
Abstract
Objective
Retrospective, observational study to explore the role of dendritic cells (DCs) in condyloma acuminatum lesions (genital warts) and their relationship with duration of the disease.
Methods
Condyloma acuminatum lesion samples were collected from male patients with the condition and compared with normal foreskin samples from male volunteers. Cellular locations of dendritic cell lysosome-associated membrane protein (DC-LAMP) and dendritic cell-specific intercellular adhesion molecule-3 grabbing nonintegrin (DC-SIGN) were detected using immunohistochemistry. Levels of both proteins were determined using Western blot analysis; levels of their corresponding mRNAs were measured using reverse transcription–polymerase chain reaction.
Results
The mRNA and protein levels of DC-LAMP and DC-SIGN were both significantly higher in condyloma acuminatum lesions (n = 30 samples) compared with normal skin samples (n = 13). Levels of DC-LAMP and DC-SIGN protein and duration of disease were inversely correlated.
Conclusions
DC-LAMP and DC-SIGN may be involved in the pathogenesis of condyloma acuminatum. Their levels were inversely correlated with the duration of disease, suggesting that DCs might be involved in human papillomavirus clearance.
Keywords
Introduction
Condyloma acuminatum, also known as external genital warts, is a common sexually transmitted disease caused by human papillomavirus (HPV). 1 It is characterized by a long disease duration and frequent recurrence, which may be associated with the reduction of local immunity. 2 Coleman et al. 3 demonstrated a large amount of T cells infiltrating regressing condyloma acuminatum lesions; CD4+ T cells predominated in regression and there was a significant chïange in the CD4+/CD8+ cell ratio. These changes in T-cell populations might be closely related to local immune disorders in condyloma acuminatum lesions. To date, dendritic cells (DCs) are considered to be the most powerful antigen-presenting cells, and are the only ones that can activate naïve T lymphocytes. 4 Functional alterations of DCs (such as migration into inflammatory tissues, change of maturation status and antigen-presenting ability) will lead to abnormal immune states. 5 Thus, the biological behaviour of DCs in the pathogenesis of condyloma acuminatum could be extremely important.
Dendritic cell lysosome-associated membrane protein (DC-LAMP or CD208) is considered to be a molecular marker of DC maturation. 6 It is exclusively located on the endomembranes of lysosomes in mature DCs, and participates in the synthesis and intracellular transportation of the major histocompatibility complex (MHC)–antigen complex of DCs. 5 DC-specific intercellular adhesion molecule-3 (ICAM-3) grabbing nonintegrin (DC-SIGN, or CD209) serves as a pattern-recognition receptor and an adhesion receptor to regulate transmembrane migration, antigen phagocytosis and immune function of DCs. 7 DC-LAMP and DC-SIGN play important roles in the antigen presentation of DCs and the stimulation of T-cell activation and differentiation.6,7
The present study compared levels of DC-LAMP and DC-SIGN protein and mRNA in condyloma acuminatum lesions and normal skin tissue samples, in order to determine whether there were any correlations between levels of DC-LAMP and DC-SIGN in condyloma acuminatum lesions and disease duration.
Patients and methods
Study Population
All tissue samples used in this retrospective observational study were obtained from consecutive patients with condyloma acuminatum treated at the Department of Dermatology, Qilu Hospital, Shandong University, Jinan, Shandong Province, China between January 2009 and June 2010. Clinical features and the results from pathological examinations were used as diagnostic criteria for condyloma acuminatum. In order to avoid the effects of oestrogen and progesterone, all patients were men. Subjects with factors that may have a potential impact on the comparability among different subjects, e.g. other diseases and the use of systemic or external medication, were excluded. The warts of all samples were treated with liquid nitrogen, except those surgically removed for analysis in this study. All patients were asked to make return visits every 2 weeks, to record the duration of condyloma acuminatum.
Between January 2009 and June 2010, normal foreskins were surgically removed from male patients with redundant prepuce who were being treated at the Department of Urology, Qilu Hospital, Shandong University. Age and sample location of the normal control tissues were matched with those of the patients.
The study was approved by the Medical Ethics Committee of Qilu Hospital. All subjects had signed the written informed consent form before the study commenced.
