
Abstract
Choromytilus chorus virtually vanished from most Peruvian archaeological sites about 1000 BC. This species’ disappearance is significant because Choromytilus chorus is one of the most recognizable mollusks in the archaeological remains of coastal Peru because of its large size and rich purple color. In this study we investigate the choro mussel, which was a highly valued food source, a raw material for fishhooks and other fishing gear, and a special ritual item in prehistoric times. We look at inconsistencies in reports of its range, habitat, and ecology to explore reasons for the major shift southward of the species' northern limits at the end of the Initial Period (1000 BC), a significant cultural boundary. We identify biological, ecological, and cultural factors that acted together to bring about the sudden extinction of the choro mussel along much of the Peruvian coast. Paramount among these are human overexploitation and periodic environmental changes related to the El Niño phenomenon.
Plain Language Summary
The large Pacific mussel, Choromytilis chorus, disappeared from the archaeological record along much of coastal Peru around 1000 BC. A significant reduction in shell size at archaeological sites over time suggests that human overexploitation was the main cause of changes in C. chorus’ northern range. Later, El Niño events further hindered the mussel’s ability to recover.
Introduction
Choromytilus chorus has long attracted the attention of biologists, malacologists, archaeologists, and shellfish harvesters and cultivators because of its distinctive color, large size, and value as both a food species and raw material for tools. This mussel is commonly known as the purple mussel in Peru and the black mussel, giant mussel, or choro zapato in Chile (Guzman, Saá, and Ortleib 1998; Ibarrola et al. 2012; López, López, and González 2008; Sandweiss and Rodríguez 1991). For purposes of consistency, the common name “choro mussel” is used here to adhere to SeaLifeBase conventions (SeaLifeBase 2020). Although it is easily recognized and frequently mentioned in archaeological and other scientific literature, C. chorus has remained something of an enigma because of persistent inconsistencies in reporting the species’ range, habitat, and ecology.
Numerous investigators working with archaeological shellfish data have observed that the northern limit of C. chorus has shifted south over time (Bird 1948; Chu 2008; Elera 1998, 274; Pozorski 1976a; Sandweiss, Maasch, and Kelly 2024; Sandweiss et al. 2001; Strong and Evans 1952). Efforts to explain this southward shift of C. chorus have focused on climate and oceanographic conditions, especially the El Niño phenomenon (Avendaño and Cantillánez 2011; Elera 1998, 274; Sandweiss et al. 2001); however, other factors are also significant. Malacologist Luis Ramorino of the University of Chile noted that when a species is not found within its previously-determined range, its absence may be due to changes in ecological conditions as well as overexploitation or an ancient error in identification (Ramorino 1974).
This study begins with a description of this species' importance to prehistoric societies along the western coast of Peru and Chile. To better understand and explain variations in the choro mussel' s range and habitat, we also critically examine published accounts of both of these components of C. chorus ecology with a focus on how inconsistencies in reporting might have occurred. To identify and evaluate possible natural causes for the observed southward shift in C. chorus' range over time we consider environmental features, including broad ENSO events and more localized water conditions, as well as biological characteristics related to C. chorus’ growth and development. Finally, as part of addressing the southward shift, we also use archaeological data to address the potential effects of human exploitation of the choro mussel. Our results suggest that the shrinking range of C. chorus may be explained by a combination of interacting ecological, biological, and cultural factors.
C. chorus virtually disappeared along the north and central coast of Peru at the end of the culturally defined Initial Period around 1000 B.C. By archaeological standards, this was a sudden event. Such a coincidence is a strong indication that human factors are significant. The large-scale extinction documented here was possible because the Andean cultural area, centered in north-central Peru, was the locus of highly cohesive cultural development (Pozorski and Pozorski 1987b). This cohesion is evident as early as the Late Preceramic Period (3000-1800 B.C.), when permanent settlements, including some with sizable platform mounds, were established along the coast and within some Peruvian valleys. Communication among communities is reflected in shared technologies (e.g., twined textiles and mussel-shell fishhooks) prior to the use of ceramics. This interaction continued to ensure a degree of cultural uniformity within the Andean core through the Initial Period (1800–1000 B.C.) when large inland sites dominated by platform mounds were established well inland near irrigated lands while sizable coastal settlements continued to supply essential marine resources (Burger 1992, 57–58; Pozorski and Pozorski 1979).
Human use of C. chorus
Archaeologists working along the coast of South America, especially investigators excavating preceramic and early ceramic sites, have long recognized C. chorus as a major food source. In addition, this distinctive bivalve was useful as raw material for artifacts and seemed to have had an intrinsic special significance.
Even today, choro mussels are a favorite food in Chile; and their presence in substantial shell middens along the coasts of Peru and Chile attest to their popularity as food in prehistoric times. Mussels of this species from refuse with food debris are usually very fragmented, evidence that they were roughly broken open and probably eaten raw rather than steamed (Figure 1).

Archaeological specimens of C. chorus including whole examples as well as umbos and fragments resulting from breaking the shells to extract the meat. Scale measures 5 cm.
Early inhabitants of Peru and Chile used C. chorus valves as raw material for tools, pendants, and other items of worked shell; and occasionally whole shells were used as expediency scraping tools (Julien and Vanhaeren 2012). Nevertheless, most tools made from C. chorus valves were fishing gear, especially fishhooks, line weights, and occasionally barbs for composite hooks (Ballester, Clarot, and Bustos 2014; Bird 1943; Fung 1969, 105–113; Silva and Bahamondes 1969). Evidence of shell fishhooks and weights is concentrated along the northern coast of Chile and the central and north-central coast of Peru as far north as Las Haldas (Fung 1969, 99, 101–102). The abundance of shell fishhooks along the north coast of Chile led Bird to define the initial occupation there as a Shell Fishhook Period and Culture spanning the coastal zone from Arica to Taltal (Bird 1943, 307).
Archaeological evidence also suggests that C. chorus was ‘special’, possibly because of its size and deep near-black purple color (Sandweiss 1982; Sandweiss and Rodríguez 1991). Numerous discoveries document examples of C. chorus in possible ritual contexts. Gabriel Prieto (2015, 140, 204) describes contexts at the coastal Initial Period site of Gramalote in the Moche Valley where C. chorus valves received special treatment: as a ritual cache of 14 valves on a reed mat and as a votive offering of seven valves within public architecture. Chen (2019) describes 20 large C. chorus valves capping an infant burial at the site of Huaca Negra in the Viru Valley. We recently discovered a profile face carved from C. chorus shell at the inland site of Taukachi-Konkán in the Casma Valley (Figure 2).

Profile face carved from C. chorus recovered from Casma Valley archaeological site of Taukachi-Konkán. Scale measures 5 cm.
Choro mussel shells were also often associated with red pigment. Examples are known from the Middle Preceramic Ring Site in southern Peru (Sandweiss and Rodríguez 1991) and Cahuachi in the Nazca Valley (Silverman (1993, 295). The Public Architecture at Gramalote also yielded 22 C. chorus valves with red pigment residue, indicating these mussel shells may have been used as vessels to mix red paint (Prieto 2015, 1033–1034). Additional evidence of the intrinsic value of C. chorus comes from its presence at sites far from its source, an indication of curation and long-distance trade. Large valves of the choro mussel were recovered within the Galería de las Ofrendas at Chavín de Huantar in the central Peruvian highlands (Sandweiss and Rodríguez 1991). Flannery and Marcus (Marcus 2016), working at the south coast site of Cerro Azul, recovered a single large whole C. chorus valve that they felt was probably imported from Nazca by noble women to serve as a pigment dish.
Defining the Range of C. chorus
The geographic range or distribution for C. chorus is variously described by malacologists as extending from Pacasmayo, Peru south to the Straits of Magellan (Guzmán, Saá, and Ortleib 1998, 62–63; Reid and Osorio 2000, 131–132) or from Callao, Peru south to the Straits of Magellan (Osorio 2002; Gobierno de Chile 2010, 37). In contrast to these more fixed range estimates, abundant archaeological data and some geological data indicate that the northern range limit of C. chorus has shifted significantly southward over the past several millennia. This southward range movement has also been documented for several prized food species often found in association with C. chorus, including Semele sp., Eurhomalea rufa, Leukoma thaca, Tellina sp., and Mesodesma donacium (Prieto 2015; Sandweiss et al. 2001).
