
Abstract
Myelination is an important process in the central nervous system (CNS). Oligodendrocytes (OLs) extend multiple layers to densely sheath on axons, composing the myelin to achieve efficient electrical signal conduction. The myelination during developmental stage maintains a balanced state. However, numerous CNS diseases including neurodegenerative and cerebrovascular diseases cause demyelination and disrupt the homeostasis, resulting in inflammation and white matter deficits. Effective clearance of myelin debris is needed in the region of demyelination, which is a key step for remyelination and tissue regeneration. Microglia and astrocytes are the major resident phagocytic cells in the brain, which may play different or collaborative roles in myelination. Microglia and astrocytes participate in developmental myelination through engulfing excessive unneeded myelin. They are also involved in the clearance of degenerated myelin debris for accelerating remyelination, or engulfing healthy myelin sheath for inhibiting remyelination. This review focuses on the roles of microglia and astrocytes in phagocytosing myelin in the developmental brain and diseased brain. In addition, the interaction between microglia and astrocytes to mediate myelin engulfment is also summarized.
Introduction
Myelin in the central nervous system (CNS) is composed of oligodendrocytes (OLs), and myelin is enriched in lipids, and tightly encase axons. 1 The special structure provides the axons with insulating layers, enabling the myelination of axons to achieve rapid and efficient conduction of electrical signals.2–4 Apart from early differentiated OLs derived from dorsal- or ventral-born oligodendrocyte progenitor cells (OPCs), new OLs are continuously generated from the proliferation, migration and differentiation of OPCs that persists during adulthood.5,6 Myelination is a relatively stable process with long lifetime proteins involved, 7 but also a dynamic process with slow turnover. 8
The balance of demyelination and remyelination in the developmental brain and diseased brain is crucial for CNS homeostasis, and it requires rapid clearance of extra myelin fragments. 9 Excessive engulfment of myelin triggers inflammation and severe demyelination, 10 while insufficient phagocytosis causes myelin debris accumulation, and results in impaired remyelination. 11 Microglia, the major resident phagocytes in the brain, play a major role in myelin engulfment. In the healthy brain, microglia are inactive with a small cell body and highly ramified branching structure. 12 Upon demyelinated injury, microglia transform into an active phagocytic phenotype, migrate, and accumulate in the injured areas. Activated microglia play dual roles and have a biphasic influence on the brain. They produce both anti- and pro-inflammatory factors, and are capable of removing both myelin debris and healthy myelin sheath. 13 Microglia are widely regarded as the main professional phagocytes in the CNS, and some researchers claim astrocytes have limited phagocytosis abilities.14–16 However, astrocytic phagocytosis has been recently found as a compensatory mechanism if microglia is dysfunctional.17,18 Astrocytes are now emerging as important contributors to white matter integrity, which may exert similar functions in myelin engulfment as microglia. Microglia and astrocytes are activated in various neurological diseases,19,20 including neurodegenerative diseases,21,22 cerebrovascular diseases,23,24 spinal cord injury (SCI), traumatic brain injury (TBI), aging, etc.. 25 Activated glial cells exert bilateral effects on myelination.
In this article, we summarized the roles of microglia and astrocytes in myelin engulfment in the developmental brain and brain after injury, including the process of phagocytosis, and the signals involved in microglia and astrocytes mediated myelin phagocytosis. In addition, the microglia-astrocytes crosstalk in myelin phagocytosis is also reviewed.
Myelination and glial cell-mediated phagocytosis in developmental brain
Healthy myelination is critical to brain development. OLs myelinated axons to achieve fast and efficient nerve signaling conduction,26,27 which depends on the proliferation, migration, differentiation and maturation of OPCs to generate lipidic sheath. The OPCs occur in several stages in mice before differentiation with spatial differences.1,28 The first stage appears at embryonic day (E) 12.5 in the ventral domain of the ventricular wall in the developing brain and spinal cord. The second one arises more dorsally at E15.5 in both brain and spinal cord. The last one emerges specifically in the perinatal cortex around birth. The mechanism of myelination is still controversial. Currently, there are three main opinions about myelination process during development.29,30 The first hypothesis supports that more myelinated white matter at birth will develop faster postnatally and finish myelination earlier. On the opposite, the second hypothesis predicts that less myelinated white matter at birth develops faster postnatally and finishes earlier. The last is the spatial-gradient hypothesis, which claims that postnatal myelination develops in a spatially organized manner. With the rapid development of neuroimaging techniques, magnetic resonance imaging (MRI) results support the second and third hypotheses, suggesting that the process of myelination in early infancy is related to both initial myelin content and spatial gradients. 29
Myelination is a complex and relatively dynamic process. 3 Firstly, OPCs migrate away from the neuroepithelium of the ventricular/subventricular zone of the brain into the developing white matter, in which they proliferate and form an evenly spaced network. Although some OPCs remain in a precursor state, others will differentiate into myelin-forming OLs. After the myelin compaction and node formation, the myelination is completed. With the maturation of OLs, they begin to express a large number of myelin-associated genes, including myelin basic protein (MBP), myelin proteolipid protein (PLP), myelin‐associated glycoprotein (MAG) and myelin oligodendrocyte glycoprotein (MOG) to finish the assembly of myelin sheaths enwrapping axons. 31 Besides, myelin has a very high lipid content, accounting for 70–75% of its dry weight. The integrity of myelin lipids and proteins is also essential to healthy myelination. In addition to OLs, the balance of a healthy glial cell population is crucial to myelin development, homeostasis, and plasticity.4,32 A certain subset of microglia populating axon tracts promotes myelination during brain development. Astrocytes influence myelin thickness and nodal gap length via thrombin protease secretion. The gap junctions between astrocytes and OLs contribute to the metabolic support of both glial cells, and myelination. Neuronal activity regulates myelin sheath dynamics and axon selection for myelination. Neuronal activity also promotes OPC proliferation, differentiation and myelination, which is a process that continues to shape myelin during adulthood. Several neurotrophic factors and growth factors, including neurotrophin-3 (NT-3), platelet-derived growth factor (PDGF), and ciliary neurotrophic factor (CNTF), are critical for OL proliferation and differentiation, and brain derived neurotrophic factor (BDNF) influences OL’s dynamics and OPC proliferation. The constructed myelin connects different brain regions, and hemostasis is crucial to cognition and learning. During the formation and development of myelination, excess pre-myelinating oligodendrocytes are generated and undergo apoptosis. Efficient clearance of dying oligodendrocytes by microglia and astrocytes is critical for white matter development. 33
Phagocytosis is commonly categorized into three steps, including “find-me”, “eat-me”, and “digest-me”, and each step is regulated by specific molecules.13,33 First, damaged or apoptotic cells released chemoattractant molecules such as extracellular nucleotides adenosine triphosphate (ATP) and uridine triphosphate (UTP), fractalkine, etc. to attract phagocytic cells and initiate the “find-me” process. Phagocytic cells express corresponding receptors to recognize the ligands in the membrane of the targeted cells (“eat-me” signals). In addition to “eat me” signals, cells also express “don’t eat me” molecules to avoid being engulfed by phagocytic cells. Finally, engulfed materials are delivered to the phagosome, and ultimately degraded in the phagolysosome by multiple enzymes, which marks the end of “digest-me” process (Figure 1).
