
Abstract
In most positron emission tomography (PET) molecular brain imaging studies, regions of interest have been defined anatomically and examined in isolation. However, by defining regions based on physiology and examining relationships between them, we may derive more sensitive measures of receptor abnormalities in conditions such as major depressive disorder (MDD). Using an average of 52 normalized binding potential maps, acquired using radiotracer [11C]-WAY100635 and full arterial input analysis, we identified two molecular volumes of interest (VOIs) with contiguously high serotonin 1A receptor (5-HT1A) binding sites: the olfactory sulcus (OLFS) and a band of tissue including piriform, olfactory, and entorhinal cortex (PRF). We applied these VOIs to a separate cohort of 25 healthy control males and 16 males with MDD who received [11C]-WAY100635 imaging. Patients with MDD had significantly higher binding than controls in both VOIs, (p < 0.01). To identify potential homeostatic disruptions in MDD, we examined molecular connectivity, i.e. the correlation between binding of raphe nucleus (RN) 5-HT1A autoreceptors and post-synaptic receptors in molecular VOIs. Molecular connectivity was significant in healthy controls (p < 0.01), but not in patients with MDD. This disruption in molecular connectivity allowed identification of MDD cases with high sensitivity (81%) and specificity (88%).
Introduction
According to conventional practice, ligand binding in the brain is quantified in anatomically defined regions determined a priori. However, these atlas-based regions may not respect the regional distribution of the binding site in question, resulting in insensitivity for detecting disease-related focal disturbances by positron emission tomography (PET), leading to equivocal results in, for example, PET studies of serotonin 5-HT1A receptors in major depressive disorder (MDD). Therefore, in our study, we delineate volumes of interest (VOIs) using a data-driven approach based on serotonin 1A (5-HT1A) receptor distribution, rather than by a priori anatomical landmarks. We are able to delineate such regions from a repository of >50 subjects previously imaged with the radiotracer [11C]-WAY100635 and full arterial input analysis. Mean, fully quantitative binding potential maps generated from these subjects resulted in two clear regions of high 5-HT1A density that do not overlap completely with anatomically defined regions. This method of VOI delineation presents a more accurate picture of receptor binding since receptor distribution may only comprise part of an anatomical region, or conversely, it may span several adjacent regions.
In addition, we evaluate the relationship between presynaptic 5-HT1A receptors in the raphe nucleus (RN) and post-synaptic receptors in the aforementioned molecular VOIs. As RN autoreceptors regulate serotonergic transmission in the brain, we would expect post-synaptic receptors to upregulate and downregulate in proportion to the number of autoreceptors. Examining the relationship between 5-HT1A density in the RN and these postsynaptic regions, therefore, could provide unique insight into receptor homeostasis and responsivity in MDD. We term the coupling of binding observed between the RN and these two sets of molecularly defined VOIs, akin to functional connectivity, molecular connectivity. Combining PET-derived VOI delineation and evaluation molecular connectivity, we evaluate PET data in an innovative way that sheds additional light on the pathophysiological mechanisms of MDD.
MDD is a debilitating psychiatric illness and one of the top causes of years lost to disability worldwide. 1 However, our understanding of its pathophysiology is limited, as evidenced by poor remission rates using conventional treatments. 2 One of the prevailing models of MDD pathophysiology involves the serotonergic system, and in particular the 5-HT1A receptor. 5-HT1A autoreceptors are found on the RN, the major source of serotonin in the central nervous system. When activated, these receptors inhibit RN neurons, 3 thus decreasing the overall amount of serotonin released. 4 Post-synaptic 5-HT1A receptors are found throughout the brain, particularly in areas such as the hippocampus, insula, and temporal pole. 5
PET has been used to measure 5-HT1A receptor density in vivo using radiotracers such as [11C]-WAY100635, 6 a 5-HT1A antagonist. In support of 5-HT1A’s role in MDD, PET studies from our lab have found markedly higher 5-HT1A binding potential in the RN in depressed human subjects compared to controls.7,8 However, differences in post-synaptic regions have not been as robust.
One reason for this may be that, while the anatomic boundaries of the RN have been clearly defined by their 5-HT1A somatodendritic distribution, the proper delineation of post-synaptic receptors has been less clear. Researchers often rely on structural MRI images and atlases generated from these images, such as the Automatic Anatomic Labeling template, 9 the Desikan-Killiany atlas, 10 and the Destrieux atlas. 11 While such labels have proven invaluable, they also present some limitations. The distribution of a receptor may not be clearly defined within an anatomical region. Voxelwise analyses can ameliorate this, but without specific hypotheses of voxel location to examine, statistical power may be compromised. Therefore, for some applications, it may be more beneficial to define post-synaptic VOIs based on their receptor distribution, which we refer to as a molecular VOI, as opposed to a structural VOI.
In a healthy state, a higher level of 5-HT1A autoreceptors in the RN results in reduced serotonin output to the rest of the brain, affecting post-synaptic 5-HT1A receptors, which upregulate in a compensatory manner. 12 However, the responsiveness of postsynaptic 5-HT1A to this RN upregulation is unknown. This can be indirectly probed by examining molecular connectivity, the correlation between RN 5-HT1A and 5-HT1A in molecular VOIs. A similar approach has been used in anxiety disorders, in which it was shown that correlation of RN BPF with post-synaptic BPF was higher in patients treated with escitalopram than in untreated patients. 13 This can be interpreted to reflect a restoration of homeostasis, or close coupling between the systems, in the treated anxious brain. It would be valuable to determine if such homeostatic disruptions are present in MDD. Due to the need for sensitive measurements, such an examination is optimally performed in molecular VOIs where there is a known high density of post-synaptic 5-HT1A. If a disruption is found, this could indicate a connectivity deficit in the serotonergic system.