Evidence for the presence of C. chorus in far northern Peru, between Puerto Pizarro and Zorritos, extends back into the Pleistocene, well before significant human occupation of the coastal zone (Ortleib and Diaz (1991). Based on more recent archaeological evidence, we know that the northern range of C. chorus extended north beyond Pacasmayo in early prehistoric times at least as far as the coast of the Lambayeque Valley (Figure 3 1 ). Choro mussels are the most abundant species within the molluscan inventory at Ventarrón, a Preceramic site on the north side of Reque River (Alva and Alva 2012). Farther south, excavations at the site of Puemape, located on the coast between the Chicama and Jequetepeque Valleys, also yielded substantial amounts of C. chorus (Elera 1998, 164–165). Assertions that the modern range of C. chorus extended as far north as Pacasmayo in Peru are probably based on the presence of the choro mussel in coastal archaeological sites such as Puemape which lies very near the water's edge about 15 km south of Pacasmayo. Erosion of these coastal archaeological sites during very high tides could easily scatter a beach with prehistoric midden-derived shells.
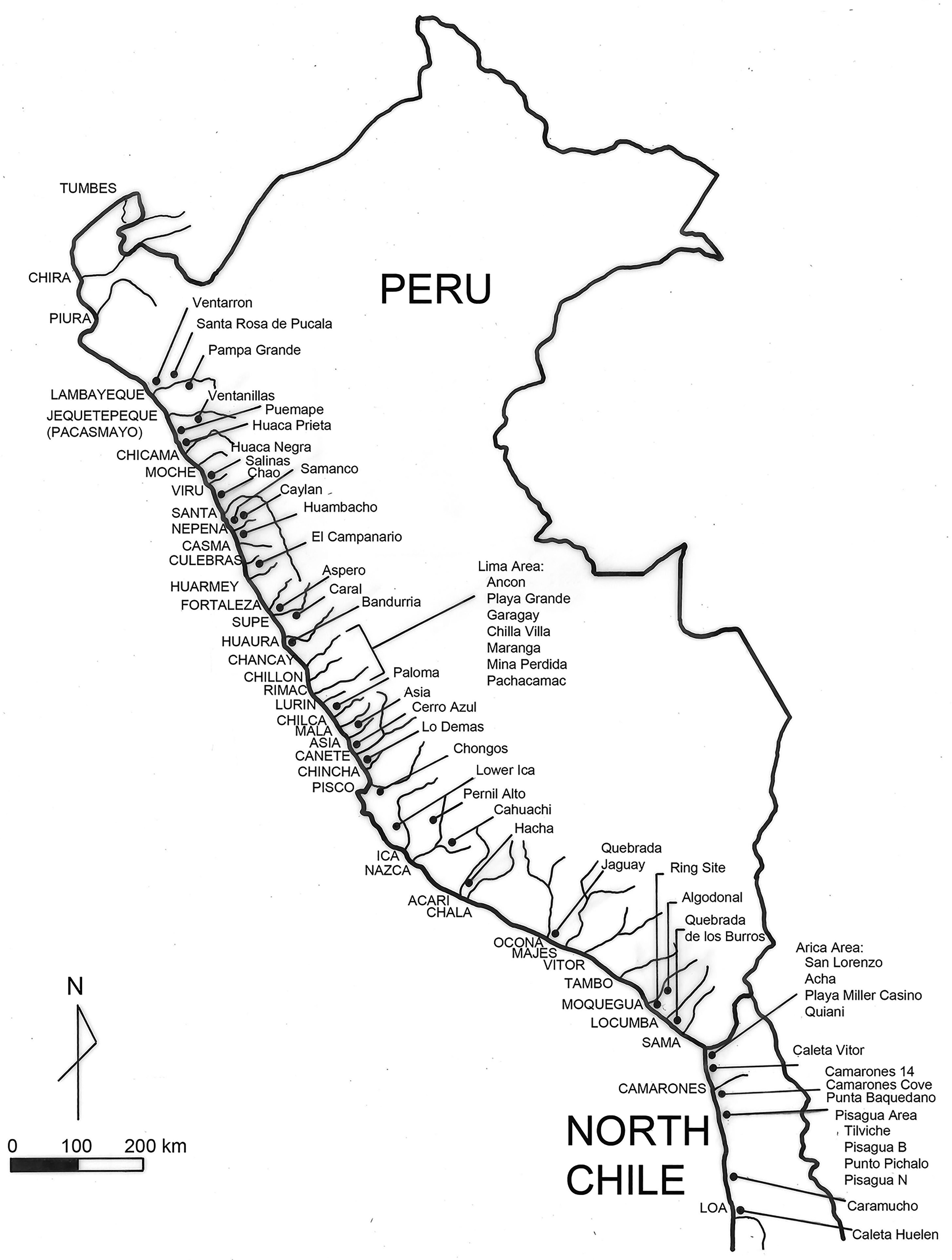
Map of Peru and northern Chile indicating river valleys (upper case) and archaeological sites (upper and lower case) mentioned in text and Tables 1-3.
Twentieth-century accounts document living C. chorus populations along southern and central Peru during the early part of the twentieth century. Dall describes a molluscan collection made by Robert Ervin Coker from 1906 to 1908 that documents the range for C. chorus as extending from Coquimbo in north-central Chile northward to Independence Bay near the Ica Valley in southern Peru (Dall 1909, 147).
More recently, Violeta Valdiviezo observed a small C. chorus population in the late 1970s at Santa Rosa, near Ancón; and archaeologist Carlos Elera collected live specimens in 1983 from Laguna Grande at the north end of Independence Bay adjacent to the Ica Valley (personal communications cited in Sandweiss 1992, 102). By 2016, C. chorus was again reported from southern Peru, along the coasts of Moquegua and Tacna (Tejada et al. 2020). These reports reveal a current northern range for the choro mussel that ends near Ancón, well south of the frequently proposed limit of Pacasmayo in northern Peru (Figure 3); and they provide clear evidence that this northern range limit has varied considerably through time.
Defining the Preferred Habitat for C. chorus
The depth at which C. chorus may be encountered has been variously described as ranging between the tide zone and 20 m (Avendaño and Cantillánez 2011; Gobierno de Chile 2010, 38; Osorio 2002, 124), however ranges extending much deeper to 100 m (SeaLifeBase 2020) and even 285 m (Cárdenas, Aldea, and Valdovinos 2008) have been reported. Published habitat data indicate that gregarious beds of C. chorus may occur on rocky, sandy, or mixed rock and sand substrates or within estuaries where they are often found covered by sand or mud (Avendaño and Cantillánez 2011; Benitez et al. 2018; Gobierno de Chile 2010, 38; Ibarrola et al. 2012; Moreno 1995; Murphy 1921; Navarro 1988). These variations in habitat descriptions, with their possible inconsistencies, led us to further scrutinize habitat data for C. chorus.
In his background discussion of mytilids of the Chilean coast, Moreno (1995) reminds us that their intense exploitation began in prehistoric times and continued to increase in intensity, especially in the mid-1900s when harvesting pressure almost caused the extinction of C. chorus (Carranza et al. 2009; Gobierno de Chile 2010; Moreno 1995). Furthermore, Moreno (1995) points out that “the ecology of low intertidal environments in Chile has [subsequently] been studied in the absence of this mussel” [C. chorus]. This should make us especially cautious in our assessment of contemporary habitat data for the choro mussel.
With this in mind, we looked for examples of in situ C. chorus populations that closely approximated their natural state prior to significant human interference. During his 1921 visit to Peru, explorer and naturalist Robert C. Murphy visited Independence Bay, a coastal zone south of Pisco which at that time was “70 miles away by water [from any human settlements] and utterly inaccessible by land” (Murphy 1921, 49). Especially pertinent to this study is his description of the local mussel population:
Moreno (1995) studied a C. chorus population at a Chilean marine reserve. He describes this protected population as follows: Between the physical limits imposed by wave action, in rocky habitats covered seasonally by sand, a large population of the bivalve C. chorus (Molina 1782) recruited and grew at the Marine Reserve of Mehuín (Chile) after this site was closed to human activities. The re-establishment of this valuable edible mussel in the low intertidal zone occurred mainly because the area was protected from shellfish harvesters, since 1978. (Moreno 1995)
A 2010 study within the Reserva Marina para el Choro Zapato Putemún in southern Chile by the Servicio Nacional de Pesca describes C. chorus beds within an estuary zone. Near the southwest margin of the estuary, a large C. chorus bed covered an area of about 27,500 m2. Within this bed there were two aggregations; one with a density of 100 individuals per square meter and a second with 4 individuals per square meter (Gobierno de Chile 2010). Jaramillo, Bertrán, and Bravo (1992) describe beds of artificially farmed C. chorus mussels within the Queule River estuary of south Chile that attain densities of up to 250–300 adults per square meter.
