
Abstract
We carried out a systematic review of modern-era (1990–2025) placebo-controlled studies assessing the acute and post-acute effects of lysergic acid diethylamide, dimethyltryptamine and psilocybin on cognitive and psychological functions. From February 28 to March 19, 2025, PubMed and APA PsychINFO were systematically searched for placebo-controlled studies examining the influence of psychedelics on empathy, reaction time, attention, inhibition, emotional processing, memory, cognitive flexibility, and related cognitive functions using experimental methods. Additional searches were done in Google Scholar. The systematic review included 32 studies. Psychedelics tended to enhance emotional empathy but had no effect on cognitive empathy. Psychedelics impaired, enhanced or had no effect on memory depending on the task and timing of the assessment. Dose-dependent impairments were seen in many of the reaction time, attention, and inhibition tasks, although some studies found no effects. Some studies found impaired recognition of negative stimuli under the acute effects of psychedelics. The findings regarding cognitive flexibility were mixed. Many studies had small samples, and it is hard to find a reliable placebo due to psychedelics’ unique subjective effects. Future studies should use bigger samples and also study more longitudinal effects of psychedelics on cognitive and psychological functions.
Introduction
Psychedelics have recaptured the attention of scientists as recent research suggests that they may be suitable for therapeutic use in the treatment of anxiety, depression, post-traumatic stress disorder, alcoholism and tobacco addiction (Belouin and Henningfield, 2018; Nutt, 2019; Nutt et al., 2020; Vargas et al., 2020). Classic psychedelics include psilocybin (i.e., magic mushrooms), LSD (lysergic acid diethylamide), DMT (dimethyltryptamine) and mescaline (found in several cacti species) (Andersen et al., 2020; Nutt et al., 2020; Nichols, 2016). These substances produce altered states of consciousness (also known as psychedelic experience) that are characterized by geometric patterning (Bressloff et al., 2002), ego dissolution (Nour et al., 2016), changes in executive functions (Barrett et al., 2018; Basedow et al., 2024; Pokorny et al., 2020), increased brain entropy (Carhart-Harris et al., 2014) and mystical experiences (Griffiths et al., 2006), although the exact mechanisms that underlie these phenomena still remain elusive (Girn, 2023) as several models have been proposed for explaining these effects (Kwan et al., 2022). The effects of classic psychedelics are primarily driven by activation of the 5-HT2A receptor, as their impact is significantly diminished when 5-HT2A antagonists, such as ketanserin, are administered (Preller et al., 2017).
Classic psychedelics have been used in healing settings since ancient times (Guerra-Doce, 2015), although they gained more public attention after 1938, when Albert Hofmann first synthesized LSD, sparking interest due to its potential in treating mental health disorders (Hofmann, 2013). The first studies on the therapeutic effects of psychedelics looked at the effects of LSD-assisted psychotherapy (Abramson, 1967; Busch and Johnson, 1950; Chandler and Hartman, 1960). After that, other classic psychedelics, for example, psilocybin, also gathered research and public attention (Hofmann, 2013). Along with their therapeutic potential, the effects of psychedelics on cognitive and psychological functioning have also been investigated. Multiple studies demonstrated that psychedelics prolonged reaction times to auditory stimuli (Abramson et al., 1955; Dykstra and Appel, 1972), altered time perception (with effects varying across studies) (Deshon et al., 1952; Malitz et al., 1960), disrupted body schema (Liebert et al., 1957) and induced visual distortions (Fischer et al., 1970) (see Aday et al., 2021 for a review).
Most of the studies had many limitations, such as small sample sizes and unrefined methodological standards, including no control groups and a lack of detailed reporting of methodology (Aday et al., 2021). The research stopped in the 1970s as many classic psychedelics were associated with social unrest and anti-war protests related to the anti-Vietnam War movement (Wesson, 2011). There were also growing concerns about the potential dangers of psychedelic use, including psychosis and unpredictable behavior. As a result, research on these substances was largely limited until the early 2000s (Griffiths et al., 2006). In the 21st century, early research in Europe investigated the immediate psychological and physiological impacts of these substances (Griffiths et al., 2006; Vollenweider et al., 1998). Studies from this century have better methodological standards compared to the previous century studies, although they still have small sample sizes (Barrett et al., 2018; Carter et al., 2005), and the placebo effect is a common problem in psychedelic research (Olson et al., 2020).