Tissue Preparation
Each tissue sample (condyloma acuminatum or normal foreskin), was divided into two parts: one part was used for extraction of total RNA (described below); the other was preserved in 10% neutral formaldehyde solution. All samples were frozen, stored in liquid nitrogen and tested within 1 month following collection. All samples were tested by an investigator (L.Z.) who was blinded to the study samples.
Immunohistochemical Analysis of DC-LAMP Proteins
Tissue sections (3-µm thick) were prepared from formalin-fixed, paraffin wax-embedded specimens, then deparaffinized in xylene and rehydrated in a graded series of alcohol. Antigen retrieval was carried out by heating sections in a 400-W microwave in 0.1 M ethylenediaminetetra-acetic acid, pH 8.0, at 97°C for 5 min, then cooling them to room temperature. Endogenous peroxidase activity was blocked by incubation in 3% H2O2 for 20 min at room temperature. Sections were then incubated with mouse antihuman DC-LAMP monoclonal antibody (1 : 300 dilution; Imgenex, San Diego, CA, USA) at 4°C overnight, or with mouse antihuman DC-SIGN monoclonal antibody (1 : 400 dilution; R&D Systems, Minneapolis, MN, USA) at 37°C for 90 min. The slides were washed three times in 0.1 M phosphate-buffered saline (PBS), pH 7.4, and undiluted horseradish peroxidase goat antimouse secondary antibody was applied, according to instructions provided by the manufacturer of the EnVision™ Kit (DAKO, Glostrup, Denmark), for 20 min at 37°C. The slides were washed again three times in 0.1 M PBS, pH 7.4, and diaminobenzidine (DAB) was used as the chromogenic reagent. Sections were counterstained using haematoxylin and eosin, differentiated in hydrochloric acid-ethanol, blued in ammonia water, dehydrated in an ethanol gradient, cleared with xylene, mounted in neutral balsam and examined under a light microscope. Normal tonsil tissues obtained from the Department of Otorhinolaryngology, Qilu Hospital, Shandong University, were used as positive controls, and study samples treated with isotype control immunoglobulin were used as negative controls.
Western Blot Analysis of DC-LAMP and DC-SIGN Proteins
For Western blot analysis, a frozen sample (2 g) of tissue was homogenized in 10 ml of lysis buffer (Beyotime, Shanghai China) for 24 h at 4°C. The total protein in the supernatant was separated by centrifugation at 2700
RNA Extraction and RT–PCR Analysis
Total RNA was extracted from skin biopsy samples (5 g) by employing the isothiocyanate method using TRIzol® (Qiagen, Valencia, CA, USA) according to the manufacturer’s instructions. Following assessment of the RNA quality by using the absorbance ratio of A260/A280, RNA samples were denatured. The RNA reverse transcription kit and PCR reaction kit were supplied by Thermo Fisher Scientific (Rockford, IL, USA); all other reagents used in the experiments were obtained from Sigma-Aldrich (St Louis, MO, USA). The RT reaction was carried out in a volume of 10 µl, containing 2 µl of 5 × RT buffer, 0.5 µl of RT enzyme mix, 0.5 µl of primer mix, 500 ng RNA, and nuclease-free water. cDNA was synthesized by incubation of the samples at 37°C for 15 min then for 95 min at 8°C. The PCR reaction was carried out in a volume of 50 µl, containing 25 µl of 2 × Quick Taq HS Dye Mix, 1 µl primer (each), 5 µl cDNA, and sterilized deionized water. The three pairs of primers (DC-LAMP; 5′-ACCTTACAGGGAAATGGG-3′, 3′-AAGAACAGTTGGTTAGCG-5′; DC-SIGN: 5′-TCTCCTGGCTCTTTGACA-3′, 3′-CCACCACGATGAATACTACA-5′; β-actin: 5′-CGGGAAATCGTGCGTGAC-3′, 3′-TGGAAGGTGGACAGCGAGG-5′) were synthesized by Sangon Biotech (Shanghai) Co. Ltd. (Shanghai, China). The cycle parameters were set as follows: a preliminary denaturation step at 94°C for 2 min, followed by 28 cycles of denaturation at 94°C for 30 s, annealing at 72°C for 30 s, polymerization at 68°C for 1 min, and a final extension step at 72°C for 10 min. The amplified PCR products were analysed by 2% agarose gel electrophoresis at 100 V for 40 min. Gel images were observed using an ultraviolet imaging system. The optical density of the bands, as well as the ratio of the optical densities (target gene/β-actin), was calculated using a gel imaging system (ChemiImagerTM 5500; Alpha Innotech). DC-LAMP mRNA was detected at 260 base pairs (bp), DC-SIGN at 127 bp, and β-actin at 343 bp.