Our previous work has shown differences in raphe 5-HT1A binding between patients with MDD and healthy volunteers. While these differences are significant in both sexes, they are greater in men than in women, such that binding differences cannot be reliably used to separate females with and without MDD, while binding differences in males can distinguish diagnostic groups. 14 Post-mortem studies have also shown sex differences in binding. 15 Therefore, to best capture differences, we focused on men in this study. We hypothesize that 5-HT1A binding in all regions will be higher in MDD than in healthy controls. In addition, we hypothesize that molecular connectivity of the RN with molecular VOIs will be greater in healthy controls than in patients with MDD. As an exploratory measure, we also compare the slope of the linear regression between RN binding potential and molecular VOI binding potential for each group—in this way we can assess not only the goodness of fit (by the molecular connectivity/correlation) of the presynaptic/post-synaptic relationship, but how post-synaptic binding changes with presynaptic binding across participants. Finally, we explore whether differences in molecular connectivity can distinguish men with and without MDD to serve as a biomarker for the disorder, through the use of Euclidean distance from the regression line (DR) between the RN and molecular VOIs. A better understanding of the interplay between pre and post-synaptic serotonin receptors may help guide better treatment and diagnosis.
Materials and methods
Participants
Participant demographicsa.
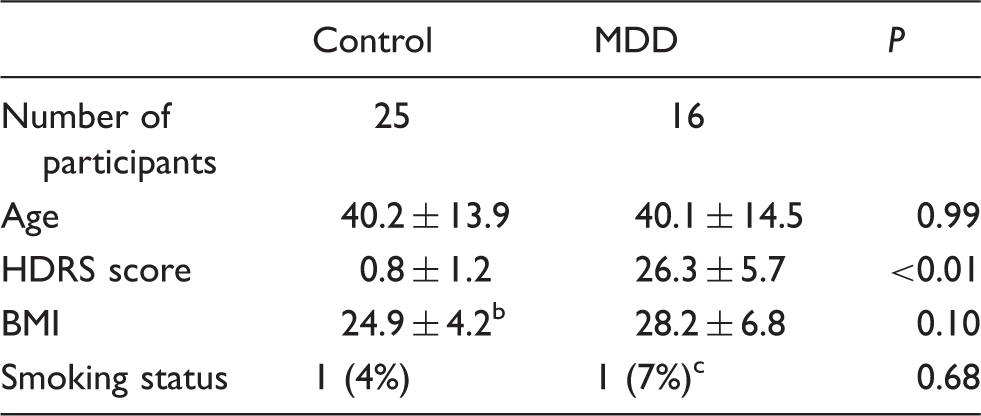
Mean ± SD is reported.
Weight was unknown for one control participant.
Smoking status was unknown for one MDD participant.
MDD: major depressive disorder; BMI: body mass index; HDRS: Hamilton depression rating scale.
Radiochemistry and input function measures
[11C]-WAY100635 preparation, as well as measurement of arterial input function, metabolites, and plasma free fraction (fp) was performed as previously described. 17
Image acquisition and analysis
Magnetic resonance imaging (MRI) and PET data were acquired as previously described. 8 MRI was performed on either a 1.5 T Signa Advantage or a 3 T Signa HDx system (General Electric, Milwaukee, Wisconsin). PET was performed using an ECAT EXACT HR+ (Siemens/CTI, Knoxville, Tennessee); emission data were collected for 110 min as 20 frames of increasing duration. Image analysis was performed using MATLAB (The Mathworks, Natick, Massachusetts), with extensions to functional magnetic resonance imaging of the brain’s (FMRIB’s) linear image registration tool (FLIRT v.5.218), brain extraction tool v1.2, 19 and statistical parametric mapping (SPM5) normalization 20 and segmentation routines, 21 as well as to the advanced normalization tools (ANTs). 22 Frame-by-frame rigid body registration was performed to a reference frame in order to correct for subject motion, and a mean of motion-corrected PET image frames was coregistered to each subject’s MRI, using eight possible coregistration options. 23 Each result was visually inspected and the best coregistration was chosen.
Delineation of molecular VOIs
A separate [11C]-WAY100635 study examined binding in 52 healthy controls, both male and female. 8 Voxel binding maps were calculated using Basis Pursuit and averaged in a standard space. To determine which regions had highest 5-HT1A binding, we used empirically derived BPF thresholds in the cortex. This same thresholding technique was also used within the brainstem to generate the RN VOI. It should be noted the BPF threshold was only used to create the atlas image. The RN and molecular VOIs were then used as an atlas for the current study; these were warped to the individual subject’s PET image using ANTs normalization through the individual’s MRI.
There was a difference in the proportion of controls vs. patients who were scanned on a 1.5 T vs. a 3 T scanner, with 21 controls and 1 patient scanned on a 1.5 T scanner. However, PET-MRI coregistration and VOI placement for each participant were manually checked by a trained technician. Therefore, magnet strength is unlikely to affect molecular VOI placement.
Derivation of outcome measures
Regional distribution volumes (VT) for 11C-WAY100635 were derived in the RN and molecular VOIs as described previously. 8 The closest measure to in vitro binding potential for PET is BPF, or the product of available binding density (Bavail, nM per g of tissue) and affinity (1/KD, nM per ml of brain). We calculate BPF as VT – VND/fP, where VT is the distribution volume in a given region, VND is the distribution volume in a reference region (in this case, the white matter of the cerebellum), and fP is the free fraction of radiotracer in the plasma. We have described kinetic considerations of this measure elsewhere. 17 Briefly, time activity curves (TACs) measured from PET scans were fit to a two tissue compartment model with plasma-to-tissue rate constants constrained to that of the cerebellar white matter, using an arterial input function. Standard error (SE) was calculated using a bootstrapping technique that incorporates errors in plasma, metabolite, and brain data. 24
Region-wise comparison
A weighted linear mixed model was used to compare BPF in molecular VOIs between healthy controls (HC) and patients with MDD. Log-transformation of BPF was applied to meet model assumptions of normality and homoscedasticity. Weights were defined as
Molecular connectivity
To test molecular connectivity between the RN and molecular VOIs, Pearson correlation was calculated between BPF in the RN and BPF in each molecular VOI in all participants. Differences between coefficients in HC and MDD participants were tested using the r.test function in the R package ‘psych’; this function uses a Fisher R to Z transform to determine significant differences between two independent correlation coefficients.
