
Abstract
Objective
The current study investigated the effect of c-Jun NH2-terminal kinase (JNK) expression on the growth of transplanted breast cancer tumors in mice.
Methods
A breast cancer transplantation model was established in BALB/c mice, which were then treated with SP600125 (30 mg/kg) for 24 days. After sacrificing the mice, the inhibitory effects of SP600125 on breast cancer growth were calculated by weighing tumors. Moreover, vascular endothelial growth factor (VEGF) expression and the tumor microvascular density (MVD) were evaluated via immunohistochemistry. Cell apoptosis was also examined using a TUNEL kit.
Results
Compared with the findings in the control group, SP600125 treatment (30 mg/kg) obviously suppressed tumor growth during the 15-day observation period. SP600125 treatment markedly inhibited JNK mRNA expression. Furthermore, VEGF protein expression (50% vs. 100%) and MVD (18.27 ± 1.70 vs. 23.17 ± 4.02) were also significantly decreased by SP600125 treatment, whereas the apoptosis index was significantly higher in the treatment group (10.23 ± 1.97% vs. 4.53 ± 1.40%).
Conclusion
Inhibition of JNK signaling can significantly suppress the growth of transplanted breast tumors in mice.
Keywords
Introduction
As an important component of the mitogen-activated protein kinase (MAPK) pathway, c-Jun NH2-terminal kinase (JNK) is extremely important for the migration, proliferation, differentiation, and death of cells.1,2 The JNK signaling pathway can be activated by a series of stimuli, such as growth factors, cytokines, injury, and chemical carcinogens, and JNK signaling widely participates in apoptosis, proliferation, metabolic activity and cell repair.3–5 In addition, the JNK signaling pathway plays an important role in the occurrence and progression of oculopathy, neurodegeneration, endocrine disease, chronic inflammation, and other conditions. Furthermore, increasing evidence has revealed that the JNK signaling pathway is closely related to the occurrence of several cancers.6–8 Additionally, JNK signaling can be excessively activated in breast cancer cells, suggesting close relationships of the JNK pathway with the occurrence and development of breast cancer.9,10
Breast cancer is one of the most common cancers in women worldwide, and most patients are diagnosed with mid- or late-stage disease. 11 As the main treatment for breast cancer, surgery is sometimes considerably limited in terms of efficacy and patient suitability. Thus, new targets for immunotherapy, as an alternative to surgery, 12 have been actively studied for the treatment of breast cancer.
SP600125, an ATP-competitive JNK inhibitor, has been used to characterize the role of JNK in apoptotic pathways.13,14 In this investigation, the effect of SP600125 on transplanted breast cancer tumors in mice was explored. Furthermore, whether the JNK signaling pathway affects the growth of breast cancer was studied.
Materials and Methods
Reagents
SP600125 was purchased from (MedChemExpress, Monmouth Junction, NJ, USA). Rabbit anti-mouse anti-factor VIII and rabbit anti-mouse VEGF polyclonal antibodies were obtained from NovoPro Bioscience (Shanghai, China). A TUNEL kit was purchased from Roche (Basel, Switzerland). DBA color development reagent was procured from Beyotime (Shanghai, China).
Animals and cell strain
Female SPF BALB/c mice (15–20 g, HFK Bioscience Co., Ltd., Beijing, China) were raised in the animal room of the State Key Laboratory of Sichuan University (No. 2018-31) under the following conditions: temperature, 25 to 27°C; humidity, 45% to 50%; high-level dust removal and decontamination; and a specific pathogen-free (SPF) environment. Mice were granted access to a normal diet and aseptic water. Mouse mammary adenocarcinoma cells (EMT6) were purchased from the China Center for Type Culture Collection (Wuhan, China).