Statistical Analyses
Data were analysed using the SPSS® statistical package, version 17.0 (SPSS Inc., Chicago, IL, USA) for Windows®. Data were presented as mean ± SD. Differences between groups were compared using an independent-samples t-test. Statistical significance was assumed as a P-value <0.01. The correlation between levels of DC-LAMP or DC-SIGN mRNA in condyloma acuminatum lesions and duration of condyloma acuminatum was analysed by Spearman’s rank correlation coefficient analysis; a P-value <0.05 was considered statistically significant.
Results
Samples of condyloma acuminatum lesions were obtained from 30 male patients with condyloma acuminatum; mean age, 27.8 years (range 19–45 years). A total of 13 normal foreskins were surgically removed from male patients with redundant prepuce (mean age 30.7 years, range 18–45 years).
In normal foreskin tissue samples, immunohistochemical staining of the DC-LAMP (Figure 1A) and DC-SIGN (Figure 1C) proteins was located in the membrane and cytoplasm of a few DCs at the basal and spinous layers of the epidermis, and superficial layer of the dermis. In condyloma acuminatum lesions, large amounts of DC-LAMP (Figure 1B) and DC-SIGN (Figure 1D) proteins were mainly located in the lower parts of the wart tissues and the superficial layer of the dermis, as indicated by staining of the cytoplasm and cytomembrane of DCs.
Immunohistochemical staining of dendritic cell lysosome-associated membrane protein (DC-LAMP) in (A) normal foreskin tissue and (B) condyloma acuminatum lesion; arrows show DC-LAMP in epidermis and dermis. Immunohistochemical staining of dendritic cell-specific intercellular adhesion molecule-3 grabbing nonintegrin (DC-SIGN) protein in (C) normal foreskin tissue and (D) condyloma acuminatum lesion; arrows show DC-SIGN in epidermis and dermis.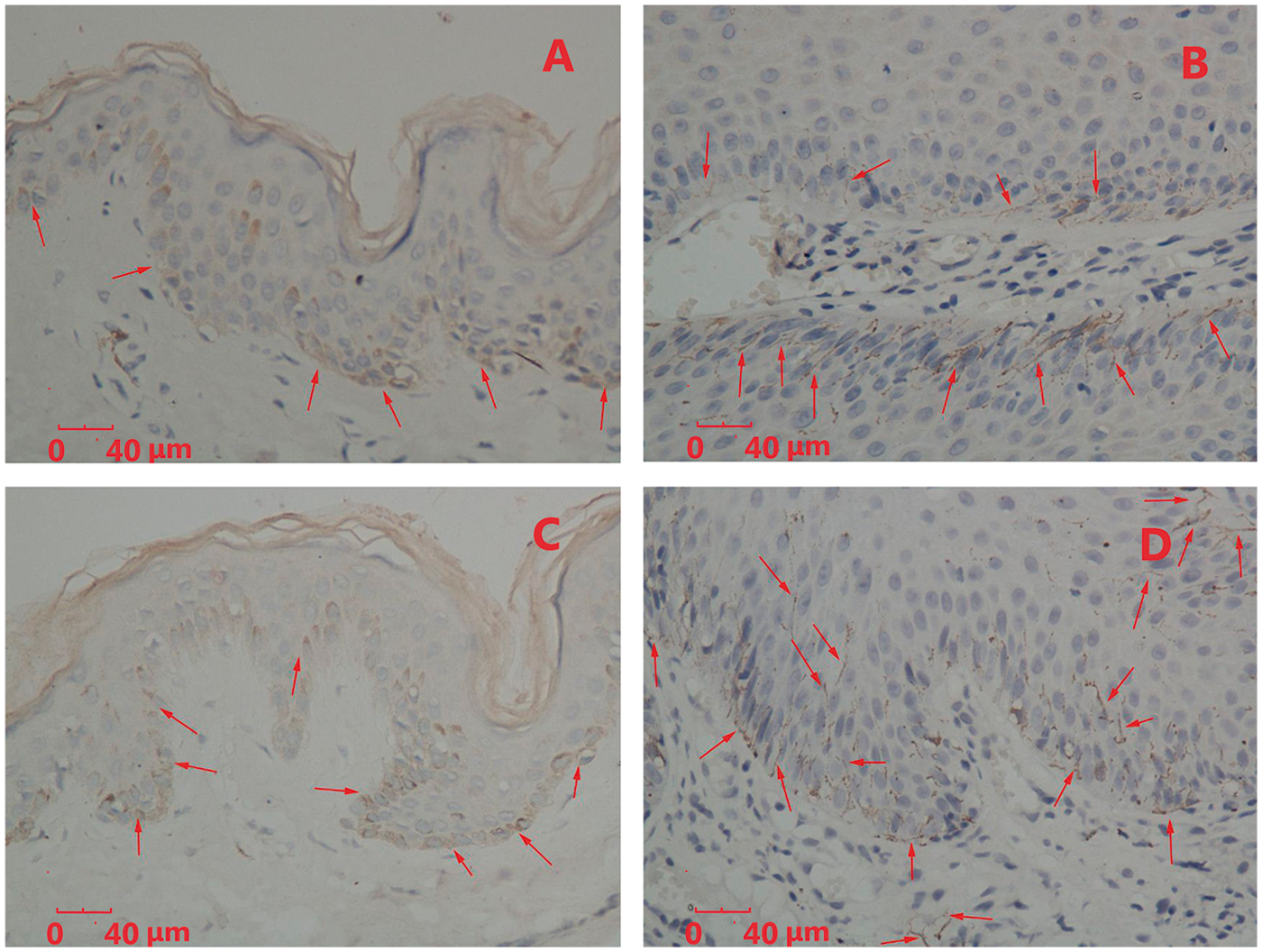
Using Western blot analysis, bands of 44 kDa corresponding to the DC-LAMP and DC-SIGN proteins were shown in normal foreskin tissue samples and condyloma acuminatum lesions. Analysis of the Western blots revealed that the mean ± SD grey value ratio for DC-LAMP (1.33 ± 0.34) and for DC-SIGN (1.57 ± 0.53) was significantly higher in condyloma acuminatum lesions than in normal foreskin tissue samples (mean ± SD 0.34 ± 0.12 for DC-LAMP and 0.47 ± 0.26 for DC-SIGN) (P < 0.01 for both comparisons).