If the linear regression between RN BPF and molecular VOI BPF in controls represents the typical relationship between regions, then deviations from this regression could be calculated as the Euclidean distance of a given participant from the control regression line r, which we denote as DR. In this way, the insight gained by differences in molecular connectivity may also be translated into a clinically useful measure. To this end, we calculated DR of RN BPF versus molecular VOI BPF. As shown in Figure 1, we calculated the vertical (y′) and horizontal (x′) distance from the regression line given the individual participant’s x and y values, where x = RN BPF and y = BPF of a molecular VOI. Given that y′, x′, and the distance between them, h, form a right triangle, we can calculate the angle θ
Schematic on calculation of DR (Euclidean distance of point p from regression line r). If y′ is the vertical distance from r (the regression line for healthy controls) and x′ is the horizontal distance from r, then x′, y′, and h form a right triangle. The angle θ can thus be calculated as tan−1(x′/y′). Using the angle θ, we can then calculate the distance from r, DR, as the altitude of this triangle using the equation sin θ = DR/y′, or DR = y′ sin θ.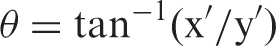
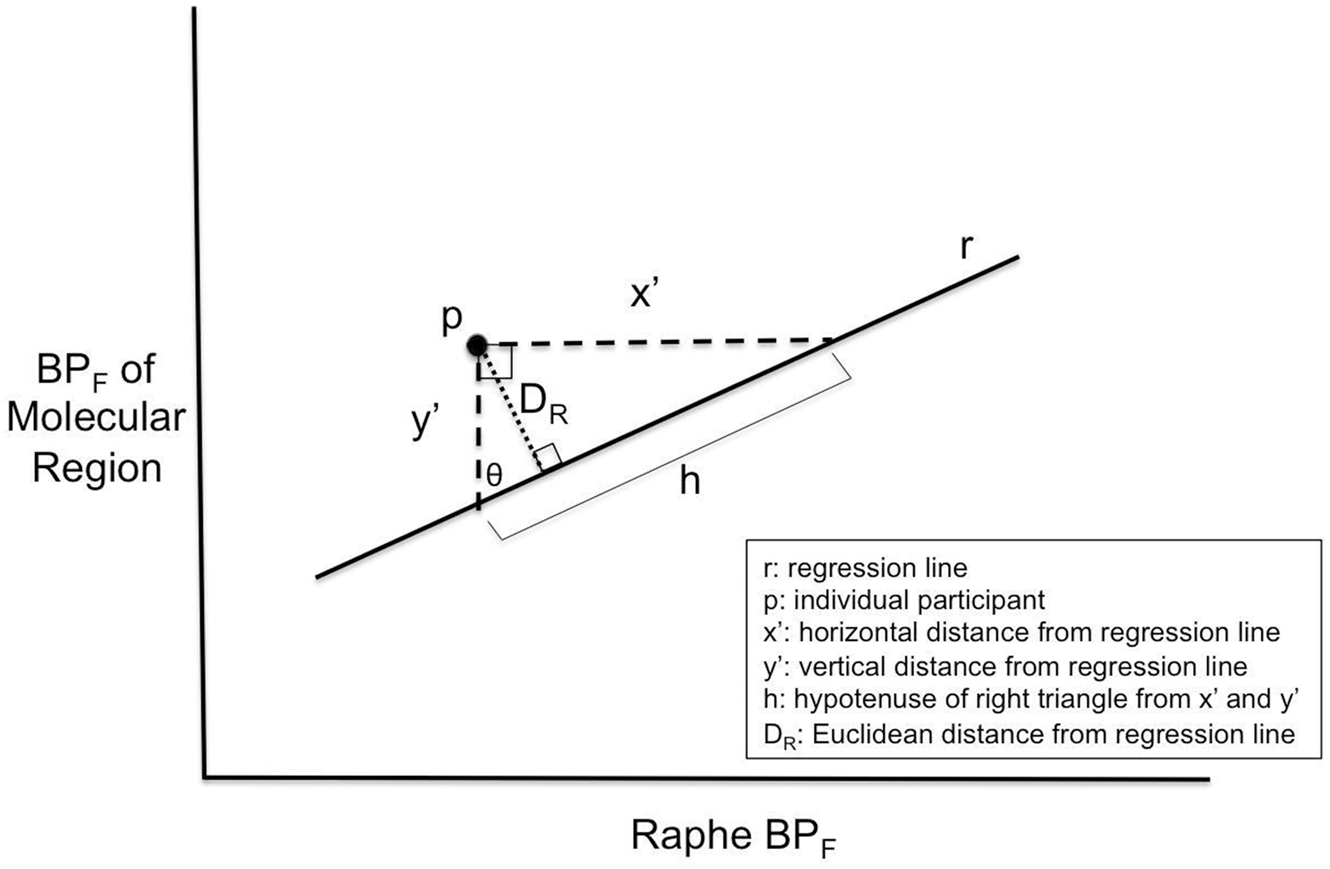
Using this value of θ, we can construct a new right triangle with the distance from the point p to the regression line (DR) as the altitude. Using the formula
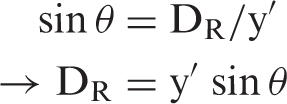
We did not explore age differences in the above analyses, as we have not found there to be a relationship between age and binding potential. 27
Results
Molecular VOIs
Through our thresholding technique, we were able to identify for the first time two PET-derived regions that did not overlap with discrete anatomical regions. The first of these was a small region lying at the posterior aspect of the olfactory sulcus (OLFS). The second was a broad strip of medial temporal cortex that encompassed piriform, olfactory, and entorhinal cortex (referred to as piriform cortex or PRF). The volumes of these regions were 440 mm3 for left OLFS, 520 mm3 for right OLFS, 4072 mm3 for left PRF, and 3624 mm3 for right PRF. Images of these regions can be found in Figure 2. To determine whether using these regions provided an advantage over traditional atlas-based VOIs, we compared binding in each molecular VOI to its closest structural atlas-based VOI (left and right anterior cingulate for left and right OLFS, respectively, and left and right hippocampus for left and right PRF, respectively) using t-tests. Results indicated that both left and right PRF had significantly higher BPF than left and right hippocampus (p < 0.01, Bonferroni corrected), but higher binding in the left and right OLFS compared to left and right anterior cingulate was nonsignificant (p = 0.08 and 0.32 for left and right respectively, Bonferroni corrected, Figure 2).
