison of FasL mRNA levels in the lung tissues of each group.
Discussion
As minimally invasive surgery becomes more common, microscopy is applied more and more frequently. A clear surgical field becomes the most basic requirement that minimally invasive surgeries must meet. 9 In hand and foot surgeries, the application of tourniquets can meet the requirements of surgeons for clarity of surgical field, but collateral injury occurs from limb I/R.10,11 Earlier studies showed that I/R of the hind limbs in rats can lead to lung injury at distance sites. The mechanisms of injury are diverse, with no unified theory. 12 The Fas protein plays a major role in apoptosis.13,14 Whether Fas protein and its ligand FasL are involved in LILIR was unclear. The purpose of this study was to study the synergistic effects of CsA pre-treatment and ETO post-treatment on LILIR and the dependence of these effects on Fas/FasL.
FasL can bind the death receptor Fas, which induces apoptosis during the development of T-lymphocytes.15,16 Fas and its ligand FasL have been thoroughly studied with a profound impact on our understanding of the mechanisms of apoptosis.17–19
CsA is a cyclic peptide comprising 11 amino acids. It is an active metabolite of fungi in soil and has strong immunosuppressive effects.20–22 CsA can reversibly and selectively alter the function of T-lymphocytes, prevent the transcription of lymphokine genes, interfere with signaling, 19 inhibit release of IL-2, interferon and other immune factors, alter humoral and cellular immunity, inhibit the killing activity of natural killer cells, and inhibit the differentiation and proliferation of lymphocytes.23,24
ETO is used for general anesthesia and is a safe imidazole derivative. Inhibition of respiratory and circulatory functions during induction of general anesthesia with ETO is relatively slight. 24 Some studies have shown that ETO pre-treatment can reduce the expression of Bax protein during LILIR in rats, mediating a protective effect. 17 ETO may play a protective role in reperfusion injury by activating gamma-aminobutyric acid receptors, increasing the permeability of the cell membrane to Cl ions and preventing depolarization the cell membrane. 25 In addition, the N-methyl-D-aspartate receptor and the Ca2+-channel are voltage dependent; inhibition of cell membrane depolarization leads to decreased Ca2+ influx and decreased cellular oxygen consumption. Therefore, ETO can increase the tolerance of cells to ischemia-reperfusion injury.2,26
Compared with the sham group, Fas, FasL mRNA and other injury indexes were increased, while PaO2 was decreased in the I/R group. This suggested that establishment of the rat LILIR model was successful. We speculate that the mechanism of lung injury is probably related to Fas/FasL. Compared with the I/R group, Fas and FasL mRNA decreased while PaO2 increased in the I/R+CsA and I/R+ETO groups. This suggested that both CsA pre-treatment and ETO post-treatment can reduce lung injury. The protective effects of CsA pre-treatment and ETO post-treatment were associated with inhibition of Fas/FasL. Compared with the I/R+CsA and I/R+ETO groups, Fas, FasL mRNA and other injury indexes were further decreased and PaO2 was further increased. It was previously shown that CsA pre-treatment can down-regulate the expression of FasL protein on the cell surface, thus inhibiting Fas/FasL signaling. 27 Based on these results, we conclude that CsA pretreatment and ETO post-treatment may have synergistic protective effects against LILIR in rats.
In conclusion, our results showed that both CsA pretreatment and ETO post-treatment can reduce LILIR in rats. The mechanism may be related to down-regulation of Fas/FasL expression. In addition, CsA pretreatment and ETO post-treatment have synergistic protective effects against LILIR in rats.
Footnotes
Acknowledgements
We thank the teachers and students who participated in the design and implementation of the animal experiments. We are also grateful for the laboratories and related equipment provided by the teachers of the animal experiment center. We thank those who helped with the purchase of experimental reagents. Finally, we would like to thank the hospital and department leaders for their support in this study.
Declaration of conflicting interest
The authors declare that there is no conflict of interest.
Funding
This study was supported by funding from the Guidance Plan Project of the Liaoning Province Natural Science Foundation (20180550618), the General Scientific Research Project of the Liaoning Provincial Department of Education (L2015540), the Science and Technology Fund Project of Shenyang Medical College (20191013), and the Science and Technology Fund Project of Shenyang Medical College (20162029).