Most of the recent psychedelic research has focused on neuroimaging studies (Barrett et al., 2020) and the therapeutic potential of psychedelics due to the current mental health crisis (Nutt et al., 2020). However, cognitive and psychological functions such as memory, empathy, emotional processing, cognitive flexibility and reaction time remain less thoroughly studied in relation to the effects of psychedelics (Bălăeţ, 2022). These processes are typically measured during the acute effects or post-drug administration, using a variety of cognitive and psychological tests (Basedow et al., 2024). Although measuring these processes is crucial for understanding psychedelics’ impact, there are also significant limitations in how these assessments reflect the psychedelic experience. The aim of this review is to systematically explore these processes and their measurement in the context of psychedelic research. This review extends previous work (Basedow et al., 2024; Cardoso and de Barros Viana, 2023; Meshkat et al., 2024; Pouyan et al., 2022, 2023; Ramos et al., 2024; Velit-Salazar et al., 2024) by specifically examining the effects of classic psychedelics on cognitive and psychological functions using experimental methods in placebo-controlled studies.
Method
Data for this systematic review were collected following the systematic reviews and meta-analyses guidelines (Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA); Page et al., 2021). One of the authors performed searches, screening and data extraction according to the PRISMA standards.
Search
PubMed and APA PsycInfo were used to find articles that were published from January 1990 to March 2025. A search of Google Scholar was conducted to identify any additional relevant articles. The last date of the search was 18.03.2025. Reference lists of included studies were checked to identify additional studies. The following search terms were used: psychedelic* OR hallucinogen* OR psiloc* OR “shrooms” OR LSD OR ayahuasca OR DMT AND “reaction time” OR “response time” OR “cognitive flexibility” OR “task switching” OR “memory” OR “working memory” OR “short-term memory” OR “visuospatial memory” OR “autobiographical memory” OR “episodic memory” OR “semantic memory” OR “emotional processing” OR “emotional face recognition” OR “empathy” OR “emotional empathy” OR “cognitive empathy” OR cognition OR attention OR inhibition OR “inhibitory control” OR “response inhibition.” The literature search targeted the title and the abstract.
Eligibility criteria
Placebo-controlled trials in clinical and normative populations were included. All age groups were included. The studies investigated acute and/or long-term effects of classic psychedelics on cognitive and psychological functions using behavioral or physiological measures. There were no limits to the timing of cognitive assessments relative to psychedelic administration. Exclusion criteria included studies that: (1) were not written in English, (2) were not available through institutional access, (3) were not peer-reviewed, (4) did not include placebo, (5) animal studies, (6) used self-reported methods (not experimental tests), (7) did not report inferential statistical outcomes (p-values or effect sizes).
Data extraction
After removing duplicates, all remaining articles underwent title and abstract screening. Studies that did not fulfill the inclusion criteria were excluded. Full-text assessments were conducted to determine eligibility. For each included study, the following data were extracted: (1) publication year and author names; (2) participant demographics; (3) cognitive and/or psychological assessment methods; (4) the specific psychedelic compound and administered dose; and (5) key findings along with measurement time points. The studies were grouped narratively by cognitive/emotional domain.
Study quality
Risk of bias was assessed using the Cochrane Risk of Bias 2.0 (RoB 2.0) tool for both parallel-group and crossover randomized controlled trials (Sterne et al., 2019). Two reviewers independently evaluated each study across all RoB 2.0 domains, with disagreements resolved through discussion. Detailed domain-level assessments for each study are provided in Appendix S1 (parallel-group trials) and Appendix S2 (crossover trials).
Identified studies
A flow diagram (Page et al., 2021) illustrating the different phases of the systematic review is presented in Figure 1.