Molecular VOIs labeled in sagittal (a), coronal (b), and oblique views (c) as a 3D image projection of an averaged [11C]-WAY100635 image. OLFS: olfactory sulcus; PRF: piriform cortex. (d) Molecular VOIs show greater binding potential than nearby structurally defined regions. ACN: anterior cingulate; HIP: hippocampus.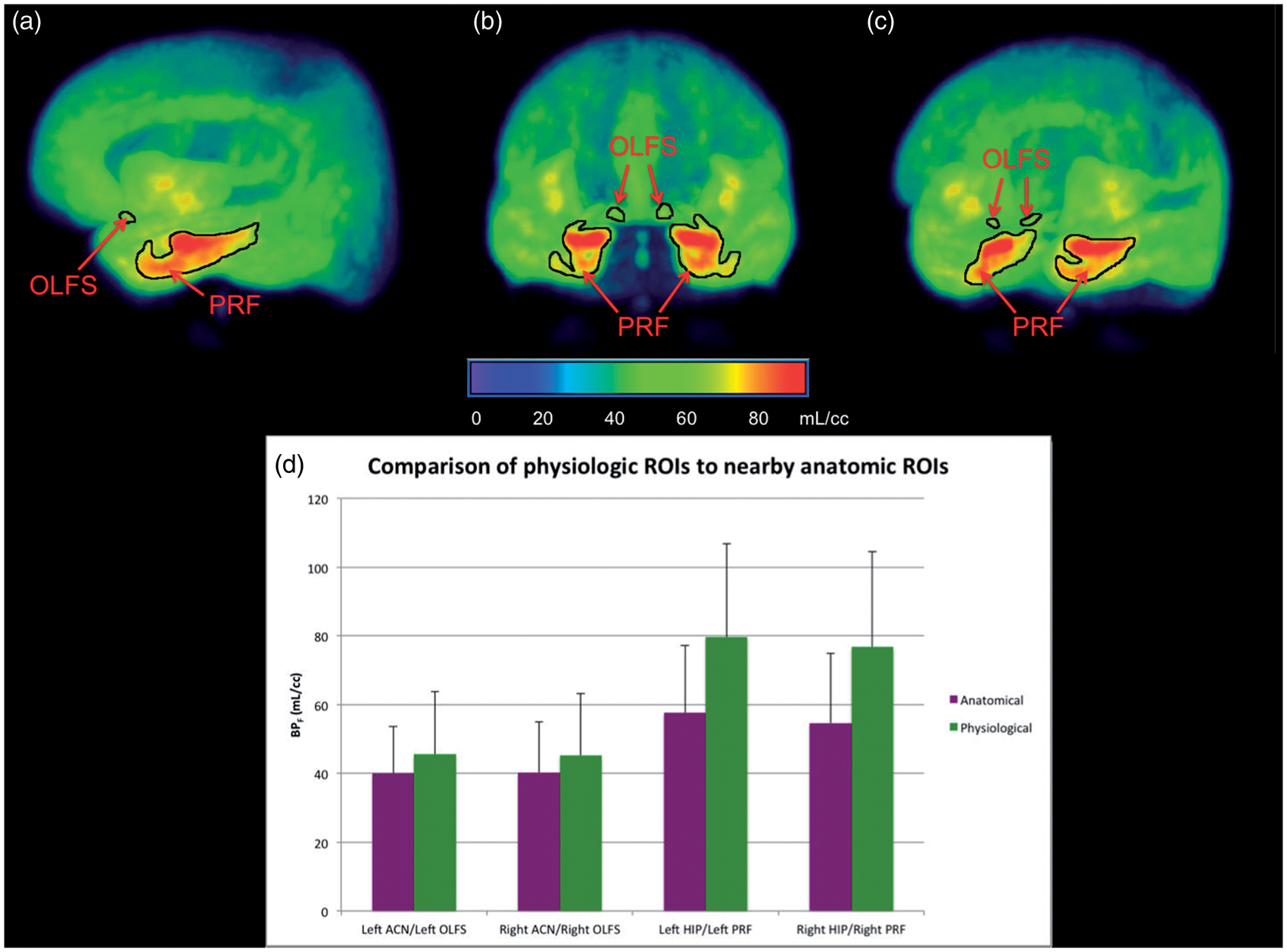
BPF comparison in OLFS and PRF
In both molecular VOIs, OLFS and PRF, bilaterally, males with MDD had higher BPF than did healthy controls (p < 0.01, Bonferroni corrected, see Figure 3). Average BPF in subjects with MDD was 82% greater in left OLFS, 54% greater in right OLFS, 67% greater in left PRF, and 67% greater in right PRF, than the average BPF of healthy controls.