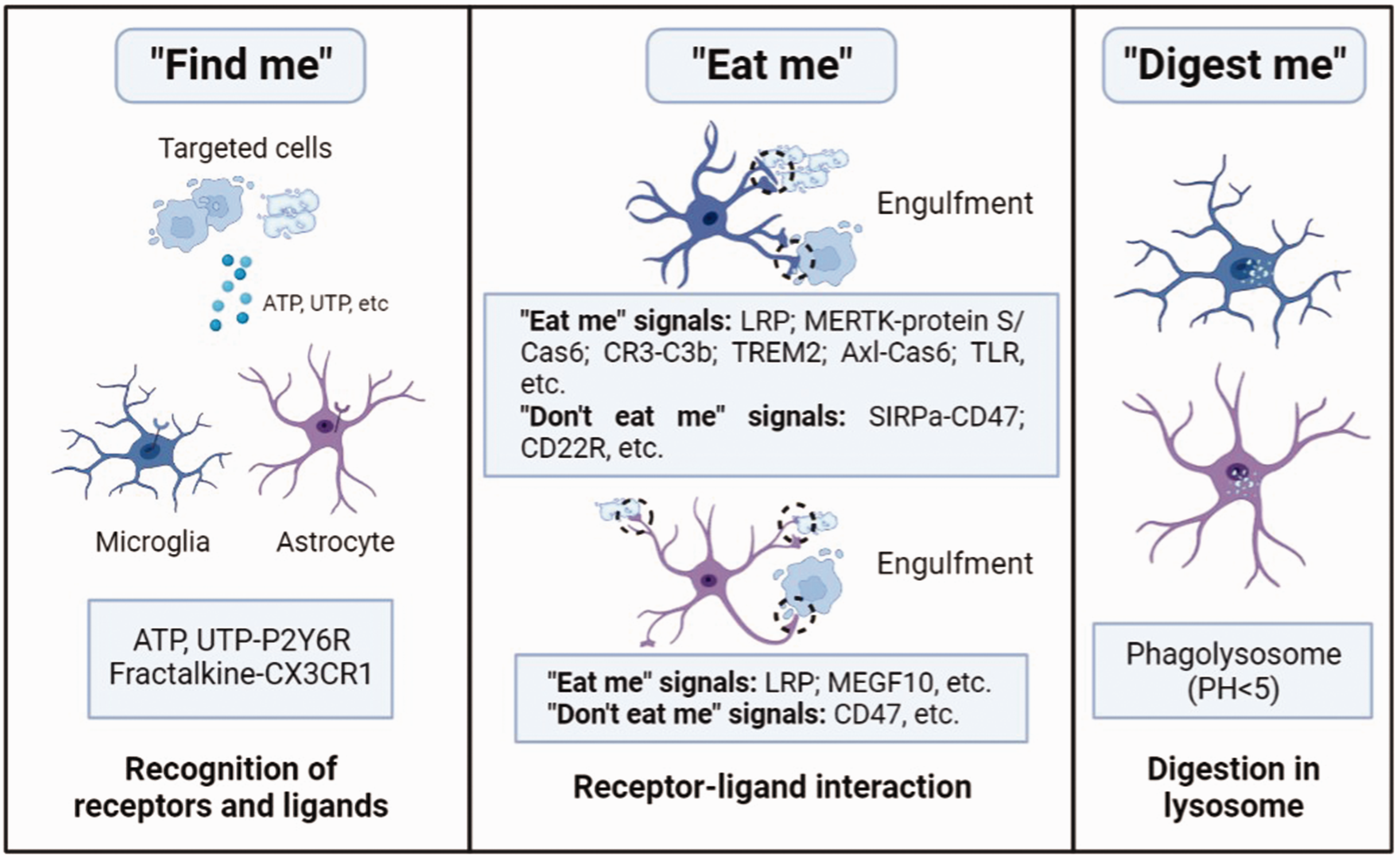
Processes involved in microglia and astrocytes-mediated phagocytosis. Phagocytic microglia and astrocytes recognize the molecules secreted by targeted cells, including ATP, UTP and fractalkine (“find me” signals). Then, the receptors in the membrane of microglia and astrocytes contact with the ligands (“eat me” signals), driving the engulfment of targeted cells and engulfed cells were delivered into the phagosome. Finally, the phagosome becomes mature, and the targeted cells are degraded in the phagolysosome within an acid environment (PH < 5).
Role of microglia-mediated phagocytosis of myelin in developmental brain
Microglia account for 10–20% of glial cells in the brain, and are considered as the main resident phagocytes in the CNS. 12 Single cell transcriptomics of microglia shows phenotypical heterogeneity in the developing brain, especially the early postnatal microglia in developing white matter recapitulate high heterogeneity with active metabolism and high phagocytic ability, which is actively involved in myelination. 34 Recently, a unique subtype of microglia defined as proliferative-region-associated microglia (PAM) is identified in postnatal day 7 (P7) mouse brain, which is critical for myelination in the developmental brain. 34 The PAM have amoeboid morphology and phagocytose newly formed OLs but not OPCs. Efficient phagocytosis of newly formed OLs by PAMs in developing white matter may be critical to provide space for later myelination. Moreover, it is reported that a subtype of microglia named “fountain of microglia”, appears only postnatally in myelinated regions such as corpus callosum and cerebellum.35,36 Hagemeyer et al. identified this specific subset of microglia arose from endogenous resident microglia pool but not circulating myeloid cells using large-scale transcriptional profiling, fate mapping, and genetic targeting approaches. 35 Microglia depletion experiments revealed an essential role of postnatal microglia in the proper development and homeostasis of OLs and OPCs. During early postnatal development, a population of ameboid microglia migrates from the ventricular zone to the corpus callosum to phagocytose OPCs before the start of myelination. 36 Fractalkine receptor-deficient mice exhibit a reduction in microglial engulfment of OPCs, increase the number of OLs, and reduced myelin thickness. Thus, microglia phagocytose OPCs might be a homeostatic mechanism for proper myelination. To further confirm the myelin engulfment of microglia during the developmental stage, Hughes et al. used zebrafish as a model to visualize and manipulate interactions between microglia and OLs. 37 They found that microglia selectively engulfed myelin sheath to shape myelination under normal physiological conditions, and the phagocytosis is bidirectionally regulated by neuronal activity. Microglia engulf more myelin from the tectal commissure when neuronal cell body layer activity is suppressed.
It has been demonstrated that microglia use classic phagocytic mechanism for engulfing myelin.12,13 Extracellular nucleotides, such as ATP and UTP, trigger microglial phagocytosis through pyrimidinergic P2Y, G-protein coupled, 6 receptor (P2Y6R) pathway, fractalkine triggers the CX3CR1 pathway. 38 Microglia target the pre-engulfed cells and contact with them via “eat me” signals. Mer tyrosine kinase (MERTK) and AXL are part of the TAM (Tyro3, AXL, MERTK) family that bind Protein S and Gas6, which have been reported to play critical roles in myelin phagocytosis. Other receptors such as low-density lipoprotein receptor-related protein (LRP), complement receptor 3 (CR3), triggering receptor expressed on myeloid cells 2 (TREM2) all participate in microglial clearance of myelin. Through the ligand-receptor interaction, phagocytosis begins, and myelin debris is rapidly digested in the phagolysosome.
Taken together, microglia phagocytose OPCs or OLs in myelinated regions during the early postnatal period for initiating and maintaining healthy myelination, as well as engulf myelin sheath to shape myelination during the developmental phase, which is regulated by neuronal activity (Figure 2).