The mean ± SD optical density ratio for DC-LAMP mRNA (3.52 ± 1.04) and for DC-SIGN mRNA (2.99 ± 0.94) was significantly higher in condyloma acuminatum lesions than in normal foreskin tissue samples (mean ± SD 1.54 ± 0.51 for DC-LAMP and 0.94 ± 0.49for DC-SIGN) (P < 0.01 for both comparisons).
The duration of condyloma acuminatum ranged between 3 and 18 months (mean ± SD 9.40 ± 4.34 months). There were significant inverse correlations between levels of DC-LAMP (r = –0.52, P < 0.05) and DC-SIGN (r = –0.56, P < 0.05) in condyloma acuminatum lesions and duration of disease.
Discussion
Condyloma acuminatum is always characterized by a long disease duration and frequent recurrence. Its cause is closely related to reduced levels of local immunity. 2 Cellular immunity mediated by T cells is regarded as the primary means by which the immune system provides protection against HPV infection. 8 During the initial stage, HPV infects primitive basal keratinocytes and these cells are activated to produce high amounts of chemokines, cytokines, and growth factors, which cause genital warts. 9 DCs (such as Langerhans cells) take up and process undefined antigens, then migrate to draining lymph nodes where they present the antigen to activate CD4+ T cells (T-helper cells). After activation, CD4+ T cells secrete chemokines and cytokines to regulate the functions of other immune cells. 8 Moreover, these T cells enter the dermis and help CD8+ T cells to recognize and lyse the infected keratinocytes. 10 In this process, DCs play key roles in activating T cells and assisting the function of other immune cells.
It has been shown that the number of T cells was significantly higher in condyloma acuminatum than in normal skin tissues, and that the ratio of CD4+ T cells to CD8+ T cells decreases in the epidermis and dermis in condyloma lesions. 3 This indicates that the infiltration and changes in T-cell subsets are highly associated with a local immune disorder. 11 Having considered the findings described above, in our opinion, changes in the numbers and subsets of T cells, and the transformation of the activation state of DCs, might be inseparable in the development of condyloma acuminatum lesions.