In males, patients with major depressive disorder (MDD) show consistently greater 5-HT1A binding in the raphe and molecular VOIs compared to controls. Error bars represent standard error. *p < 0.01, Bonferroni corrected.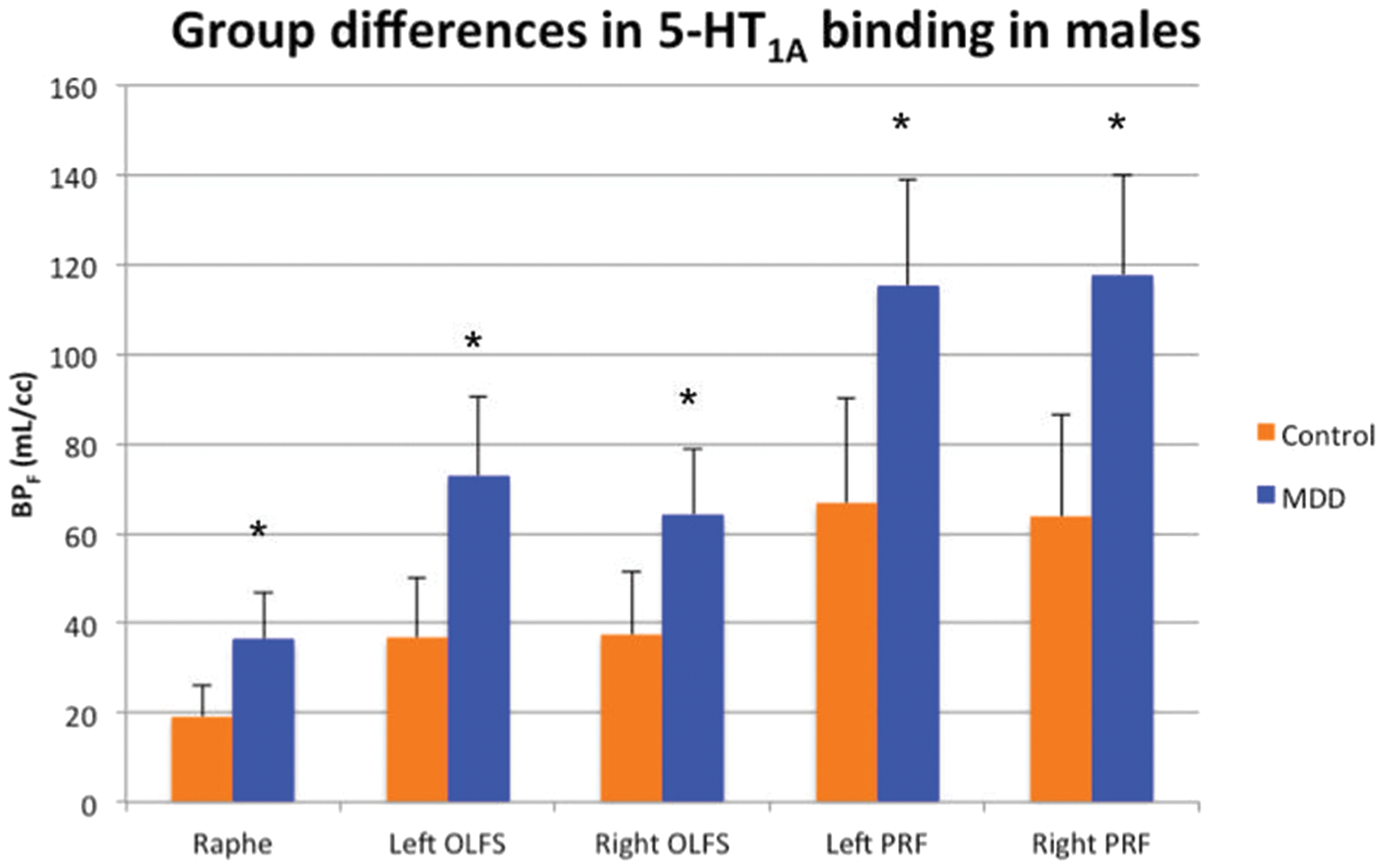
Molecular connectivity
Molecular connectivity.
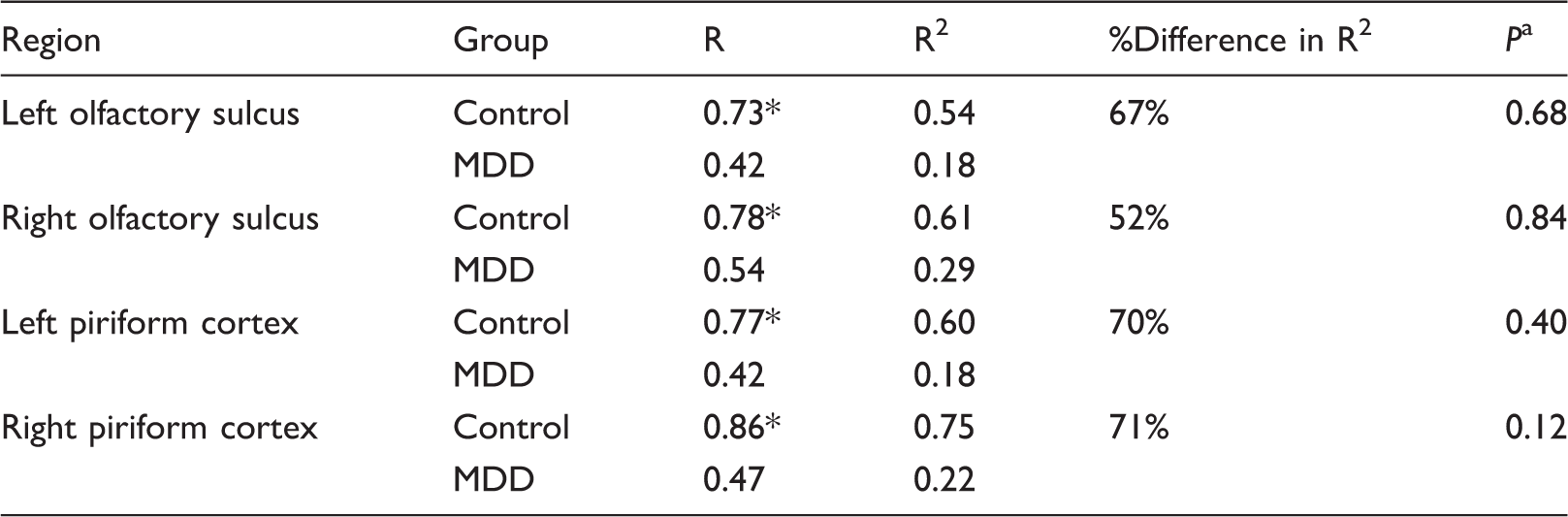
Bonferroni corrected, determined by Fisher R to Z transform.