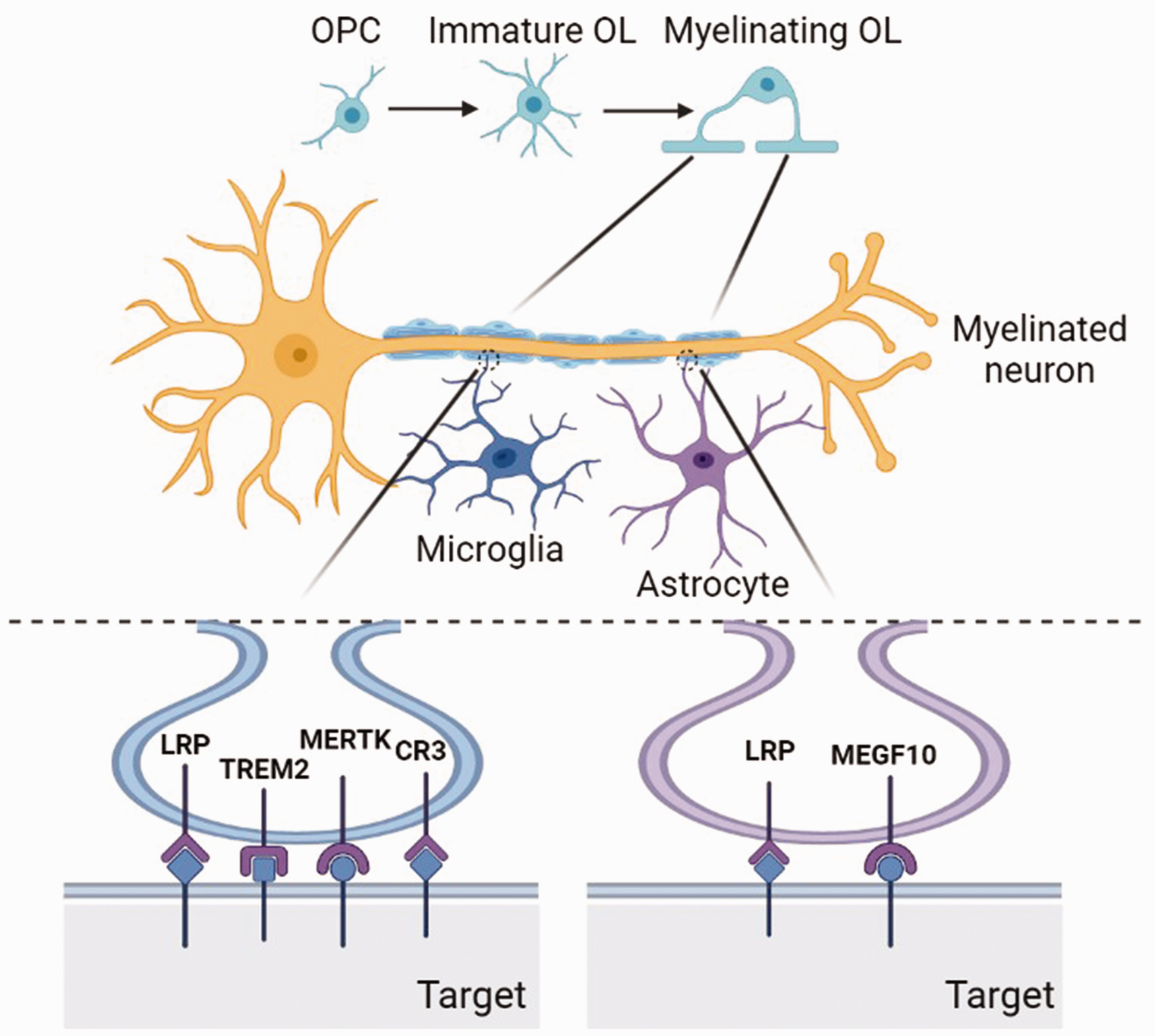
Microglia and astrocytes phagocytose myelin in the developmental brain. Myelinating OLs are derived from the proliferation, migration and differentiation of OPCs. Matured OLs generate lipidic myelin sheath. The excessive myelin needs to be cleared timely by phagocytic cells. Microglia phagocytose myelin through the receptors of LRP, TREM2, MERTK, CR3, etc. Astrocytes mediate myelin engulfment via LRP, MEGF10, etc.
Role of astrocytes-mediated phagocytosis of myelin in developmental brain
Astrocytes make up 20–40% of glial cells. 39 They are the largest glial cells in the CNS, and play an important role in supporting the function of neurons. 40 In recent years, it has been found that besides microglia, astrocytes are also phagocytic cells in the brain, and show high regional heterogeneity.15,17,41 According to the different morphology of astrocytes in gray and white matter, astrocytes are divided into protoplasmic astrocytes and fibrous astrocytes. 12 The protoplasmic astrocytes in gray matter are the biggest population, mainly found in the cortex and hippocampus. 12 Their cell bodies are extremely ramified, which allows them to contact numerous synapses in these regions, thus playing a neuro-modulatory effect. 42 The fibrous astrocytes line along white matter tracts; they are smaller and have lesser branching points and can touch the nodes of Ranvier, thus contributing to keeping the homeostasis of white matter.
Astrocytes in the white matter contribute to myelin formation and regeneration through both direct interactions with OLs, and indirect crosstalk with resident microglia. 41 These astrocytes support developmental myelination by expressing supportive factors such as CXCL1 and tissue inhibitor of metalloproteinase-1 (TIMP1). The bulges of astrocytes contact with myelinated fibers and participate in myelin turnover and maintenance in normal adult brains. In addition, it has been demonstrated that astrocytes have different morphologies in developing and adult white matter. 12 At different developmental stages astrocytes exert distinct roles in white matter myelination. GFAP+ mature astrocytes in white matter are first detected at E16, the time when OPCs begin to migrate throughout the brain for further differentiation. Myelination is accelerated during early postnatal phase, with the increase of astrocytes. Astrocytes might control the proliferation and migration of OPCs by producing platelet-derived growth factor-α (PDGFα), which is the most important survival factor for OPCs and inhibits differentiation of OPCs, thereby controlling the time of myelination during development. 12 Astrocytes also play crucial roles in maintaining healthy myelination throughout adulthood. In addition, lipid droplets or myelin membrane-like debris accumulation were observed in the lysosomes of astrocytes, indicating that astrocytes are also involved in maintaining myelin homeostasis via phagocytosing myelin. 43 There is evidence that optic nerve astrocytes can clear myelin in a nonpathological manner during development using milk fat globule epidermal growth-factor 8 (MFG-E8) or Rac family small GTPase 1 (Rac1). 44 The continuous removal of dystrophic myelin from intact axons could provide homeostasis to prevent the large-scale loss of myelin and fragmentation of axons, which triggers a pathological immune response.
Similarly, astrocytes get in touch with targeted cells, and then phagocytose myelin via receptors of LRP1, multiple EGF-like domains 10 (MEGF10), etc, to recognize “eat me” signals of phosphatidylserine (PS), etc. After the fusion of phagosome and lysosome, myelin debris is digested in acid phagolysosome.10,15 Astrocytes could engulf myelin in a LRP1-mediated endocytosis in various human demyelinating diseases. Besides, astrocytes recognize targeted myelin debris with exposed PS in the outer membrane through PS receptors (Figure 2).
Glial cell-mediated phagocytosis of myelin in the diseased brain
White matter is the major region that is enriched in myelin. 45 The internal capsule received myelinated fibers from the motor cortex to the spinal cord and is critical for movement. The largest white matter tract in the brain is corpus callosum, which is involved in the integration of sensorimotor and cognitive information. Deficits of myelination are relevant to delayed remyelination and tissue damage, and result in learning and memory deficits. 46 Numerous CNS diseases cause white matter injury, including Alzheimer’s disease (AD), multiple sclerosis (MS), amyotrophic lateral sclerosis (ALS), cerebral ischemia, etc. In the damaged myelinated areas, remyelination is the key point for initiating white matter repair, which requires the activation, migration, proliferation and differentiation of OPCs before the final formation of new myelin sheaths. 47
During demyelination, the amount of myelin debris in the white matter is closely associated with inflammation, and inefficient clearance of myelin debris impairs remyelination, suggesting the balance between remyelination and demyelination is critical for the recovery of demyelinating diseases. Thus, timely removal of myelin debris is crucial for remyelination process. Accumulating evidence shows that glial cells, including microglia and astrocytes, are actively involved in the clearance of myelin debris, and the prevailing view is that myelin debris clearance by phagocytic cells is beneficial to myelination. However, recent studies argue that over-activated glial cells cause excessive myelin engulfment, which may contribute to the secondary brain damage.12,48
Role of microglia-mediated phagocytosis of myelin in diseased brain
Role of microglia-mediated phagocytosis of myelin in neurodegenerative disease
Multiple sclerosis (MS)