Mice breast cancer transplantation model
EMT6 cells were placed in a 5-mL culture bottle at a concentration of 5 × 107/mL. Subsequently, the cells were injected using an aseptic syringe into the abdominal cavity of four BALB/c mice (2 mL each), which were then cultivated. After the mice were sacrificed via cervical dislocation after 9 to 11 days, the abdomen of each animal was wiped with alcohol, the abdominal skin was opened using aseptic dissecting scissors, the abdominal wall was lifted using aseptic tweezers, and ascetic fluid was extracted from the abdominal cavity using aseptic syringes. Then, cells from the subcutaneous tissue of the napes of 12 BALB/c mice (1 × 106 cells/mL) were inoculated at a cell concentration of 1 × 107/mL in RPMI-1640 medium. The inhibition rate of tumor growth was calculated using the following formula:

Grouping and samples
Twelve mice were randomly divided into two groups: experimental and control groups (each n = 6). SP600125 (30 mg/kg, i.p.) was injected into the abdominal cavity of each mouse in the experimental group once a day for 24 days, whereas mice in the control group received injections with an equal volume of PBS. The mice were decapitated after treatment, and the tumors were removed and weighted. The tumor tissue was frozen in liquid nitrogen until use in RT-PCR, and the remaining tissue was used for immunohistochemical evaluation.
Determination of JNK mRNA expression
Total RNA was extracted from tumor tissues and stored in a −80°C freezer. The primers were designed using sequences in GenBank as follows: forward, 5′-CTCTCCAGCACCCGTACATCAA-3′; and reverse, 5′-CTTAGTT-CGCTCCTCCAAATCCA-3′. RNAs were detected by RT-PCR using the manufacturer’s protocols (Promega, Madison, WI, USA).
Immunohistochemistry
Vascular endothelial growth factor (VEGF) expression was analyzed via streptavidin–biotin complex (SABC) immunohistochemistry using a rabbit anti-mouse VEGF polyclonal antibody (Shanghai New Biotechnology Co., Shanghai, China). Sections were cut from routinely prepared paraformaldehyde-fixed, paraffin-embedded blocks of tumor tissue, deparaffinized in xylene, and heated by microwave in sodium citrate buffer solution (0.01 mol/L, pH 6.0). After cooling to room temperature, the tissue sections were incubated with normal goat serum away from light at room temperature for 20 minutes. The tissue sections were incubated with primary antibody, washed three times in phosphate-buffered saline (PBS), and incubated with biotin-conjugated secondary antibody (IgG) at 37°C for 30 minutes. Samples were again washed three times in PBS. The sections were incubated with streptavidin–horseradish peroxidase. Diaminobenzidine (DAB) was used for visualization; in addition, the sections were counterstained with hematoxylin followed by hydrochloric acid alcohol differentiation. The tissue sections were dehydrated, transparentized, mounted, and observed under a microscope at high magnification (×400).
VEGF-positive cells were identified by the presence of brown yellow particles in the cytoplasm. Five representative fields were randomly selected in each section for analysis (approximately 400 cells in a single field). The staining intensity was scored as follows: 0, no staining; 1, light yellow or only individual yellow to brown yellow cells; and 2, yellow to brown staining. The percentage of positive cells was scored as follows: 0, <5%; 1, 5% to 25%; 2, 26% to 50%; 3, 51% to 75%; and 4, >75%. Then, the total score was obtained by summing the scores of staining intensity and positivity as follows: 0 to 1, negative, 2 to 3, weakly positive; 4 to 5, moderately positive; and 6 to 7, strongly positive. All samples were analyzed by the experimenters preliminarily and re-evaluated by pathologists who were blinded to the experiment to reduce bias.
The tumor microvascular density (MVD) was analyzed via SABC immunohistochemistry using anti-Factor VIII as the primary antibody. Samples were processed as previously described. The tissue sections were first observed under low magnification to identify the most densely populated region of the tumor blood vessels and followed by observation under high magnification. Any brown-stained endothelial cells or endothelial cell cluster was considered a single countable microvessel, in addition to the branched structures that were not connected. Blood vessels with a muscular layer or with a lumen diameter exceeding the diameter of eight red blood cells were not considered countable. A modification of Weidner’s method was applied to calculate MVD. 15 Five fields were selected for each tissue section, and the mean value of the five counted fields was taken as MVD of the tumor tissue.