P < 0.05, Bonferroni corrected.
MDD: major depressive disorder.
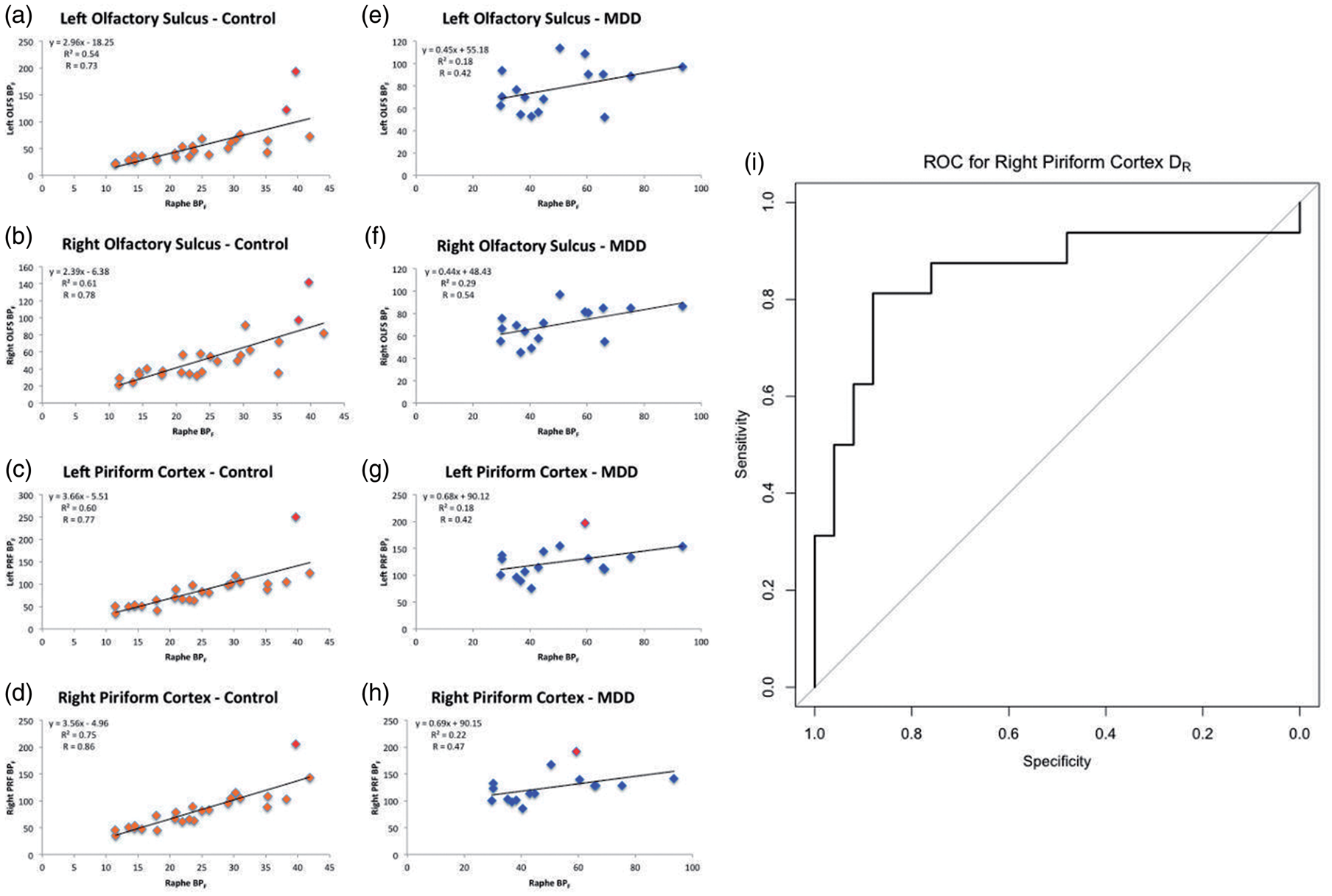
(a–h) Molecular connectivity (correlations) between raphe nucleus BPF and BPF in left OLFS (a,e), right OLFS (b,f), left PRF (c,g), and right PRF (d,h) in males. Outliers (defined as outside the interval [75% quantile ± 1.5*interquartile range] in either the raphe nucleus or in molecular ROIs) are marked in red. (i) Receiver operating characteristic (ROC) curve for the use of distance from the regression line (DR), using the linear regression between raphe binding potential (BPF) and right piriform cortex (PRF) BPF, as a diagnostic test in major depressive disorder.