Additional data relevant to this discussion of C. chorus habitat come from the study by Avendaño and Cantillánez (2011) in the Pisagua and Taltal areas of Chile. They observed that colonization of the bays by C. chorus resulted in the downward displacement of Aulacomya atra to a deeper habitat, enabling C. chorus to inhabit the shallower zone 4–13 m in depth while Aulacomya atra occupied a deeper zone 13 or more meters in depth (Avendaño and Cantillánez 2011).
Based on these accounts, C. chorus tends to occupy relatively shallow intertidal zones when human overexploitation is not a factor, even competing for this zone with another mussel species. This suggests that C. chorus’ current occupation of deeper habitat zones may be more a result of human exploitation than a reflection of its true natural habitat. Based on evidence that choro mussels in their true natural habitats readily occupy relatively shallow zones, including estuaries, very early inhabitants of coastal Peru and Chile would have had access to C. chorus in much more accessible habitats.
Environmental Factors Affecting C. chorus
El Niño or the ENSO phenomenon is paramount among environmental factors that potentially impacted C. chorus, especially with respect to variations in the species’ range. Factors such as water acidity and salinity have been examined, and C. chorus generally appears resilient when subjected to potential environmental hazards. Navarro (1988) found that C. chorus was well adapted to survive moderately reduced salinities. Núñez and Moragas (1978) correlated the abundance of C. chorus in early occupations at Tilviche sites in Chile with localized drainage of fresh water that decreased the ocean's salinity. When Benitez et al. (2018) subjected C. chorus to acidification, the choro mussel exhibited an ability to cope with increased acidity. In the studies by Navarro and Benitez et al., C. chorus resilience was attributed to the fact that the mussel was preadapted because of routine exposure to varying acidity and salinity in its natural habitat, especially estuarine zones and bays that receive freshwater runoff (Benitez et al. 2018; Navarro 1988).
Geologically ENSO events can result in strong wave action and flooding which can generate delta or beach-ridge zones near river mouths (Martínez-Villalobos et al. 2024; Odériz et al. 2020; Sandweiss, Maasch, and Kelly 2024). Such phenomena can both displace and generate habitats suitable for the formation of C. chorus beds.
Looking more closely at the impact of El Niño, Urban (1994) examined the upper temperature tolerance of ten species of bivalves from northern Chile and southern Peru. He assessed the time span and temperature at which 50% mortality (LT50) was reached. C. chorus was among eight species collected from central Chile; four species were also collected from southern Peru. Among the Chilean species studied, including C. chorus, lethal temperature values for most varied from each other by only 1.2°C. These data indicate that C. chorus shows about the same susceptibility to rising water temperature as other Chilean bivalves and is therefore not significantly more sensitive to this phenomenon than other species in the same zone. When data from two species sampled in both Peru and Chile were compared, their lethal temperatures measured about 3°C higher in Peru than Chile; and this difference was attributed to acclimation associated with more northern vs more southern sampling locales. Based on these comparative data, it is also possible that C. chorus populations previously established in Peru might have exhibited similar acclimatization to higher local water temperatures farther north. Urban concluded that all the bivalve species studied, including C. chorus, could tolerate temperature increases of about 5°C which characterize moderate El Niño events without facing danger of extinction (Urban 1994).
Empirically we would expect the impact of individual El Niño events to be reflected archaeologically primarily in the form of laminar sediments clearly derived from rainfall runoff. There may also be faunal changes reflecting the effect of El Niño on sea life. Mauricio (2012), working at the Early Intermediate Period site of Maranga in the Rimac Valley (Figure 3), documented rainfall-related laminar deposits from a strong El Niño event that occurred about A.D. 600. She also identified changes in the recovered shellfish that appear to correlate with expected responses to changes in water temperature. Contained within a single site, these data appear to document the relatively temporary effects of El Niño as shellfish populations weaken, die off, fall prey to predators, and ultimately begin to recover.
Archaeological data from our Casma Valley excavations also provide evidence regarding the ability of C. chorus to rebound after an El Niño event. Geologist Wells documented two ENSO events during her survey of the Casma Valley area that she was able to bracket in time between 3000 and 1500 B.C. (Wells 1988, 152). We believe that the later event coincides with an El Niño event we first noted at the Initial Period inland site of Pampa de las Llamas-Moxeke which was part of the larger Sechín Alto polity. We were able to estimate a date for this phenomenon of 1400 cal. B.C. based on radiocarbon evidence (Pozorski and Pozorski 2006). Subsequent excavations at the Initial Period coastal site of Bahía Seca discovered evidence of what was probably the same El Niño. Bahía Seca is a small coastal administrative center that also formed part of the Sechín Alto polity based within the Casma Valley (Pozorski and Pozorski 1998) and served the inland polity by supplying marine resources to larger inland mound-dominated sites in return for agricultural products. At Bahía Seca, wash sediments from the El Niño rainfall on the nearby ridge of coastal hills impacted the site and are clearly visible in our excavations. Chronologically, this wash seals the earlier Sechín Alto polity Initial Period occupation, providing a clear separation between it and a later Initial Period reoccupation of the site soon after the El Niño wash event. The new inhabitants were outsiders associated with the Las Haldas polity centered on the coast 20 km south of the Casma Valley (Pozorski and Pozorski 2006). Middens rich in shellfish remains formed part of both occupations, and both contained substantial C. chorus remains. The presence of C. chorus in both pre- and post-Niño occupations at Bahía Seca attests to the species' ability to rebound relatively quickly after this type of environmental challenge.
Data from the Maranga and Bahía Seca sites summarized above indicate that, while the impact of strong El Niño events on shellfish may be severe, the evidence of rebound potential described here suggests that massive total extinctions along long segments of coastline are unlikely. Intervals among ENSO events are generally measured in multiple decades, allowing time for C. chorus to reproduce and become quite large despite its relatively slow growth rate.
On a larger scale, Sandweiss et al. (2001) and Sandweiss, Maasch, and Kelly (2024) used the biogeography of mollusks to develop an El Niño history for north and north-central coastal Peru. Based on three early sites with warm-water shellfish species, they argued that the waters off northern Peru were warmer before about 3800 B.C. and that ENSO events were also absent or significantly different prior to that time. An alternative explanation for these pockets of warm-water shellfish argues that they represent colonies of thermally anomalous fauna whose larvae were carried southward within the warm currents of a periodic ENSO and settled in sun-warmed bays and estuaries at the mouths of some rivers (DeVries and Wells 1990; Pozorski and Pozorski 2003; Wells and Noller 1997). Research by DeVries and Wells (1990) and our studies at Almejas (Pozorski and Pozorski 2003) documented contemporary assemblages of cold-water mollusks in adjacent open water zones that further support the argument that the ENSO phenomenon was in place well before 3800 B.C. This is in accord with studies that describe dates for the origins of ENSO at more than 100,000 years ago and perhaps even millions of years ago (Cane 2004; Giralt et al. 2007).
Sandweiss et al. (2001) also correlate their proposed date of 3800 B.C. for the onset of fully modern ENSO with the onset of monumental mound construction associated with complex cultural development along the Peruvian coast. They assert that archaeological shellfish data reflect an increase in ENSO frequency between 1200 and 800 B.C., the time span correlated with abandonment of monumental constructions along the Peruvian coast. Focusing on the archaeological prehistory of Mesodesma donacium and C. chorus, Sandweiss et al. (2001) describe both species as absent from coastal sites north of 10oS prior to 3800 B.C. 2 , common in archaeological middens from 3800 to 1000 B.C., and virtually gone from the archaeological record by 800 B.C. when they were replaced by Donax obesulus. Using this information, Sandweiss et al. (2001) conclude that the rapid disappearance of Mesodesma donacium and C. chorus from the archaeological record probably reflects an increase in the frequency of strong ENSO events that caused the near extinction of these molluscan species while simultaneously influencing prehistoric cultural development.
It seems clear that the El Niño/ENSO phenomenon potentially impacted the survival of C. chorus because of its varied and sometimes strong effect on the coastal environment. However, the scenario proposed by Sandweiss et al. (2001) overemphasizes this impact because it so narrowly advocates for environmental determinism, leaving little room for the impact of human agency on subsistence strategies or even cultural development.