Assessment of cell apoptosis
Samples were exposed to paraformaldehyde for 30 to 60 minutes, and apoptosis was determined using a TUNEL kit according to the manufacturer’s protocols. Five representative fields were analyzed using a microscope at high magnification, and the positive cells (apoptotic cells) were identified by the presence of brown yellow-stained nuclei. The samples were analyzed by the experimenters preliminarily and re-evaluated by pathologists blinded to the experiment to reduce bias. The apoptotic index was calculated as follows: apoptotic index (%) = number of apoptotic cells/total number of cells × 100.
Statistical analysis
Data were analyzed using SPSS version 22.0 software (IBM, Armonk, NY, USA). Continuous variables are presented as the mean ± SD. Student’s t-test was performed to analyze differences between the two groups. P < 0.05 was considered statistically significant.
Ethics approval
This study was approved by the Ethics Committee of Sichuan University (Chengdu, People’s Republic of China).
Results
The anti-tumor activity of SP600125 in tumor-bearing mice
All mice in the experimental and control groups were alive with shiny coats and normal diet consumption, activity, and excretion at the time of sacrifice. As shown in Figure 1, the transplanted tumors in mice were round or ovoid. Furthermore, SP600125 at a dose of 30 mg/kg obviously suppressed tumor growth during the 15-day observation period compared with the findings in the control group (P = 0.027, Table 1 and Figure 2).

Subcutaneously transplanted breast cancer tumors in mice.
The average tumor weight and rate of tumor growth inhibition.

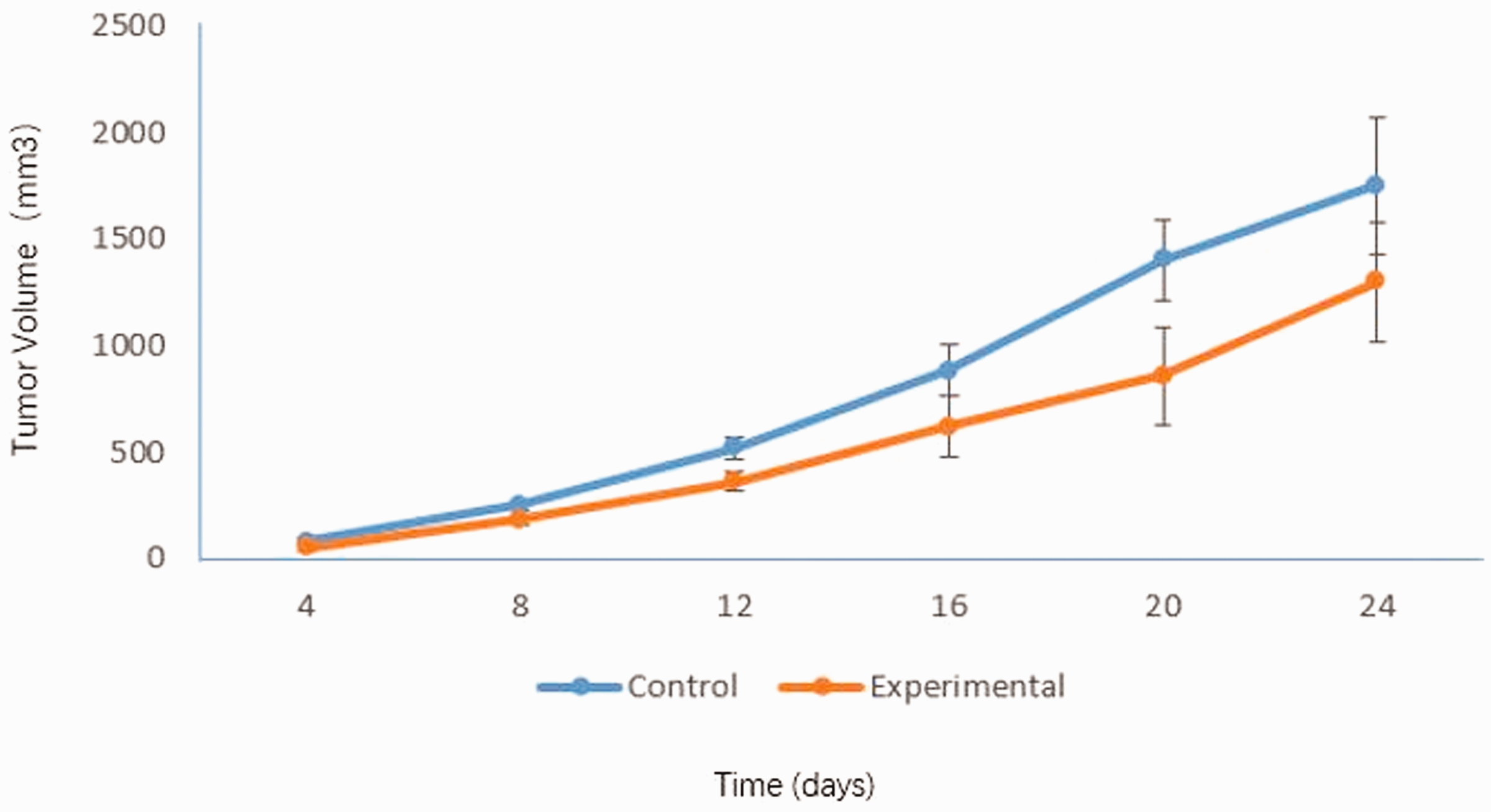
SP600125 treatment suppressed tumor growth during the 15-day observation period.