In demyelinated diseases, efficient clearance of myelin debris or degenerated myelin is vital to rebuilding a beneficial environment for remyelination. Myelin debris contains substances for inhibiting OPC recruitment and differentiation, and its clearance is important for myelinating OLs. 47 MS is an inflammatory, demyelinating, and neurodegenerative disease of the CNS triggered by autoimmune mechanisms. 49 Experimental autoimmune encephalomyelitis (EAE) is the most commonly used animal model to mimic MS with extensive axonal degeneration and T cell infiltration in the CNS, often induced by proteins of MBP, PLP or MOG. 50 Another model is the cuprizone model, with diffuse white matter injury and no classical T cell infiltration and blood-brain barrier (BBB) impairment. 51
It is well known that microglia-mediated phagocytosis of myelin debris in the demyelination area is critical for promoting remyelination in MS disease. By adding 14 C-labeled myelin to cultured microglia, Fu and colleagues found that tumor necrosis factor (TNF)-α and TNF-γ enhanced myelin debris clearance. 12 Besides, receptors of fragment crystallizable (Fc), complement and Galectin-3/MAC-2 have also been regarded as important mediators in microglia-mediated myelin phagocytosis in vitro. 52 Cytokines and adhesion molecules have been demonstrated to regulate the phagocytic activity of microglia. For example, IL-4 and IL-10 increased phagocytosis of microglia, and reduced inflammatory response. 12 rHIgM22, an experimental human recombinant IgM antibody, promotes remyelination in cuprizone-mediated animal models of MS by enhancing microglia-mediated clearance of myelin debris in vitro in a dose-dependent manner, and this process requires actin polymerization, activation of rHIgM22 Fc domain and CR3. 53 In addition, phase 1 clinical trial showed that single infusion of rHIgM22 was well tolerated by MS patients, suggesting its safety for clinical translation. TREM2 is another molecule highly expressed on myelin-laden phagocytic microglia in demyelinating lesions of MS. 9 Treatment with TREM2 agonist promotes myelin uptake and degradation, resulting in accelerated myelin debris removal by microglia in the cuprizone mouse model. Importantly, antibody-dependent TREM2 activation on microglia increases OPCs density in the demyelination area, as well as the formation of OLs, thus enhancing remyelination and axonal integrity. Tyro3, Axl and MERTK are TAM receptors with tyrosine kinase activity that bind to the “eat-me” signal on cell membranes to initiate phagocytosis. Using a cuprizone-mediated demyelination mouse model and single-cell RNA sequencing, Shen and colleagues found that knockout of MERTK impaired myelin debris clearance and remyelination. 54 The accumulated myelin debris triggered the secretion of interferon gamma (IFN-γ) by microglia, as well as inhibited microglial phagocytosis and remyelination. Axl functions in the recruitment of microglia to the lesion brain and clearance of debris following demyelination. In an EAE model, loss of the receptor tyrosine kinase Axl leads to enhanced inflammation and delayed removal of myelin debris. 55 It is observed that T cells infiltrated into the brain in MBP-induced MS model, which could enhance the microglial response to axonal damage and microglial phagocytosis of myelin debris associated with the degenerating axons. 56 The increased phagocytosis of myelin debris might promote regeneration after T cell-mediated immune response in MS. It is reported that LRP1 is expressed by microglia and serve as an essential receptor for myelin phagocytosis in MS. 57 Using extracted myelin vesicles (MV), Gaultier et al. found that microglia expressed LRP1 and mediated MV uptake, which could be inhibited by blocking the interaction of LRP1 with other ligands. Using single-nucleus RNA sequencing to assess transcriptomics of multiple cell lineages including neurons, OLs, microglia and astrocytes in MS patients, Schirmer et al. found that there is a cluster of microglia related to phagocytosis and enriched in OL-specific markers PLP1, MBP, suggesting myelin are digested by phagocytic microglia in MS. 58 However, the role of this cluster of microglia remains unclear in MS patients.
Amyotrophic lateral sclerosis (ALS)
ALS is a neurodegenerative disease characterized by the degeneration of both upper motor neurons and lower motor neurons, leading to muscle weakness and eventual paralysis. 59 Efficient removal of degenerated neurons or cell debris is necessary to rebuild an appropriate environment for neuronal regeneration. 12 In ALS, damage-associated molecular pattern (DAMP) arising from neuronal debris can directly trigger inflammation and activate microglia. Activated microglia aggregate around the lesioned area, exhibiting potential phagocytic activity. Enhancement of myelin phagocytosis by microglial promotes neuro-regeneration and restoration of locomotor function. In an ALS mice model of SOD1G93A, the neutralization of voltage-gated proton channel HVCN1 with antibody in brain promotes migration of microglia and improves myelin debris clearance, providing a potential strategy to ameliorate the symptom of ALS. 60
Alzheimer’s disease (AD)
Excessive engulfment of myelin sheath will bring negative effects on tissue repair. The major pathologies of AD are the amyloid beta (Aβ) deposition, and the neurofibrillary tangle formed by hyperphosphorylated tau.61,62 Demyelination extensively occurred in AD brain, not only in the area full of Aβ plaques but also in the adjacent periphery. 63 The mechanism of demyelination may relate to the activation of microglia and over-phagocytosis of oligodendrocytes by microglia.
In conclusion, the myelin debris clearance of microglia mediated by CR3, TRME2, MERTK, Axl, LRP1, etc. brings a positive effect on remyelination in MS and ALS. However, the hyper-phagocytosis of myelin by microglia may also cause severe demyelination in AD. Therefore, microglia-mediated phagocytosis of myelin plays different roles in different diseases (Figure 3).
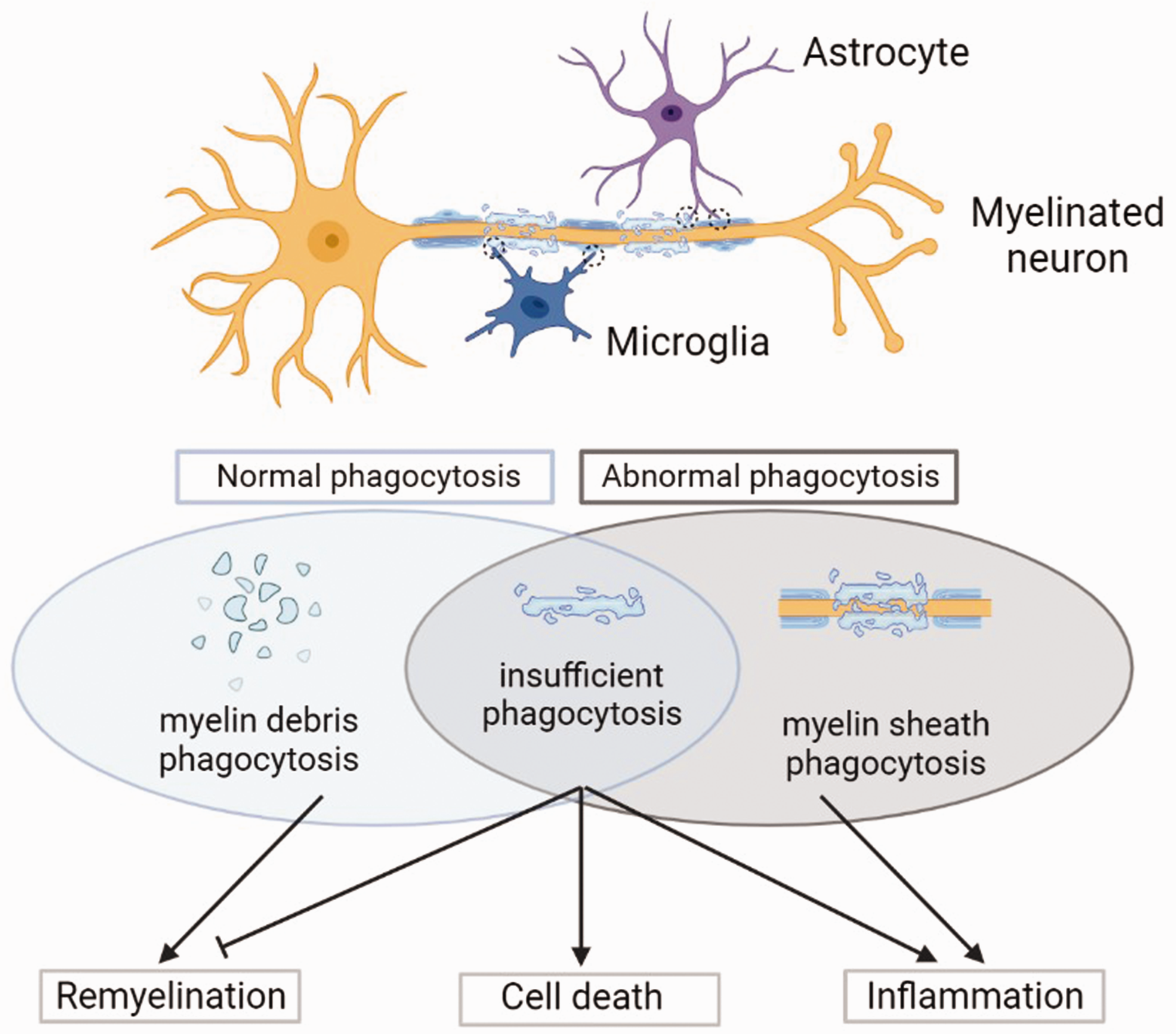
Myelin phagocytosis of microglia and astrocytes in CNS diseases. In diseased brain, microglia and astrocytes are activated and become phagocytic phenotypes. Myelin phagocytosis brings two side effects. On one hand, the normal engulfment of myelin debris will attenuate inflammation and be beneficial to remyelination. On the other hand, the abnormal or insufficient engulfment will trigger severe inflammation or even cell death and inhibit remyelination.