Biological Factors Affecting C. chorus
The symbiotic relationship between C. chorus and a red alga as well as the choro mussel's relatively slow growth rate are biological features that potentially make this species more susceptible to the effects of environmental and cultural factors. C. chorus is not well adapted for recruitment directly onto open rocky faces. The choro mussel has a weak byssus (López, López, and González 2008), and successful recruitment of C. chorus is facilitated by interaction with the filamentous red alga Asterfilopsis furcellata [formerly known as Gymnogongrus furcellatus] (Carranza et al. 2009; Moreno 1995; Neely, Brickle, and Laptikhovsky 2010). Asterfilopsis furcellata forms a critical refuge for juvenile C. chorus individuals, protecting them from the predatory whelk, Nucella crassilabrum, until they reach a length of 3 cm which places them outside the preferred size range of the whelk's prey (Moreno 1995). As young C. chorus individuals attain this size, their combined weight bends the fronds of the alga into contact with the substrate allowing them to attach there and form beds. This phase of the life cycle of C. chorus potentially makes the choro mussel more susceptible to environmental changes because the alga Asterfilopsis furcellata is initially adversely affected by warmer El Niño waters (Vinueza et al. 2006). Warmer El Niño waters can also indirectly impact C. chorus because known predators of the mussel, such as Thaisella chocolata, greatly increase in frequency as they prey on species weakened or killed by rising water temperatures (Diaz and Ortleib 1992; Gómez-Cornejo 1986).
The choro mussel gets one of its common names (giant mussel) from its potential to attain a very large size. Maximum lengths for this species have been reported as 186 mm (Reid and Osorio 2000, 131–132), 200 mm (Guzmán, Saá and Ortleib 1998, 62–63), and even 300 mm (Suchanek 1985). It takes a very long time to attain such a large size. Choro mussels initially grow rapidly, reaching a size of 60–70 mm during the first 2 years (Nurambuena and Solis 1978). Additional studies indicate C. chorus may reach sexual maturity as early as one year of age (Neely, Brickle, and Laptikhovsky 2010) and at a size as small as about 40 mm (Ruiz-Velásquez et al. 2017). However, it is only once the choro mussel reaches 60–70 mm that its growth slows considerably, a rate decrease likely correlated with the onset of fully active reproduction and the greater energy expenditure it requires (Gobierno de Chile 2010, 39). Measured individuals reached a size of ca 102 mm during the next three years and stabilized at 12 years and 144 mm (Gobierno de Chile 2010, 38; López, López, and González 2008; Nurambuena and Solis 1978). C. chorus also has a slow overall growth rate relative to the other large South American mussels. Both Aulacomya atra and especially Mytilus chilensis are faster feeders and more able to digest and absorb more food in a given time than C. chorus (Ibarrola et al. 2012). C. chorus' slower relative growth rate may be related to these key metabolic factors
Human Exploitation of C. chorus
Along most of the coast of Peru, C. chorus virtually disappeared from archaeological sites within the coastal valleys at the end of the Initial Period, ca 1000 B.C. (Pozorski 1976a; Pozorski and Pozorski 1987a; Sandweiss et al. 2001). If human exploitation played a role in this disappearance, we would expect the archaeological faunal record to reflect a gradual decrease in the number and/or size of C. chorus valves through time. A change in subsistence strategies might also be evident, reflecting efforts to compensate for decreasing marine protein by exploiting alternative sources. Broader associated cultural changes might accompany changes in subsistence, especially as economic relationships among sites change.
To better define this phenomenon of near extinction and to explore its magnitude and extent, we looked at archaeological sites of various time periods with reported subsistence data along the coastal zone of Peru and northern Chile (Figure 3). Table 1 shows the results of this investigation. The time-frame designations used in the table are generally based on data from Rivera (2008) and Quilter (2022). Precise quantitative subsistence data were often not available; therefore, we used a relative scale of “abundant” through “absent” expressed by the use of shading. These data confirm the time frame for the near extinction of C. chorus at the end of the Initial Period and reveal that the geographical extent of this phenomenon appears limited to valleys north of the Chincha area of southern Peru. Looking farther south in Peru and northern Chile, we start to see significant amounts C. chorus remains in both early and later sites, indicating that C. chorus persisted as a significant food source there well after the end of the Initial Period. The apparent chronological connection between the disappearance of C. chorus and a significant and recognized cultural boundary (end of the Initial Period) suggests that human exploitation of the mollusk may have been a key factor related to the mussel ' s decline while the existence of a southern boundary for this phenomenon indicates environmental factors are also important.
Relative Quantification of Choromytilus chorus Remains From Early Preceramic Through Early Historic Archaeological Sites Along the Coastal Zone of Peru and Northern Chile.

Numerous efforts have been made to investigate and demonstrate how ancient populations may have impacted coastal biodiversity by overharvesting certain molluscan species. Jerardino et al. (1992) looked at locos (Concholepas concholepas) and limpets (Fissurella limbata) at coastal sites in central Chile; and Jerardino (1997) studied mussels (Choromytilus meridionalis) and two species of limpets (Cymbula granatina and Scutellastra granularis) as well as the rock lobster (Jasus lalandii) from western South Africa. Milner, Barrett, and Welsh (2007) investigated potential overharvesting of limpets (Patella sp.) used as codfish bait by Vikings in the Orkney Islands of Great Britain. In all cases, a decrease in shellfish size, documented by measurements of whole shells or near-whole shells, comprised the key evidence for overexploitation.
We set out to investigate human impact on C. chorus’ northern range in detail by looking at archaeological data from the north and north-central coastal valleys of Moche and Casma where we conducted subsistence studies that yielded quantitative data on shellfish use and consumption, including C. chorus. Within the Moche Valley (Figure 4), Shelia Pozorski carried out stratigraphic excavations of midden deposits at nine sites ranging in date from Late Perceramic through Inca times (2500 B.C.‒A.D. 1500) (Pozorski 1976a). Within the Casma Valley (Figure 5), we conducted similar subsistence studies at 10 sites spanning the Middle Preceramic through the Early Horizon (7000 B.C.‒A.D. 1).
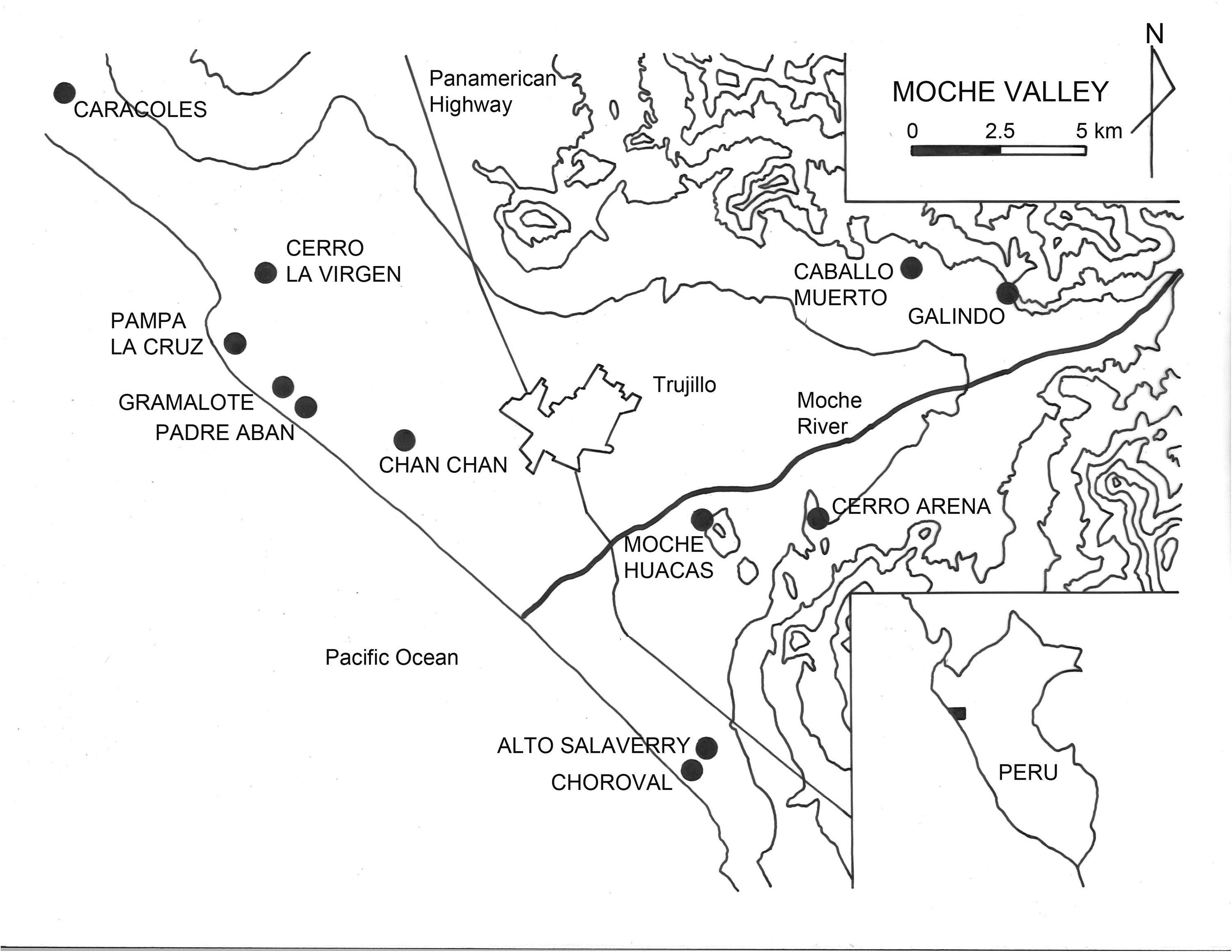
Map of Moche Valley indicating archaeological sites mentioned in text and Tables 1 and 2.