SP600125 downregulated JNK expression
The mRNA expression of JNK in SP600125-treated mice was evaluated using RT-PCR. As shown in Figure 3, JNK mRNA expression was significantly downregulated in tumor tissue following treatment with SP600125 compared with that in the control group (0.68 ± 0.03 vs. 0.98 ± 0.169, P < 0.05), indicating that SP600125 could block JNK expression.
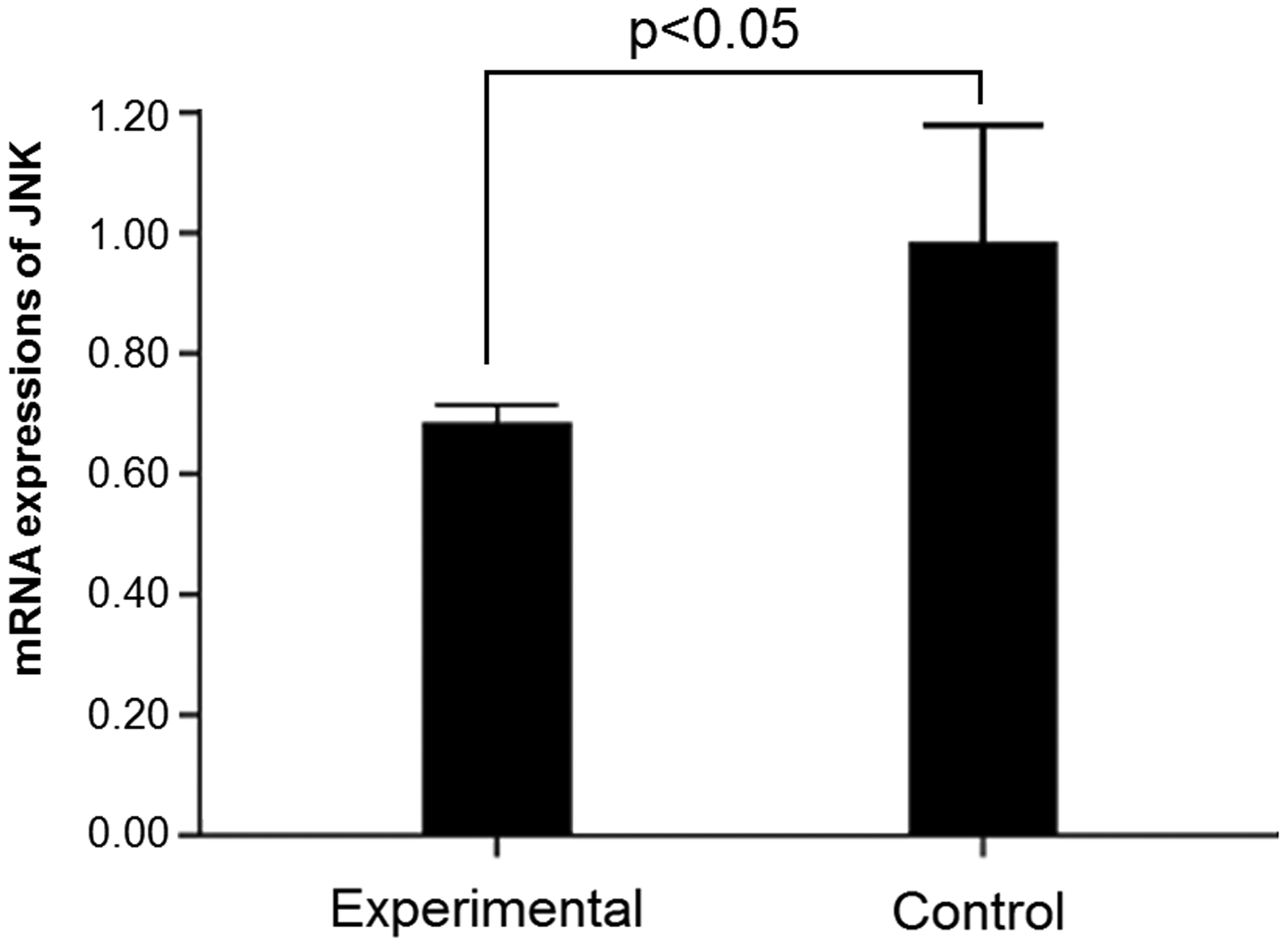
c-Jun NH2-terminal kinase mRNA expression in tumor tissue in mice. *P < 0.05.
SP600125 downregulated VEGF expression and decreased MVD
Images of immunohistochemically stained transplanted tumor tissues in the control and experimental groups are presented in Figure 4. The percentage of VEGF-positive cells was obviously reduced by SP600125 treatment (33.3%, P < 0.05, Table 2). Meanwhile, MVD was notably lower in the experimental group than in the control group (18.27 ± 1.70 vs. 