Role of microglia-mediated phagocytosis of myelin in cerebrovascular diseases
Cerebrovascular disease includes stroke, carotid stenosis, aneurysms and vascular malformations, etc., in which an area of the brain is temporarily or permanently affected by ischemia or bleeding. 64 Stroke is the second cause of death and the leading cause of disability worldwide and can be divided into ischemic stroke and hemorrhagic stroke. 65 After ischemia injury, microglia become activated, rapidly migrate to the lesion area, and engulf live or dead neurons, neuronal synapses, myelin debris, myelin sheath, apoptotic cells, and leukocytes, triggered by the exposure of “eat-me” signals or the loss of “don’t eat-me” signals, and ultimately affect stroke outcome. 66 After stroke, white matter injury accounts for nearly half of ischemic injuries, but few studies are focused on the roles of microglia-mediated myelin clearance in ischemic stroke.67–70 Microglial phagocytosis is initiated similarly with related receptors under pathological circumstances. CR3 is involved in the microglial phagocytosis of myelin debris. 71 Pseudoginsenoside-F11 (PF11), an ocotillol-type saponin, could promote microglia-mediated phagocytosis of myelin debris in both oxygen-glucose deprivation (OGD) model in vitro and permanent middle cerebral artery occlusion (pMCAO) model in vivo through CR3 and reduce the release of inflammatory factors, which attenuates cerebral ischemic injury. TREM2 is also an important receptor in microglia-induced phagocytosis, and it could mediate microglial migration to engulf myelin debris and apoptotic cells, but its role in stroke remains unclear. 72 Our group previously demonstrated that microglia-mediated synapse phagocytosis through MEGF10 and MERTK in both ischemic stroke and hemorrhagic stroke, inhibiting the phagocytosis improved neurobehavioral outcomes and attenuated brain damage. 73 The functions of MEGF10 and MERTK on myelin phagocytosis are still unknown.
Chronic cerebral hypoperfusion (CCH) is a cerebrovascular disease that causes white matter injury, and contributes to cognitive impairment and vascular dementia. 24 In CCH, aberrantly activated microglia exacerbate white matter injury via C3-C3aR pathway to hyper-engulf myelin sheath and aggravate neuroinflammation in bilateral common carotid artery occlusion (BCCAO) rats. 24 Genetic deletion of C3aR significantly inhibited the activation of microglia and reversed white matter injury. Moreover, C3aR antagonist SB290157 decreased the number of microglia involved in myelin phagocytosis, and attenuated cognitive deficits in chronic hypoperfusion rats. However, other receptors involved in myelin engulfment by microglia in CCH remain to be studied in the future.
Role of microglia-mediated phagocytosis of myelin in other CNS diseases
SCI often results in severe functional impairment of the limbs below the injured level. 74 Myelin debris is a hallmark of SCI. 75 Acute SCI disrupts vasculature, leading to tissue ischemia and BBB breakdown and generating myelin debris. Myelin injury occurs at the onset of the injury, and cellular debris is present mainly in areas of damaged white matter within 24 hours. Myelin debris increases within the first week of SCI and persists in the chronically injured spinal cord for years. Degraded proteins and lipids in myelin debris exacerbate the inflammatory response of microglia after SCI. 76 On the other hand, the presence of myelin debris inhibits axonal regeneration, OPC differentiation, and remyelination. 77 CR3/MAC-1 signaling pathway was involved in the clearance of degenerated myelin. The increased expression of surface molecules, including ICAM-1, CD14, CD44, and CD45, upregulated phagocytosis, and significantly enhanced microglia-mediated tissue debris removal. 78 Myelin structures, such as Nogo-A, myelin-associated glycoprotein, oligodendrocytes myelin glycoprotein, the transmembrane semaphoring 4 D (Sema4D/CD100), and ephrinB3, locate on injured myelin and act as inhibitors of axonal regeneration. Wu et al. demonstrated that metformin promoted the clearance of myelin debris by microglia and accelerated nerve repairment after SCI with the activation of microglia and the attenuation of inflammatory response. 74 Therefore, efficient microglial phagocytosis of damaged myelin debris is beneficial for repairing injured neurons and myelin regeneration in acute SCI, resulting in prominent functional recovery. CD47, a “don’t eat me” signal, was demonstrated to express on myelin. When CD47 reacted to immune inhibitory receptor signal regulatory protein-α (SIRP-α) on microglia, phagocytosis of myelin was reduced. 13 Toll-like receptor (TLR) 4 signaling plays a critical role in inducing OLs genesis in the naive spinal cord, and lack of TLR4 signaling inhibited white matter recovery after SCI, which is due to delayed myelin phagocytosis. 79
TBI is a brain disease with characteristics of progressive neurodegeneration, sustained neuroinflammation and chronic neurological dysfunction. 80 It is often accompanied by diffuse axonal injury, and there is chronic upregulation of Galectin-3 in microglia in the corpus callosum. 81 There is evidence of TBI-induced activation of microglia and the hyper-phagocytosis of myelin debris, which may be consistent with the pathology of TBI. Following diffuse TBI, Galectin-3+ microglia engulfed damaged axons and myelin, which may contribute to the secondary injury to TBI. The molecular signals that regulate myelin phagocytosis after TBI remain to be addressed.
In the aging CNS, myelin debris was gradually released from aged myelin sheaths and subsequently cleared by microglia. 11 Myelin fragmentation increased with aging and led to the formation of insoluble, lipofuscin-like lysosomal inclusions in microglia. The reduced number, monitoring capacity, and phagocytic activity of aged microglia led to a decreased ability to clear myelin debris, which in turn resulted in slower remyelination compared with the young CNS.82,83 Besides, in a study of simulated demyelination using a lysophospholipid demyelinating toxin injection, it was observed that middle-aged mice recruited fewer microglia than young mice at 7 days after demyelination and did not reach the same level as young mice until 21 days post-injury. 82 Moreover, histological Oil Red O staining indicated that phagocytosis of lipids in middle-aged mice was lower than that of young mice at 7 days after demyelination. In a demyelination model, aged mice failed to resolve the inflammatory response initiated after myelin injury. 84 Aged phagocytic microglia accumulated excessive amounts of myelin debris, which triggered cholesterol crystal formation and phagolysosomal membrane rupture and stimulated inflammasomes. Thus, age-associated decline in engulfment capacity of microglia limits the restoration functionality in CNS after injury. Using single-cell RNA sequencing from white matter and gray matter of aged mice, Safaiyan et al. identified white matter-associated microglia (WAMs), which share parts of the disease-associated microglia (DAM) gene signature and are characterized by activation of genes implicated in phagocytic activity and lipid metabolism. 85 WAMs depend on TREM2 signaling and are aging-dependent. In the aged brain, WAMs are independent of apolipoprotein E (APOE). Within the white matter, microglia frequently cluster in nodules, where they are engaged in clearing degenerated myelin. In this case, they propose that WAMs represent a protective metabolic response required to clear the increasing amounts of myelin debris that accumulate during aging. However, the harmful functions, such as immune reactivity, could not be excluded, for the WAM signature is also associated with phagosome as well as antigen processing and presentation. CD22 (also known as a “don’t eat me” signal) was discovered as an age-related genetic modifier of microglial phagocytosis via the combination of CRISPR-Cas9 knockout screens with RNA sequencing analysis. 86 CD22 is a negative regulator of phagocytosis that is upregulated on microglia after aging. The inhibition of CD22 promotes the clearance of myelin debris, amyloid-β oligomers and α-synuclein fibrils in vivo. Long-term CNS delivery of an antibody that blocks CD22 reprograms microglia to a homeostatic transcriptional condition and improves cognitive function in aged mice. 86
Microglia-mediated phagocytosis of myelin also plays a critical role in other demyelinated injuries. PLX5622, an inhibitor of colony stimulating factor 1 receptor (CSF1R), was used to deplete microglia in mice infected with a neurotropic coronavirus, resulting in impaired myelin repair and prolonged clinical disease. 87 Large deposits of extracellular vesiculated myelin and cellular debris were detected in the spinal cords of PLX5622-treated mice, and this could not be compensated by infiltrating macrophages. Activated microglia contribute to pathogenesis of demyelination, and regulate viral replication and IFN-γ production during the acute phase of hepatitis virus (MHV) infection. 88 IFN-γ can in turn activate microglia by promoting phagolysosome maturation and engulfment of the myelin sheath, leading to demyelination. The studies in different CNS diseases present the Janus-faced roles of myelin phagocytosis by microglia. Molecules involved in microglia-mediated phagocytosis of myelin after CNS injuries are summarized in Table 1.