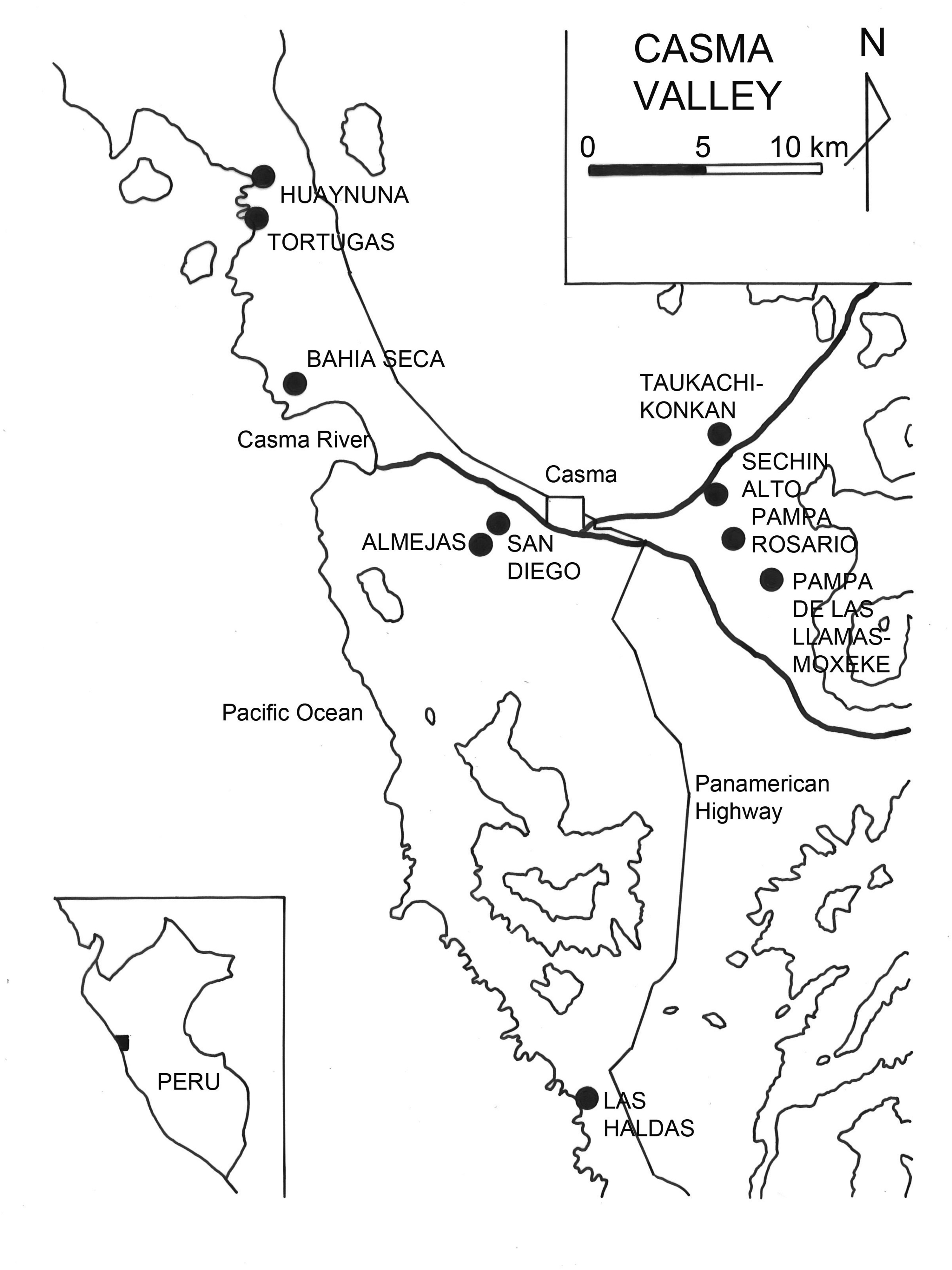
Map of Casma Valley indicating archaeological sites mentioned in text and Tables 1 and 3.
All the samples examined for this study came from controlled stratigraphic excavations of midden or refuse contexts. Molluscan remains from these excavations were washed and identified to species or genus as feasible. Counts were made of whole individuals or whole valves as well as unduplicated parts of fragmentary specimens in order to determine the minimum number of individuals
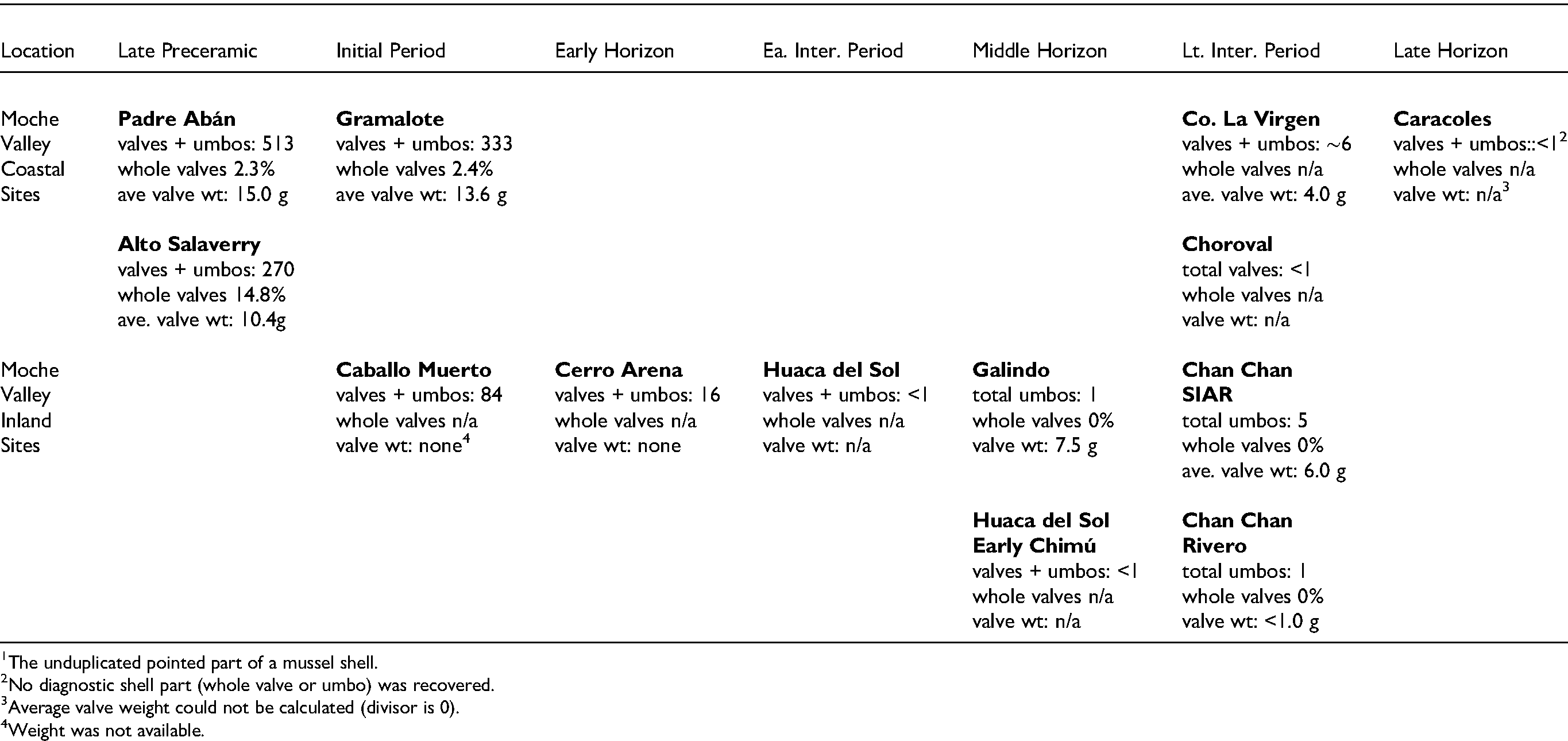
As we focused on C. chorus data from these subsistence studies, we realized that very few whole valves had been recovered, especially valves of larger specimens, because C. chorus was routinely broken open to extract the meat. Percentages of whole valves ranged from 14.8% at the Moche Valley preceramic site of Alto Salaverry to 0% at numerous sites with small sample sizes, with most values between 0.1% and 7.6% (Tables 2 and 3). This evidence of such severe breakage of predominately larger valves made it clear that changes in valve size could not be reliably assessed using linear measurements. As an alternative we assessed changes in shell size using total weights of shell samples. We counted the total number of whole valves plus umbos (the unduplicated pointed parts of a mussel shell) at each site, and we calculated average shell weights (total weight of shell recovered ÷ total number of valves and umbos) for each site or cultural component. Assessing shell size by weight instead of MNI or whole-shell length enabled us to account for the many fragmented specimens in our calculations. The results of this assessment are presented in Tables 2 and 3. Data within the tables reveal that sample sizes are variable and often very small, reflecting the scarcity of C. chorus in later time periods.