23.17 ± 4.02, P < 0.05, Table 2 and Figure 5). In addition, the percentage of apoptotic cells was significantly higher in the experimental group than in the control group (10.23 ± 1.97% vs. 4.53 ± 1.40%, P < 0.01, Table 2 and Figure 6).

Vascular endothelial growth factor expression was examined via immunohistochemical staining in mice carrying transplanted breast cancer tumors (×400).
VEGF expression, MVD, and the apoptotic index in mice carrying transplanted breast cancer tumors.

*P < 0.05, **P < 0.01.
VEGF, vascular endothelial growth factor; MVD, microvascular density.
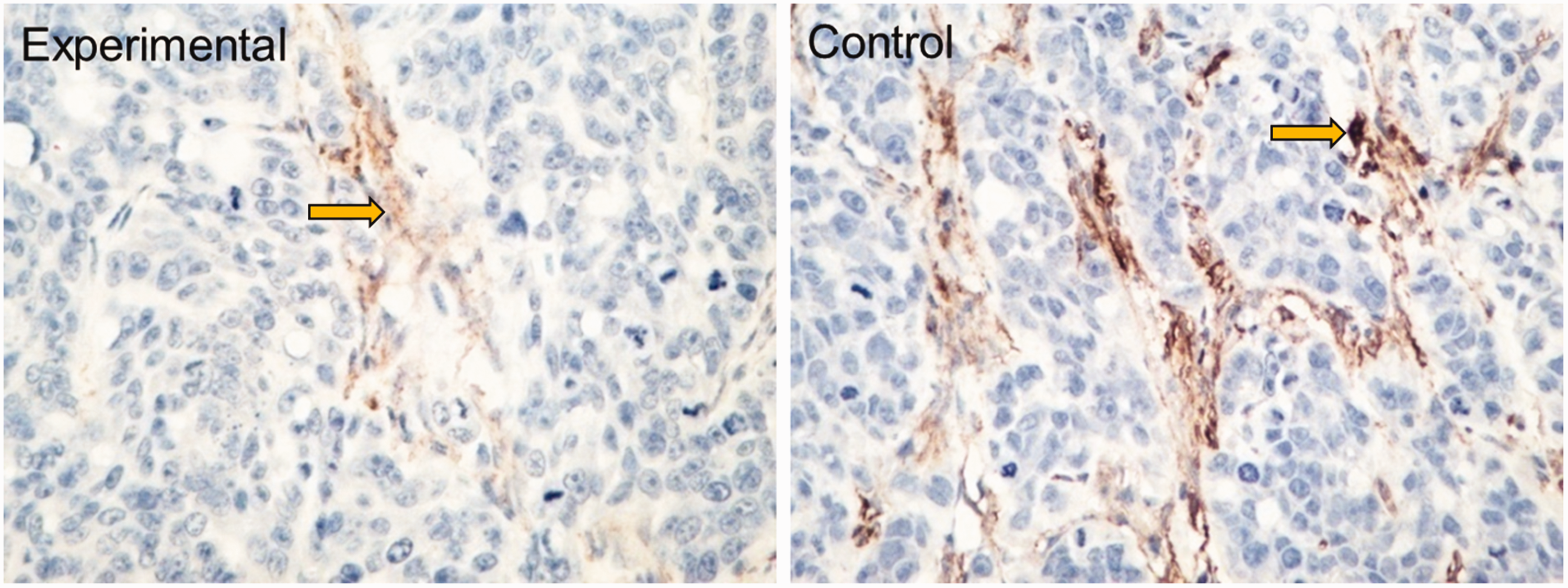
Microvascular density was examined via immunohistochemical staining in mice carrying transplanted breast cancer tumors (×400).
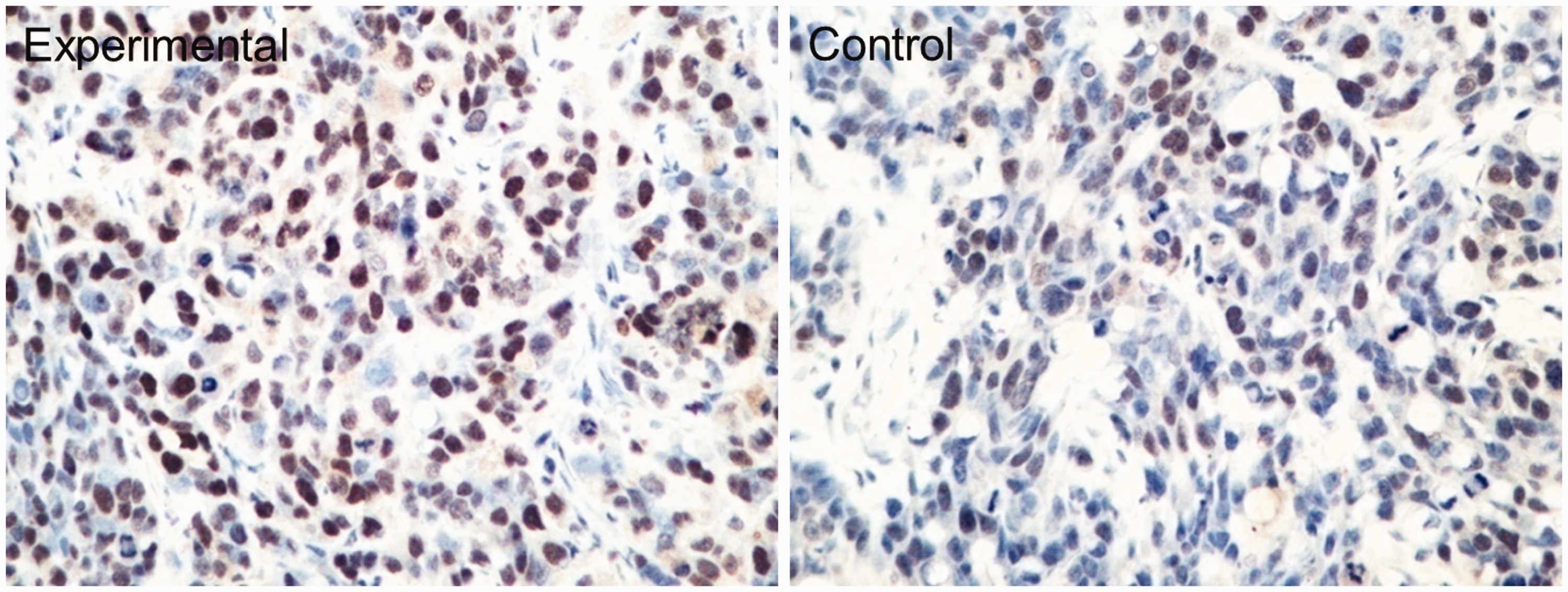
Apoptotic cells (brown yellow) were counted in mice carrying transplanted breast cancer tumors (×400).