Receptors involved in microglia-mediated phagocytosis of myelin.
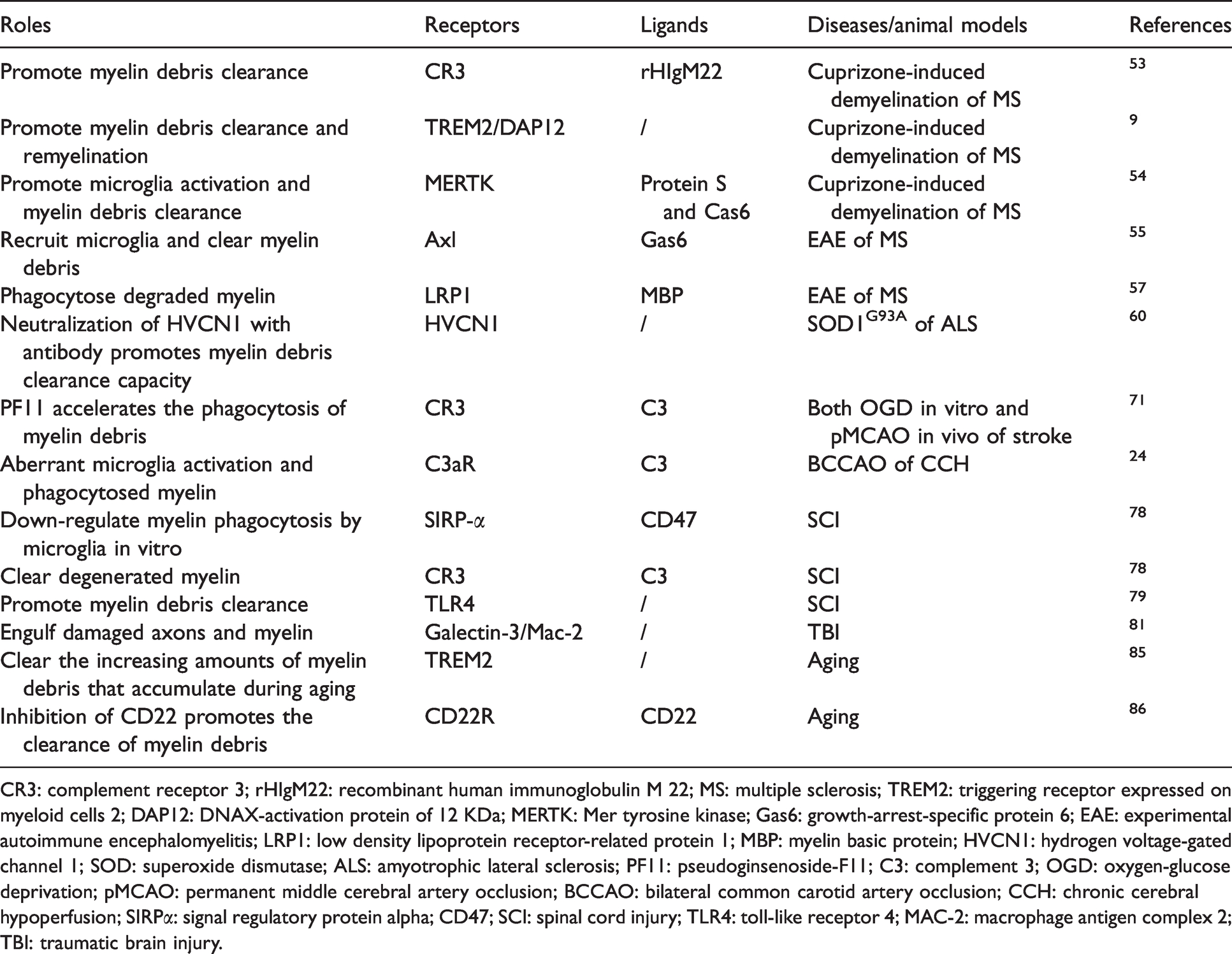
CR3: complement receptor 3; rHIgM22: recombinant human immunoglobulin M 22; MS: multiple sclerosis; TREM2: triggering receptor expressed on myeloid cells 2; DAP12: DNAX-activation protein of 12 KDa; MERTK: Mer tyrosine kinase; Gas6: growth-arrest-specific protein 6; EAE: experimental autoimmune encephalomyelitis; LRP1: low density lipoprotein receptor-related protein 1; MBP: myelin basic protein; HVCN1: hydrogen voltage-gated channel 1; SOD: superoxide dismutase; ALS: amyotrophic lateral sclerosis; PF11: pseudoginsenoside-F11; C3: complement 3; OGD: oxygen-glucose deprivation; pMCAO: permanent middle cerebral artery occlusion; BCCAO: bilateral common carotid artery occlusion; CCH: chronic cerebral hypoperfusion; SIRPα: signal regulatory protein alpha; CD47; SCI: spinal cord injury; TLR4: toll-like receptor 4; MAC-2: macrophage antigen complex 2; TBI: traumatic brain injury.
Role of astrocytes-mediated phagocytosis of myelin in diseased brain
Role of astrocytes-mediated phagocytosis of myelin in neurodegenerative disease
Multiple sclerosis (MS)
Recently, astrocytes have been proven as another important phagocytic cell in the brain. Astrocytes proliferate and become activated within demyelinating lesions after MS, suggesting their critical roles in white matter injury and axonal degeneration. 89 Activated astrocytes exert effects in two ways. On one hand, astrocytes contribute to the clearance of myelin debris in demyelination area, through both phagocytosis of myelin and recruitment of macrophages and microglia. On the other hand, they express calpain, a protein that is involved in the degradation of axonal and myelin proteins, and neurofilament proteins, located in or near inflammatory areas. Astrocytes are reported to be linked to rapidly progressive MS. 90 Within early-active and active lesions, hypertrophic astrocytes are involved in lesion pathology through the phagocytosis of myelin debris. Thus, hypertrophic astrocytes not only play roles in the pathogenesis of MS lesions, but also exert persistent harmful effects on tissues in the absence of severe inflammation. By analyzing purified astrocytes, Cahoy et al. for the first time finds that astrocytes are enriched in phagocytosis-associated pathways, such as LRP1, MEGF10, MERTK, which may play roles in myelin engulfment. 91 After stimulation by activated microglia, a phenotype of neurotoxic reactive astrocytes reduces the capacity to clear myelin debris with the downregulation of MEGF10 and MERTK. 92 In human brain tissue samples of MS, myelin debris have been shown to be taken up by astrocytes via LRP1-mediated endocytosis and lead to astroglial NF-κB activation and chemokines secretion, resulting in immune cells recruitment. 10 This response of astrocytes occurs in an early stage of MS, which may affect the lesions. Activated astrocytes also produce chondroitin sulfate proteoglycans (CSPGs) in the MS lesions, which have been found accumulated in the astrocytic scar and inhibit remyelination. 93 CSPGs consist of a protein core with one or more covalently linked chondroitin sulfate chains. 94 As major components of extracellular matrix, CSPGs will rapidly response when CNS disease happens, and are critical for modulating remyelination. Remyelination in the CNS is negatively associated with the number of reactive astrocytes and level of CSPGs.95,96 After MS, a number of CSPGs are increased following demyelination, and inhibit growth and differentiation of OPCs and OLs, and result in impeding remyelination. 95 In addition, CSPGs reduced morphological differentiation, adhesion, and the growth of human OPCs and OLs in vitro. 97 The demyelinating effect could be reversed via inhibiting CSPGs. It has been proved that preventing CSPGs deposition in damaged area could promote growth of OPCs in vitro and accelerate remyelination of MS mice. Therapies targeting CSPGs such as block CSPGs synthesis, 93 digesting glycoaminoglycan (GAG) chains, 98 inhibiting CSPG signaling pathway, 99 or disrupting CSPG receptors, 100 have been shown to promote remyelination and axon regeneration, suggesting CSPGs is an important therapeutic target for MS.