Total Number of Valves and Umbos,1 Percentages of Whole Valves, and Average Valve Weight Values for Choromytilus chorus from Moche Valley Archaeological Sites.
Total Number of Valves and Umbos,1 Percentages of Whole Valves, and Average Valve Weight Values for Choromytilus chorus from Casma Valley Archaeological Sites.

Recognizing that other studies of human impact on shellfish species typically use shell length to quantify size (Jerardino 1997; Jerardino et al. 1992; Milner, Barrett, and Welsh 2007), and to assess the relationship between shell weight and shell size, we also made measurements on whole shells. For example, linear measurements on some of the larger whole specimens we recovered archaeologically indicate that individual valves with a weight of 30 g are typically about 130 mm in length. Using our sample of 11 whole excavated valves that ranged from 57–74 mm in length we calculated an average weight of 5.4 g as the approximate weight of an archaeological specimen that had reached the length of 60–70 mm indicative of full sexual maturity.
Detailed data from the Moche Valley come from three sites that predate the critical Initial Period/Early Horizon boundary when C. chorus virtually disappeared from the archaeological record and six sites that postdate the boundary (Figure 4) (Table 2). Choro mussel data from the Moche Valley suggest that a decrease in the size of C. chorus valves may have occurred along with the species' decrease in frequency at the end of the Initial Period (Table 2), and this possible size decrease seems to begin even before the end of the Initial Period. At the Preceramic site of Padre Aban, the average weight of a single valve of C. chorus is 15.0 g. This is one of the largest average sizes recorded for all the sites studied, and the valves recovered were described during analysis as mostly large and riddled with parasites (Pozorski 1976a, 74–75). These observations suggest that the inhabitants of Padre Aban were likely among the first people to exploit the local C. chorus population in the Huanchaco Bay area and that they were selectively harvesting large individuals. Alto Salaverry, the other Preceramic Moche Valley site studied, yielded smaller C. chorus shells with an average weight of 10.4 g. The smaller size of these valves and the absence of effects of parasites were noted during analysis (Pozorski 1976a, 82–83). These data likely reflect more intense and/or longer exploitation by the population of the larger site of Alto Salaverry which put more pressure on C. chorus populations accessible from the site. Excavations at the Initial Period site of Gramalote yielded C. chorus valves with an average weight of 13.6 g, a value closer to the average for Preceramic Padre Aban. The sites are very near each other and their inhabitants likely exploited the same local beds of C. chorus within the Huanchaco Bay area. Individual specimens from Gramalote were also described as mostly large individuals, with some valves affected by parasites (Pozorski 1976a, 96–97). Taken as a whole, these data may indicate that, by Initial Period times, the local C. chorus population was showing evidence of exploitation pressure because the average size was slightly smaller and fewer very old individuals with parasites were present.
Subsequent excavation by Prieto at Gramalote also yielded abundant remains of C. chorus (Prieto 2011). In their report on the Gramalote mollusks, Gorriti and Martín (2011) assessed individual shellfish along a continuum of size ranges from very large (150 mm/+) to very small (< 79 mm). Results showed that most of the mussels consumed were medium to small, with many more also being very small. Despite the difference in measurement methods used (weight vs. length), Prieto' s data seem to support our determination that choro mussels consumed at Gramalote were smaller than choro mussels consumed at the earlier nearby site of Padre Aban. Prieto (2014) also hypothesized that local marine resources consumed at Gramalote, including mollusks, diminished through time, potentially causing social tension and ultimately a change in subsistence focus from sandy to rocky beaches.
During the Initial Period, Moche Valley populations located well inland also depended on marine resources for animal protein. Within the valley, the establishment of the sizeable site complex of Caballo Muerto reflects an emphasis on irrigation agriculture by its location near arable land (Pozorski 1976b) (Figure 4). However, these sites initially depended primarily on marine resources for protein, confirming an inland-coastal link. Prominent among the Caballo Muerto shellfish is C. chorus, represented by 84 valves (Table 2). Based on coincident species inventories between Gramalote and this early Caballo Muerto occupation, we suggested that the two sites formed an early subsistence exchange system that served to distribute marine resources to inland sites and agricultural resources to coastal sites during the Initial Period (Pozorski and Pozorski 1979). By the end of the Initial Period, the Moche Valley economy was about to transition from a coastal orientation to a predominantly inland subsistence focus dominated by agricultural products and domesticated animals, and this transition was nearly complete by Early Horizon times. At the Early Horizon Salinar site of Cerro Arena, Curtiss Brennan describes a faunal inventory dominated by domesticated camelids and guinea pigs (Brennan 1978, 339–341). He notes that “marine resources played an unexpectedly reduced role in the Cerro Arena diet, and their contribution was restricted almost entirely to shellfish.” Within the molluscan inventory of Cerro Arena, C. chorus is represented by 16 whole shells or valves (Brennan 1978, Table 6) (Table 2).
As the very small sample sizes for the later sites in Table 2 make very clear, C. chorus had virtually disappeared from the archaeological record of the Moche Valley by the Early Intermediate Period (A.D. 1–650). Neither coastal nor inland sites yielded significant amounts of C. chorus, an indication that site location is not a factor. Average shell weights are also much smaller. Values for the three later sites where average valve weight could be calculated show shell-weight values considerably smaller than the values for earlier Preceramic and Initial Period sites. These values of 7.5, 6.0, and 4.0 g are also closer to or even smaller than the estimated 5.4 g average weight that correlates with full sexual maturity. We believe that this considerable decrease in both the frequency and size of C. chorus is due largely to human overexploitation of this valued food species. The few C. chorus individuals that continued to be harvested at later sites probably reflect occasional choro mussel larvae carried north as plankton within the Peruvian Current that settled in hospitable environments and grew large enough to be noticed by local shellfish collectors but rarely grew large enough to reproduce and form a colony.
Detailed subsistence data from the Casma Valley are restricted chronologically to the Middle Preceramic through the Early Horizon (7000 B.C.‒AD 1) (Table 3), and this enabled us to focus on the critical time span when the most severe decline in C. chorus populations occurred. Table 3 also reveals that the story of C. chorus size changes within the Casma Valley is a complex one, reflecting the interplay of various cultural factors. We have Preceramic data from five coastal sites both near and within the Casma Valley. Huaynuná and Tortugas lie north of the valley mouth; Almejas and Bahía Seca are near the Casma River mouth; and Las Haldas is well south of the Casma Valley edge (Figure 5). The Initial Period sample comes from later occupations at the coastal sites of Huaynuná, Tortugas, Bahía Seca, and Las Haldas as well as from the inland sites of Sechín Alto, Taukachi-Konkán, and Pampa de las Llamas-Moxeke. The Early Horizon is represented by very small terminal occupations at the coastal sites of Huaynuná and Las Haldas, by the coastal site of San Diego, and by the inland site of Pampa Rosario.