Discussion
A number of studies have revealed that the JNK signaling pathway is excessively activated in breast and other cancers. This suggests that JNK signaling is closely related to the occurrence and development of breast cancer. 5 Karin et al. discovered the essential role of the JNK signaling pathway in breast cancer. The results demonstrated that the repression of inhibitor of κB kinase-beta (IKKβ) facilitates the activation of JNK following exposure to chemical carcinogens in mouse mammary adenocarcinoma cells mice, thus increasing the chance of carcinogenesis. 16 In addition, their study indicated that JNK gene deletion could prevent the occurrence of breast cancer and reverse the promotive effect on IKKβ deletion on tumorigenesis. Sakurai et al. 9 revealed that the JNK signaling pathway was excessively activated in a mouse breast cancer model induced by diethylnitrosamine (DEN), and the suppression of JNK signaling reduced the sensitivity of mammary adenocarcinoma cells to DEN. In our study, which used a mouse breast cancer model, we found that JNK signaling was significantly suppressed by SP600125 treatment in mammary adenocarcinoma cells.
The current study also revealed that JNK blockade could inhibit the growth of transplanted breast tumors in mice. Meanwhile, VEGF expression in tumor tissues and intratumoral MVD were decreased by SP600125 treatment, whereas mammary adenocarcinoma cell apoptosis was increased. These findings indicated that the JNK signaling pathway could increase MVD by upregulating VEGF expression, facilitate cell proliferation, and inhibit apoptosis, thereby facilitating the growth of breast cancer. However, Kennedy et al. 17 reported that the inhibition of JNK signaling accelerated the growth and increased the number of tumors in mice. It was also revealed that the activation of JNK signaling induced apoptosis in gastroenteric tumors. 18 At present, the mainstream view still considers that the JNK signaling pathway exerts a positive effect on the growth of breast cancer, in line with our conclusion.
The detailed mechanism of the effect of JNK signaling on cancer remains unclear. Some studies found that cellular stress and inflammatory factors usually increase the stability of COX-2 mRNA in endothelial cells via the MAPK pathway, thereby catalyzing the synthesis of PGE2.5,19 PGE2 is highly expressed in tumor tissues, and it plays an important role in the pathological processes of tumor development and spread. It has also been reported that PGE2 has close relationships with the size, stage, metastasis, and other clinical characteristics of tumors.20,21 As an important component of the MAPK pathway, the JNK pathway is extremely important for cellular stress and the migration, proliferation, differentiation, and death of cells. Therefore, we speculate that JNK signaling pathway might influence the occurrence and development of tumors by upregulating PGE2 expression in local tissues of the body. These mechanisms should be studied in future research.
The JNK signaling pathway plays vital roles in cell differentiation, apoptosis, stress reaction and the occurrence and development of a variety of human diseases. Thus, this pathway can be considered an important regulatory target for normal and diseased states of cells. The compensatory proliferation of cells mediated by JNK has an important role in the occurrence of breast cancer, and thus, inhibition of JNK signaling can significantly repress the growth of mammary adenocarcinoma cells. The JNK signaling pathway represents a potential molecular target for clinical tumor treatment.