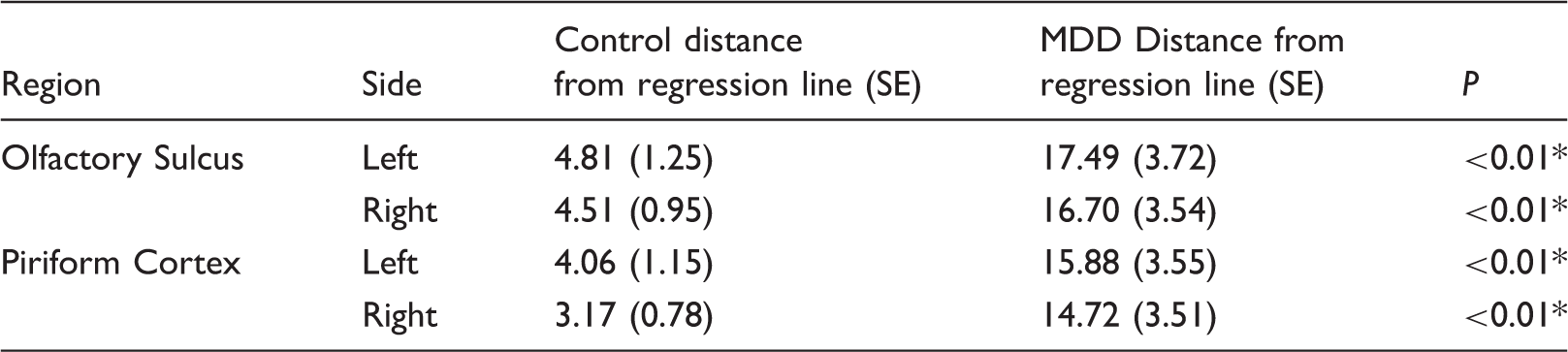
Euclidean distance from regression line (DR)
Distance from regression line (DR).
P < 0.05.
SE: standard error, defined as (standard deviation ÷ √(n)); MDD: major depressive disorder.
Discussion
As the RN is the central output of serotonin to the remainder of the central nervous system, we would expect pre and post-synaptic 5-HT1A binding to be correlated in a healthy state. However, the relationship between these two entities in MDD has been unknown. Using PET-derived measures, we were able to assess molecular connectivity, i.e. the relationship between pre and post-synaptic 5-HT1A receptors in molecularly defined regions for the first time. Importantly, we find this connectivity to be disrupted in MDD in men; RN autoreceptor binding potential (BPF) is not significantly correlated with that in molecular VOIs, as it is in controls. As BPF is significantly higher in both the RN and molecular VOIs in MDD, such a relationship may not have been observable through traditional analyses. The significance of this finding is that reduction of molecular connectivity in MDD suggests a disruption in serotonin transmission and homeostasis.
It is likely that both molecular VOIs are connected to the RN via the medial forebrain bundle, and that while the OLFS is innervated by projections from the median RN, PRF is innervated by projections from the dorsal RN. 28 That the OLFS lies directly between the subgenual ACC and ventromedial PFC, both heavily implicated in MDD,29,30 further supports its involvement in depressive symptomology. Both molecular VOIs are largely implicated in olfactory function, a sensory modality that has not been well studied in MDD. However, removal of the olfactory bulb in rats is used as a model for MDD.31,32 In addition, clinical studies have shown differences in olfactory sensitivity in MDD—two studies showed a decrease,33,34 while one showed an increase. 35 A recent MRI study showed lower OLFS sulcal depth in MDD. 36 Using these molecularly defined VOIs could allow greater exploration of the role olfaction plays in MDD, and whether these effects pertain to sensation in general.
We found that 5-HT1A binding in the RN was significantly correlated with 5-HT1A binding in molecular VOIs in healthy controls, but not in the MDD group, which suggests dysregulation. This has implications for future MDD monitoring and treatment and further suggests that examining either pre or post-synaptic receptors in isolation (as is most often performed) may not yield a complete picture. Although a direct comparison of molecular connectivity across groups did not reach statistical significance, there are several reasons to investigate this further. (1) In an analysis with outliers (marked in red in Figure 4) removed, molecular connectivity differences were accentuated, with bilateral PRF showing significantly greater connectivity in control males than males with MDD (p = 0.02 and 0.01 for left and right PRF, respectively, Bonferroni corrected). In future analyses with larger cohorts, such outliers may not significantly affect overall connectivity findings. (2) Molecular connectivity differences in right PRF were significant before multiple comparisons correction, indicating that this region would be a good a priori choice for future studies. This region in particular may be valuable to study for several reasons. (a) It is large and therefore not as susceptible to partial volume effects 37 as smaller regions may be. (b) As a composite region including olfactory, piriform, and entorhinal cortex, it may have far reaching neural effects; entorhinal cortex in particular is known to have widespread afferent fibers from the cortex 38 and is critical in sending efferent fibers to the hippocampus via the perforant pathway. 39 (3) Molecular connectivity slopes were significantly different between healthy controls and patients with MDD, indicating an altered relationship between pre and post-synaptic receptors in molecular VOIs. It is important to note that there need not be a difference in correlation (molecular connectivity) for this to be true—two groups can have an equal goodness of fit but have differing linear relationships. Moreover, correlations may be more sensitive to noise and variability than regression slopes—therefore, a difference in slopes between diagnostic groups strongly suggests that there is a real difference in the relationship between pre and post-synaptic receptors in MDD.
It is plausible that molecular connectivity differences are the result of differences in structural connectivity of serotonergic neurons between the RN and molecular VOIs. Unfortunately, there is relatively little agreement in the literature with respect to diffusion tensor imaging (DTI) studies in MDD—many regions of the brain show differences in structural connectivity in MDD, but these regions are not consistently implicated. The most common of these regions are areas of frontal and temporal cortex,40,41 which partially validate our current findings, as our molecular VOIs lie in these regions. In a large multi-site study, our group found no differences in structural connectivity between depressed patients and healthy controls, though fractional anisotropy of a strip of right medial orbitofrontal cortex correlated negatively with depression severity. 42 This region lies directly anterior to, and may overlap with, the right OLFS. However, a previous study from our lab showed that 5-HT1A and serotonin transporter levels do not correlate. If transporter levels are taken as a proxy of axonal density, one potential implication of this is that autoreceptor density and innervation density might not share a direct relationship. 43 A DTI study involving these regions in a larger sample would clarify the relationship between molecular and structural connectivity—we have performed similar DTI studies between the RN and anatomical regions.44,45 In addition, these regions could be assessed with functional connectivity; while molecular VOIs have not been examined in this context, there have been studies examining functional connectivity of the RN in healthy controls.46,47
A second possibility is that reduced molecular connectivity is due to a disproportionately higher BPF in the RN with respect to molecular VOIs in MDD, which may be dependent on receptor upregulation alone, independent of structural connectivity or innervation density. This explanation is more consistent with our finding of a lack of association between 5-HT1A and serotonin transporter. Based on preclinical studies, we hypothesize that post-synaptic changes occur as a result of presynaptic changes. In this case, in healthy controls, regression slopes indicate that a difference in 5-HT1A autoreceptors between participants would correspond to a two to threefold difference in post-synaptic receptors. This was the opposite in MDD; in these patients, a difference in autoreceptors would only correspond to around half of that difference post-synaptically. Even though all regions examined had greater binding potential in subjects with MDD than in controls, these differences in molecular connectivity slope indicate that BPF differences are not uniform; compensatory 5-HT1A upregulation in MDD may not be proportional to increases in 5-HT1A autoreceptor density.