Almejas, the earliest Casma Valley site, dates to the Middle Preceramic and consists of a small shell midden. When Almejas was occupied, the inhabitants’ subsistence focus was a rich estuary at the river mouth that teemed with young fish and warm-water mollusks from the north that had colonized the sun-warmed habitat during an El Niño episode (Pozorski and Pozorski 2003). Cold-water species from sandy bay and open-ocean habitats were also occasionally exploited, including C. chorus. The large average weight of 13.9 g for the few examples of C. chorus recovered suggests harvesting was selective, reflecting a focus on relatively large individuals (Table 3).
Data from the four subsequent occupations reveal that exploitation of C. chorus had intensified by Late Preceramic times. Huaynuná and Las Haldas are located near sandy bays, likely prime habitats for C. chorus (Pozorski and Pozorski 1990b). The relatively large average shell-valve weight of 11.9 g for Preceramic Huaynuná and 11.2 g for Preceramic Las Haldas (Table 3) may indicate selective harvest of larger individuals from populations that had not previously been exploited. Analysis notes for Las Haldas describe some hinges as coming from mature individuals, an observation in accord with the large shell-weight value. Inhabitants of Preceramic Tortugas and Bahía Seca focused more exclusively on C. chorus, and this is reflected in the larger valve totals as well as smaller average shell sizes of 4.3 g and 6.9 g respectively (Table 3) which may indicate indiscriminate harvesting whereby whole colonies of C. chorus were scooped up without regard for size. This practice resulted in pressure from overexploitation, collecting of large numbers of individuals that had not reached reproductive size of 5.4 g.
Data from Casma Valley Initial Period occupations suggest a more complex set of cultural variables impacted average valve sizes (Table 3). The average valve weight value of 27.3 g for Initial Period Huaynuná is surprising. Several explanations are possible. The sample size is very small (total MNI = 10) and large individuals may have been selectively harvested. The Initial Period occupation of Huaynuná is also relatively small, possibly resulting in lower selective pressure during this time span. At Tortugas, there was less emphasis on C. chorus during Initial Period times, and the valve-weight value of 13.2 g is relatively high. Both selective harvesting of larger individuals and lower selective pressure may also have impacted this value.
In contrast to the higher shell-weight values for Huaynuná and Tortugas, we see a marked decrease in valve-weight values for Bahía Seca and Las Haldas during Initial Period times (Table 3). The Initial Period archaeological sequence at Bahía Seca is complicated because of evidence that people with Las Haldas-type ceramics reoccupied the site near the end of the Initial Period. For most of the Initial Period, the Sechín Alto Polity within the Casma Valley centered at the inland site of Sechín Alto coexisted with a polity centered at the coastal site of Las Haldas, 20 km to the south; however, there is no evidence of interaction between the two groups until late in the Initial Period (Pozorski and Pozorski 2006). For most of its existence, coastal Bahía Seca was vital to the Sechín Alto polity as part of a subsistence exchange system that facilitated movement of coastal marine resources to inland sites and agricultural products to coastal sites (Pozorski and Pozorski 2012b). 3 We argued that the coastal component of this exchange system was administered from Bahía Seca based on an administrative structure at the site, and the high volume of shellfish collected there indicates most of the marine resources that moved inland probably came from the vicinity of Bahía Seca. The high demand for marine resources to feed both coastal and inland populations probably resulted in indiscriminate harvesting and extreme pressure from overexploitation, intensifying a pattern established during Preceramic times. The shell-weight value of 2.7 g for Bahía Seca reflects such severe overexploitation because it is the lowest recorded for the Initial Period (Table 3). C. chorus exploitation from the polity center at Las Haldas that occurred during most of the Initial Period is reflected by the shell-weight value of 5.4 g (Table 3). This low value also indicates both indiscriminate harvesting and exploitation pressure on local mussel populations because half of the individuals taken were below average reproductive size. Our analysis notes also describe the collection as consisting of mostly small, immature individuals.
Late in the Initial Period (about 1350 B.C.), substantial construction was carried out at the coastal polity center of Las Haldas; and the polity aggressively expanded as people affiliated with the coastal center reoccupied several sites within the Casma Valley, including the inland sites of Sechín Alto and Taukachi-Konkán as well as Bahía Seca on the coast (Pozorski and Pozorski 2006). C. chorus data from the Las Haldas late Initial Period occupation of Bahía Seca is remarkable because the total of 6,670 valves reflects an intense focus on this species while the shell-weight value of 4.2 g is one of the smallest recorded. Our discussion of environmental factors documented an El Niño episode immediately prior to the Las Haldas late Initial Period reoccupation of Bahía Seca, and the relatively small size of the C. chorus within debris from this occupation may reflect some impact from this El Niño. The large number of individuals harvested combined with the small valve size also indicate that harvest was both intensive and indiscriminate. Few individuals reached reproductive size, a factor that argues strongly for severe overexploitation.
Sechín Alto, Taukachi-Konkán, and Pampa de las Llamas-Moxeke are all inland sites that formed part of the Sechín Alto polity centered at Sechín Alto site (Pozorski and Pozorski 2012b). The average shell-weight values for these sites of 10.7, 8.9, and 8.4 g respectively are relatively large (Table 3). In the context of the proposed subsistence exchange system, these sites would have provided agricultural products to coastal sites in return for marine resources. Bahía Seca likely administered the coastal side of this exchange and also probably provided most of the shellfish for the exchange. As part of this system, larger specimens of C. chorus were apparently selected for shipment inland, thereby accounting for the larger shell-weight values from inland sites. Sechín Alto site, the polity capital, may have received the largest mussels because it had more prestige, power, and influence within the polity. Removal of larger specimens from the total Bahía Seca harvest for distribution to inland sites would have impacted average valve-size values at the coastal site, and this may account for the unusually small average valve weight of 2.7 g that we initially calculated for Initial Period Bahía Seca (Table 3). We subsequently calculated a combined value for all three inland sites plus Bahía Seca, and the average valve weight for all four sites is 4.6 g. This value is more consistent with the values of 5.4 and 4.2 g for Initial Period and late Initial Period Las Haldas; however, it still reflects a harvested population of mostly sexually immature individuals.
By Early Horizon times, C. chorus had virtually disappeared from the archaeological record at both coastal and inland sites in the Casma Valley area (Table 3). Evidence of the mussel is scarce and in two of the four sites there is no diagnostic unit (whole valve or hinge) to count or to calculate an average valve-weight value. Where average valve-weight values could be calculated, they are small, again reflecting overexploitation and rarity of C. chorus by this time. As was the case for the Moche Valley, the few C. chorus mussels taken during the Early Horizon probably represent isolated individuals that traveled north as plankton and settled in the Casma Valley area.
The disappearance of C. chorus from the archaeological record was permanent for much of the north and north-central Peruvian coast. It is not found there in modern times. While working at Huaca Negra in the Viru Valley in the 1940s, Strong and Evans (1952, 23) questioned local inhabitants about abundant C. chorus remains found in their excavations; and everyone asserted that this species does not exist in the area today. When we questioned fishermen and shellfish collectors from coastal towns in the Moche Valley during the 1970s, none of them had ever seen live C. chorus.
López, López, and González (2008) provide an historic overview of the ramifications of overexploitation of C. chorus in Chile within the current range of the species. According to Reyes (1986 cited in López, López, and González 2008) C. chorus was one of the two “most exploited species at the beginning of the twentieth century.” This prompted some regions to take action as early as 1907 by imposing a seasonal moratorium on the harvest of C. chorus during breeding seasons (Gobierno de Chile 2010, 2). Nevertheless, shellfish and fish capture in 1926 totaled 20,000 tons, and 3,688 tons of this total consisted of C. chorus. Harvest of C. chorus peaked again from 1938–1944 when landed tonnage for this species averaged 1,700 tons annually (11,900 tons over 7 years). From 1945–1986 average annual tonnage harvested dropped almost 90% to only 190 tons per year, a decrease attributed to overexploitation that also resulted in a harvesting ban (Guzmán, Saá, and Ortleib 1998; Hancock 1969 cited in López, López, and González 2008; Lozada and Cruz 1979 cited in López, López, and González 2008) and establishment of a minimum legal size limit of 105 mm in 1986 (Ávila et al. 1994 cited in Carranza et al. 2009). The harvesting ban lasted 10 years, ending in 1992 (Osorio 2002). Culturing of C. chorus was implemented in 1965; and over 40% of the harvest since that time represents cultivated individuals (López, López, and González 2008). In 1981, a Genetic Reserve (Reserva Genética Putemún) was created in southern Chile to conserve genetic stock for C. chorus and Mytilus chilensis and insure larvae production (Carranza et al. 2009; Gobierno de Chile 2010).