Role of astrocytes-mediated phagocytosis of myelin in cerebrovascular disease
In stroke brain, both microglia and astrocytes were activated and showed phagocytic phenotypes to differentially contribute to the remodeling of brain tissue after ischemic stroke. 101 Compared to microglial phagocytosis, astrocytic phagocytosis displayed a distinct spatiotemporal pattern. Phagocytic microglia are mainly observed in the core area at the early stage of stroke, while phagocytic astrocytes are firstly discovered at day 3, peaked at day 7, and existed at least 14 days after MCAO mainly in the penumbra region. Astrocytes could mediate the clearance of neuron debris through upregulating phagocytic pathway molecules of ATP-binding cassette transporter A1 (ABCA1), engulfment adapter phosphotyrosine binding domain containing 1 (GULP1), MEGF10, etc. In another rat model of photochemically induced focal cerebral ischemia, transforming growth factor (TGF)-β2 suppressed microglia-mediated myelin engulfment during injury. 102 Recently, Wan and colleagues found that LCN2/LRP1 regulates astrocyte-mediated myelin phagocytosis in a mouse stroke model of distal MCAO (dMCAO). 103 After stroke, reactive astrocytes derived-LCN2 was upregulated, mediating astrocytic phagocytosis of myelin in non-ischemic areas of the corpus callosum, which causes progressive demyelination. Knockdown of LCN2 or its receptor LRP1 reduced astrocytes-induced myelin engulfment and reduces demyelination. In a rat two vessel occlusion (2VO) model of CCH, it was demonstrated that physical exercise alleviated cognitive impairment, improved white matter repair, and promoted astrocytes polarization to neuroprotective phenotype to facilitate myelin debris clearance and remyelination. 104
Role of astrocytes-mediated phagocytosis of myelin in other brain diseases
In aged non-human primates, the presence of phagocytosed myelin in astrocytes and short internodal myelin provided clues for the remyelination process in aging brain. 105 In SCI, 106 astrocytes directly clear myelin debris through endocytosis pathways and followed by excessive gliosis, which inhibit axon regeneration and contribute to further tissue damage. In patients with Creutzfeldt-Jakob disease (CJD), many hypertrophic astrocytes were found to engulf one to several OLs within their cytoplasm (“emperipolesis”) in injured white matter. 14 The presence of emperipolesis in the white matter in CJD was correlated with the severity of disease. It should be noted that “emperipolesis” may not mean phagocytosis, as the limited phagocytic ability of astrocytes and the lack of evidence of intracellular digestion, thus it needs further study to verify the roles of astrocytes in mediating myelin phagocytosis in CJD. Key molecules involved in myelin phagocytosis of astrocytes after CNS injuries are summarized in Table 2.
Receptors involved in myelin phagocytosis of astrocytes.
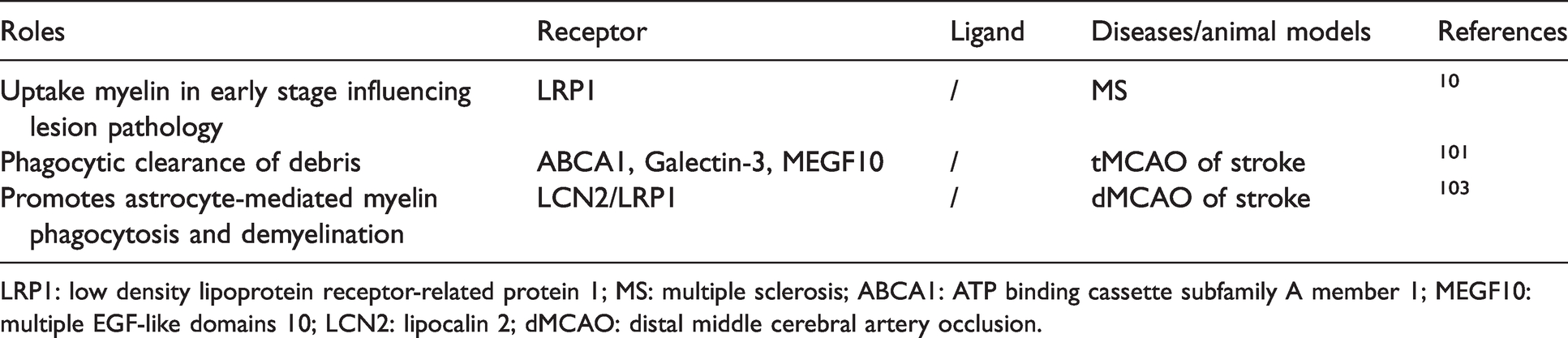
LRP1: low density lipoprotein receptor-related protein 1; MS: multiple sclerosis; ABCA1: ATP binding cassette subfamily A member 1; MEGF10: multiple EGF-like domains 10; LCN2: lipocalin 2; dMCAO: distal middle cerebral artery occlusion.
Interaction of microglia and astrocytes in myelin phagocytosis
The interaction between microglia and astrocytes affects their phagocytotic ability and disease progression after CNS injuries. Microglia co-cultured with astrocytes phagocytosed more myelin than cultured alone. 107 Astrocyte-conditioned medium treatment also increased phagocytic activity of microglia by 3.5-fold. The results reveal that astrocytes may secret growth factors to promote microglia survival and proliferation, which may contribute to enhancing the ability of microglial phagocytosis. However, the phagocytic capacity of microglia was significantly reduced in the presence of both neurons and astrocytes in vitro. 108 Neurons and astrocytes reciprocally orchestrate microglial homeostasis, which may be regulated by the transforming growth factor beta 2 (TBGF-β2) signaling. The different results suggested the complexity of microglia-astrocyte interaction. Astrocytes may have beneficial effect on OL regeneration during remyelination in an experimental demyelination model induced by cuprizone through the interaction with microglia. 109 Skripuletz et al. demonstrated that astrocytes recruit microglia to clear myelin debris in a cuprizone-induced rodent demyelination model, which is regulated by chemokine CXCL10. 110 Ablation of astrocytes using glial fibrillary acidic protein-thymidine kinase transgenic mice caused a decrease in microglial activation, resulting in the failure of damaged myelin removal and a consecutive delay in remyelination. However, using the same animal model, Madadi and colleagues revealed that ablation of astrocyte by La-aminoadipate (L-AAA) brought an improvement of myelination via hampering microgliosis and astrogliosis. 111 What’s more, Petkovic and colleagues found that astrocyte-targeted production of IL-6 in cuprizone-treated mice led to significant reduction in astroglial and microglial activation and accumulation, as well as CXCL10, CXCL1 and CCL4 levels in the corpus callosum. In addition, the microglial receptor involved in myelin phagocytosis including TREM2 and CD68 had lower expression, causing impaired removal of degraded myelin. Reduced microglial accumulation resulted in insufficient removal of degraded myelin, impaired early OL differentiation and remyelination 112 (Figure 4). In ICH, a study revealed that ICH induced neurotoxic reactive microglia and astrocytes through ROS-induced NF-κB p65 translocation that impaired OPC maturation, neurotoxic astrocytes inhibit microglia-mediated phagocytosis of myelin debris via astrocytic C3-microglial C3aR axis. 113 In turn, microglia may affect the phagocytosis of astrocytes via cytokines. It is reported that activated microglia induce neurotoxic reactive astrocytes by secreting Il-1α, TNF and C1q, and this subtype of astrocytes lose the ability of engulfing myelin debris, which may cause the death of OL. 92 However, the interaction of microglia and astrocytes in myelin phagocytosis in vitro and in vivo needs more investigation.