As molecular connectivity is not correlated with depression severity, it may not be a causative mechanism of MDD. However, it may still serve as a biomarker—to that end, we explored the possibility of using molecular connectivity for diagnostic purposes through the use of Euclidean distance from the presynaptic/postsynaptic 5-HT1A regression in controls, DR. This measure can give an indication of how much an individual deviates from the typical presynaptic/postsynaptic receptor relationship. Males with MDD had much higher DR values than controls in all regions, suggesting that DR may serve as a biomarker. Our lab has previously explored the potential for a biomarker in the RN; 14 in males, RN 5-HT1A thresholds showed high specificity and sensitivity for diagnosing MDD. Our current results indicate both left and right PRF show 81% sensitivity and 88% specificity. This is a slight increase from our previous study. Therefore, a biomarker that examines both pre and post-synaptic receptors may be more efficacious, and provide more information about serotonergic function, than one that only examines autoreceptor binding. The high sensitivity and specificity of such a biomarker exceed the diagnostic accuracy of current clinical measures, 48 further supporting its use—using DR, we may be able to study subtypes of MDD, evaluate targeted therapies, and identify people at increased risk. 49 Should molecular connectivity show a relationship with DTI, it can also open the door for less invasive and costly surrogates.
There are limitations to this study. Our sample consists of 25 healthy controls and 16 patients with MDD; our findings should be replicated in a larger sample. There is also a possibility that the smaller sample size of the MDD group relative to the control group contributes to the lack of significance in molecular connectivity, though it should be noted that the correlation coefficient would not be expected to change given the variance in this group. In addition, it would be useful to examine sex differences in molecular connectivity. Given evidence that sex hormones may play a role in 5-HT1A expression and action,50,51 future investigations in molecular connectivity should record and covary for differences in menstrual cycle at the time of PET scans.
An important caveat to our study of WAY100635 is that this receptor also binds to the dopamine D4 receptor, though at lower affinities than 5-HT1A. 52 As D4 has been found in areas such as the entorhinal cortex,53,54 which comprises part of our PRF region, this may have an effect on PRF binding potential. The close correspondence of binding in controls between PRF and RN, which does not have significant D4 binding, 54 gives us confidence that our findings are based on the serotonin system. However, further study of the dopamine D4 receptor in MDD may provide insight that the serotonin system alone cannot elucidate.
Conclusions
We have identified, for the first time, two molecular VOIs, defined by serotonin 1A (5-HT1A) receptor distribution, that show significantly higher binding in MDD. In studying the association between presynaptic and post-synaptic 5-HT1A in the RN and these molecular VOIs in males, which we term molecular connectivity, we found that while RN autoreceptors are tightly correlated with post-synaptic receptors in healthy controls, they are nonsignificantly correlated in MDD. Further, on average, healthy controls had two to three times greater molecular connectivity slopes than patients with MDD. Examining PET-derived data in this way suggests that structural and compensatory mechanisms of post-synaptic receptor regulation are compromised in MDD. Individual differences in molecular connectivity are so large that they may even serve as a biomarker for the disorder. Finally, in applying molecular connectivity to other disorders, we may be able to identify patterns of disruption that we would not be able to examine through analysis of single regions.
Footnotes
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The work was supported by the following grants: F30MH109412 (PI: Pillai), R01MH40695 (PI: Mann), P50MH062185 (PI: Mann), R01MH074813 (PI: Parsey), R01MH090276 (PI: Parsey), and K01MH091354 (PI: DeLorenzo) awarded by the National Institutes of Health, a Clinical and Translational Science Award (CTSA) from Columbia University, and grants from the American Foundation for the Prevention of Suicide (AFSP) and the National Alliance for Research on Schizophrenia and Depression (NARSAD).
Acknowledgments
We acknowledge the biostatistical support provided by the Biostatistical Consulting Core at School of Medicine, Stony Brook University, and the anatomical support by Dr. Mary F. Kritzer and Dr. Efrain Azmitia in helping us identify molecular VOIs. We also acknowledge the support in patient recruitment carried out by Dr. Greg Sullivan.
Declaration of conflicting interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Oquendo receives royalties for the use of the Columbia Suicide Severity Rating Scale. Her family owns stock in Bristol Myers Squibb. Mann receives royalties for commercial use of the C- SSRS from the Research Foundation for Mental Hygiene and has stock options in Qualitas Health, a startup company making omega-3-fatty-acid products. Pillai, Zhang, Yang, Parsey, and DeLorenzo report no biomedical financial interests or potential conflict of interest.
Authors’ contributions
Rajapillai Pillai processed and analyzed the data and wrote the manuscript. Mengru Zhang and Jie Yang performed statistical analysis and wrote relevant sections of the methods. J. John Mann, Maria A. Oquendo, and Ramin V. Parsey acquired PET data. Christine DeLorenzo oversaw this project and provided feedback on writing and analysis. All authors read and approved of this manuscript.