The present study demonstrated that DC-LAMP and DC-SIGN were mainly distributed at the bottom of lesion tissues and at the superficial layer of the dermis in condyloma acuminatum. As DCs migrate from peripheral blood to draining lymph nodes to present antigens to CD4+ T cells, 10 the current study suggests that the recruitment, antigen-presenting capacity and migration of DCs might be activated in condyloma acuminatum lesions.
Dendritic cell-specific intercellular adhesion molecule-3 grabbing nonintegrin, also known as CD209, is a type II transmembrane protein with a molecular weight of 44 kDa, belonging to the superfamily of C-type exogenous lectins. 12 It is divided into three regions: (i) an N-terminal domain located in the cytoplasm without signal peptide, whose main function may be related to DC endocytosis and signal transduction; (ii) a transmembrane region with seven complete and one incomplete repeat domains, which greatly enhances the affinity between DC-SIGN and its ligands, and amplifies the signals; (iii) an extracellular region, containing a carbohydrate recognition domain, which is a specific sequence of C-type exogenous lectin and the major binding site of ligands, which can bind two Ca2+ molecules and one mannose molecule.13–15 DC-SIGN is mainly expressed on the surface of immature DCs in peripheral tissues and on mature DCs in lymphoid tissues, but not in naïve T cells, granulocytes, thymus cells, CD4+ bone marrow cells or activated T cells. 12 The physiological functions of DC-SIGN include two aspects: recognition of pathogens by receptors and cell adhesion. 16 DC-SIGN on the surfaces of peripheral immature DCs can interact with ICAM-2, which is highly expressed in the vascular and lymphatic endothelium, in order to facilitate these DCs to enter lymph nodes and other immune organs. 17 Thus, antigens can be captured, engulfed and then processed by DCs into MHC–antigen complexes in the lysosomes, which are presented to T cells.18,19 DC-LAMP, the DC maturation marker, is expressed in these mature DCs. 5 DC-SIGN on the surface of mature DCs can also bind ICAM-3, which is highly expressed on naïve T cells, to form a stable contact between DCs and T cells. 20
The present study showed that mRNA and protein levels for DC-LAMP and DC-SIGN were low in normal skin, but high in condyloma acuminatum lesions. This is possibly due to a small amount of resident DCs being present in normal skin; these are mainly immature Langerhans cells that provide an immune surveillance function. 21 Levels of DC-LAMP and DC-SIGN are so low in these Langerhans cells that activation of T cells rarely occurs. During HPV infection, however, large numbers of DCs are recruited and matured in order to take up, process and present antigens to naïve T cells. 22 DC-LAMP and DC-SIGN facilitate the activation of a high number of T cells. 23 The infiltration of T cells and changes in the quality of T cell subsets occur in response to disorder of local cell-mediated immunity. 3
Statistical analysis revealed that the mRNA levels of DC-LAMP and DC-SIGN were inversely correlated with the duration of condyloma acuminatum. According to the view that naïve T cells are activated by DCs via DC-LAMP and DC-SIGN, 24 it can be hypothesized that these DCs, expressing high levels of DC-LAMP and DC-SIGN in the lesions of condyloma acuminatum, have a strong capacity for antigen processing and presentation. Their ability to activate T cells and stimulate the immune response is very powerful. Once the interaction between DCs and T cells occurs, antigen-specific cellular and humoral immune responses play important roles in defending against HPV infections, whose clearance is very rapid and effective. As a result, the disease duration in patients with high levels of DC-LAMP and DC-SIGN protein might be expected to be shorter than that in patients with low levels.
In conclusion, levels of DC-LAMP and DC-SIGN were higher in condyloma acuminatum lesions than in normal foreskin tissue samples, which indicated that the two proteins may be involved in the pathogenesis of condyloma acuminatum. The mechanism might be strongly associated with antigen presentation by DCs and activation of T cells. In addition, the mRNA levels of DC-LAMP and DC-SIGN were inversely correlated with the duration of condyloma acuminatum, suggesting that DCs might be involved in HPV clearance.
Footnotes
Declaration of Conflicting Interest
The authors declare that there are no conflicts of interest.
Funding
This project was supported by a grant from Shandong Province Natural Science Foundation (No. ZR2010HM018).