Discussion
C. chorus virtually disappeared from over 550 km of coastal Peru around 1000 B.C. By archaeological standards, the event was sudden; and it coincided with a cultural boundary at the end of the Initial Period (Tables 1–3). Such a coincidence is a strong indication that human factors are significant, making it informative to examine the cultural context that enabled such a phenomenon to occur. The large-scale extinction documented here was possible because the Andean cultural area, centered in north-central coastal Peru, was the locus of highly cohesive cultural development (Pozorski and Pozorski 1987b, 42–43). Preceramic settlements located on the coast reflect the early importance of marine resources as the major protein source, and this reliance on fish and shellfish continued through the end of the Initial Period. The Initial Period along most of coastal Peru saw a marked change in settlement patterns whereby large sites dominated by monumental platform mounds were established well inland near irrigated lands while sizable coastal settlements were maintained to supply essential marine resources (Burger 1992, 57–58; Pozorski and Pozorski 1979). The reliance on marine resources established during Preceramic times continued through the end of the Initial Period, however, the terminal Initial Period-Early Horizon transition saw a sudden decreased dependence on marine resources, especially shellfish, as domesticated mammal species already consumed in the highlands (Stahl 2008) increased in importance nearer the coast. In keeping with this, settlement patterns shifted again as many of the large Initial Period coastal sites ceased to be occupied. Later sites of the Early Intermediate Period and subsequent time periods relied even more heavily on domesticated mammals, especially camelids.
This study has focused on the fate of C. chorus as we used data from coastal Peru and northern Chile and especially the Moche and Casma Valleys to demonstrate how the choro mussel decreased significantly in size from Preceramic through Initial Period times until it faced near extinction at the end of the Initial Period. This gradual size decrease shows the sequence of change-how the initial selective harvest of large old specimens evolved into overexploitation as harvesting became indiscriminate and increasingly intense, finally culminating in the capture of mostly immature individuals, thereby crippling the species' ability to reproduce itself. C. chorus was not the only overexploited shellfish species. Other large, high-yield bivalves, including Eurhomalea rufa, Semele corrugata, and Leukoma thaca became very scarce or absent in the archaeological record at the end of the Initial Period. These data suggest that overharvesting of prime food shellfish generally intensified through the Initial Period, rendering the large early coastal sites less and less essential and ultimately resulting in their abandonment. The changing economics predicated by the decline of marine resources may have helped push post-Initial Period cultures toward a reliance on domesticated mammals for protein.
Conclusions
No single factor seems totally responsible for the synchronous near extinction of C. chorus along much of the Peruvian coast about 1000 B.C. Nevertheless, human overexploitation stands out as the most deleterious factor examined here. C. chorus was a highly prized shellfish valued as food, as a raw material for tools, and intrinsically because of its size, shape, and color. By Initial Period times, human populations had increased in size as irrigation agriculture expanded, putting serious pressure on protein sources. As a result, exploitation of C. chorus became more intense and indiscriminate as whole beds of the choro mussel were likely harvested. Toward the end of the Initial Period, once most of the individuals harvested were below reproductive age and size, it was only a short step to virtual extinction.
While human overexploitation appears to have been the major causal factor in the virtual disappearance of C. chorus, biological and environmental factors were also significant, making C. chorus more vulnerable to overexploitation in several ways. Based on data for pristine or underharvested mussel colonies, the optimal habitat for C. chorus is shallow waters where they form large and dense concentrations of individuals. This accessibility would have made the species more susceptible to harvest and ultimately to overexploitation. The choro mussel's slow and uneven growth rate may have impacted its ability to compete with other species and rebound quickly while also resulting in individuals that were large enough to eat, but not sexually mature.
Environmental changes related to ENSO events had both a direct and indirect impact on the survivability of C. chorus. The mussel was impacted directly because the warmer water temperature causes sizeable kills of most mollusk species. However, C. chorus does not seem to be significantly more affected than other associated bivalves, and archaeological data from the Rimac and Casma Valleys attest to the choro mussel's ability to rebound after an ENSO event. C. chorus’ survival is also indirectly affected by ENSO events because warmer water initially adversely impacts its red alga host and predators of the mussel such as Thaisella chocolata thrive on carrion and individuals weakened by the water temperature increase.
Although El Niño events were probably significant causal factors in the near extinction of the choro mussel, data examined in this study reveal that C. chorus shows considerable resiliency with respect to El Niño-related environmental hazards. Prior to human occupation of the Peruvian coast, C. chorus colonies were present at least as far north as the Lambayeque valley area. Once humans settled along the coastline, there is archaeological evidence of substantial harvests of the choro mussels, and these data also document a gradual size decrease in C. chorus valves through time indicative of overexploitation. This suggests that human exploitation also posed a significant threat to C. chorus because if El Niño events were the only major threat to coastal mollusks, we would expect more of a cyclical pattern reflecting the mussels’ ability to rebound and reach a large size during the decades separating each ENSO occurrence. Despite these threats, the same archaeological data also reveal that this mussel survived for hundreds of years along most of the Peruvian coast in the face of both human exploitation and periodic El Niño perturbations.
We suggest that changes in the distribution of C. chorus through time can best be explained primarily in terms of the interplay between two key factors: human exploitation and El Niño-related periodic environmental changes. It seems most likely that overexploitation was initially the stronger force moving C. chorus toward extinction, especially as the breeding population was increasingly affected by the removal of immature individuals. The resultant extinction event was able to happen simultaneously along much of the Peruvian coast because this nuclear Andean area shared a similar trajectory of cultural development accompanied by a parallel increase in population that put more and more pressure on marine resources, the major protein source at the time. This relatively uniform and synchronous cultural development spanned the valleys from Lambayeque in the north to around Chilca in the south. Consequently, at about 1000 B.C., C. chorus was suddenly gone from more than 550 km of coastline along northern and central Peru, making recovery and rebound of the species virtually impossible. After this time, human populations increased even more and domesticated animals became the main protein source; however individual shellfish collectors continued to harvest the few individual mussels that managed to recruit and grow–probably collecting them before they reached reproductive maturity.
As overexploitation progressed, the impacts of El Niño-related effects came to be felt more severely by C. chorus colonies already depleted and reproductively weakened by human overexploitation. Rebound and recovery of C. chorus populations in the aftermath of El Niño events are only possible if significant numbers of adult individuals are available and survive the higher water temperatures and/or larvae travel north in the returning colder Peruvian current and manage to reach maturity in sufficient numbers to create a breeding population. Such reestablishment by larvae carried north as plankton is typically a gradual multistage progression that involves colonization of nearby suitable habitats where enough larvae survive and mature to form a breeding population from which their larvae can be carried to habitats even farther north and so on as the process repeats itself.
In reality, the process of northward colonization is more haphazard due to the same human and ecological factors that caused the initial extinction of C. chorus in much of Peru and pushed its range much farther south. When C. chorus became extinct or virtually extinct south to around the Chincha Valley in southern Peru about 1000 B.C., this zone became the approximate northern limit of its range at that time. Historic twentieth-century accounts that document the changing northern boundary of the choro mussel probably represent records of rebound and recovery in process as colonies became established and succeeded or failed. Their success continues to be hampered by the two key factors identified here: human overexploitation and effects of ENSO events. Farther south, however, in southern Peru and northern Chile, where El Niño events are rare to absent and prehistoric populations were not large, the archaeological data document sites with substantial C. chorus remains dating both before and after 1000 B.C. (Table 1) that were not severely impacted by climatic or human factors.
Footnotes
Acknowledgments
Permission to excavate at the many sites with C. chorus remains was granted by the Instituto Nacional de Cultura and the Ministry of Culture of Peru. We also want to thank all the U.S. and Peruvian students and workers involved in the excavation and analysis that provided data for this study.
Funding
The authors disclosed receipt of the following financial support for archaeological excavations and analysis. Funding for excavation and analysis came from National Science Foundation grants BNS-823452, BNS-8406598, SBR-980-6833, 0455333, and 0924535 as well as the National Geographic Society, O’Neil and Netting Research Grants from Carnegie Museum of Natural History, Wenner-Gren Foundation for Anthropological Research, Heinz Charitable Trust and Heinz Family Foundation, J. M. Kaplan Fund, Brennan Foundation, the Institute of Latin American Studies at the University of Texas-Austin, the University of Texas-Rio Grande Valley, and a generous anonymous donor.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.