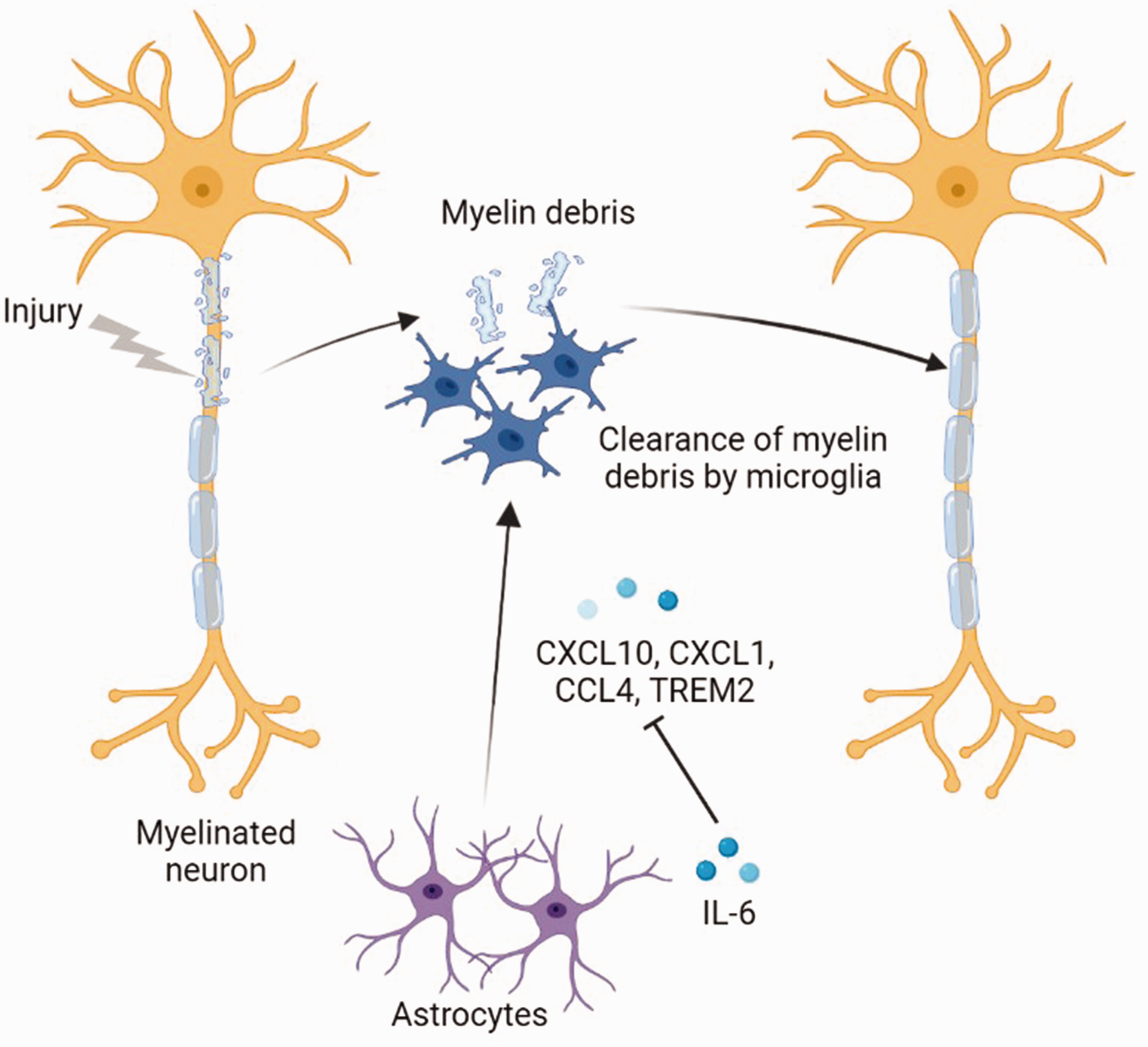
Interaction of microglia and astrocytes in myelin phagocytosis in CNS diseases. After CNS injury, it will produce myelin debris causing tissue damage. Astrocytes could recruit microglia to clear myelin debris for further remyelination through CXCL10. Moreover, the secreted IL-6 by astrocytes could inhibit the production of microglial attracting chemokine CXCL10, CXCL1 and CCL4, and the phagocytosis associated receptor TREM2.
Conclusions and perspectives
In summary, microglia and astrocytes are key phagocytic cells in the CNS, which mediate myelin engulfment for maintaining internal microenvironment of the CNS. In developmental brain, microglia and astrocytes engulf OLs, OPCs or unnecessary myelin sheath to form healthier myelination. Under disease conditions, phagocytic cells exert roles in myelin debris clearance for better remyelination, which is beneficial to tissue repair. However, the excessive engulfment of myelin sheath or insufficient clearance of myelin debris will trigger inflammation and inhibit remyelination, resulting in worse outcomes. What’s more, astrocytes and microglia interact with each other and coordinately regulate myelin phagocytosis. There are several key issues that still need to be investigated to reveal the roles of glial cell-mediated phagocytosis of myelin in CNS.
Except for the above-mentioned molecules, other molecules that regulate glial cell-mediated myelin phagocytosis need to be continually explored in order to find more therapeutic targets. Different molecules may play synergistic or antagonistic action on myelin phagocytosis, thus the interaction between different molecules also requires further investigation. Nowadays, most attractions related to myelin phagocytosis focus on the study of typical demyelinated diseases, such as MS and SCI. While it may also play critical roles in other CNS diseases including AD, stroke, etc., as the myelin integrity is essential to axonal and tissue recovery. Moreover, astrocytes have been highlighted with their phagocytosis capacity as a supplementary mechanism of microglia-mediated phagocytosis, but their roles in myelin engulfment and how they affect remyelination remain unclear. Macrophages are often seen as critical phagocytic cells as microglia, especially after BBB is impaired in diseased brain. It has been reported that macrophages could phagocytose myelin and neuronal fragments, which is beneficial to remyelination and axonal repair or regrowth.83,114 Some researchers point out that the contribution of myelin phagocytosis by macrophages is more important than microglia,
83
but most studies don’t distinguish their roles in myelin phagocytosis. The distinct effects of microglia and macrophages on myelin phagocytosis need more exploration. Recently, the routinely used methods to investigate phagocytosis include immunohistochemistry (IHC), transmission electron microscope (TEM), and flow cytometry. IHC is easy and convenient, but it cannot confirm if myelin debris is inside the glial cells. TEM is intuitive and accurate, but it is time-consuming and cost-effective. Flow cytometry can be used to quantitatively evaluate the phagocytosis of myelin debris by labeling myelin debris with fluorescent dyes or indicators. PHrodo, a PH sensitive indicator that could show fluorescence if myelin debris is digested in lysosome.115,116 Very recently, adeno-associated virus (AAV) carrying mcherry and GFP under different promoters are used to assess phagocytosis based on the fact that lysosomal hydrolases degrade GFP first so that GFP fluorescence is attenuated in lysosomes, whereas mcherry fluorescence is more stable in lysosomes. However, it takes several weeks for virus transfection, and not every synapse or myelin can be evenly labeled by the virus. Thus, developing more quantitative and precise methods to evaluate phagocytosis will provide a powerful platform for phagocytosis research in future. The accumulated myelin debris in cell bodies will trigger severe inflammation and have detrimental effects on remyelination.
10
Therefore, we should also take attention to the transportation and molecules involved in the digestion and degradation process after engulfment.
All in all, the myelin phagocytosis of microglia and astrocytes indicate potential effects on myelination, which may become a targeted therapeutic strategy for CNS intervention and diseases.
Footnotes
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by grants from National Key R&D Program of China #2019YFA0112000 (YT), the National Natural Science Foundation of China (NSFC) projects 82071284 (YT), Shanghai Rising-Star Program 21QA1405200 (YT), the Scientific Research and Innovation Program of Shanghai Education Commission 2019-01-07-00-02-E00064 (GYY), Science and Technological Innovation Act Program of Shanghai Science and Technology Commission, 20JC1411900 (GYY).
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.