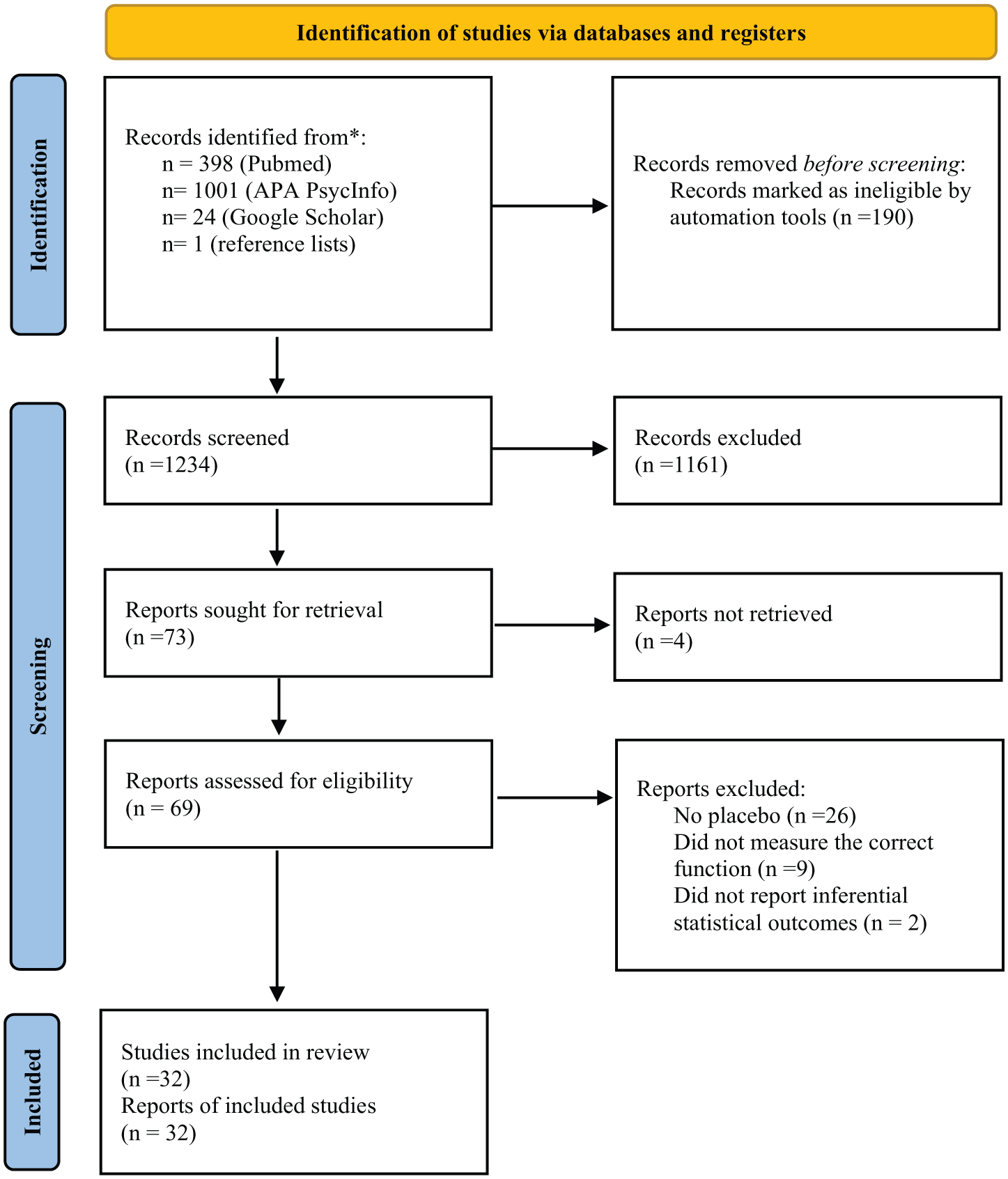
PRISMA flow diagram for study selection.
Study selection
One thousand two hundred thirty-four references were screened based on titles and abstracts and 1161 records were excluded during screening, leaving 73 potential research papers (see Figure 1). Full-text reports of the 69 identified research papers were assessed according to eligibility criteria. The systematic search identified 32 eligible studies in total—14 for reaction time, attention and inhibition, 5 for cognitive flexibility, 14 for memory, 8 for emotional processing and 6 for empathy. Because several studies assessed more than one cognitive domain, individual studies were counted in multiple categories. Thirty-one studies were done in healthy volunteers, and 1 study was done in people with depression.
Results
Psychedelics’ effects on reaction time, attention and inhibition
Reaction time (RT) is measured by the elapsed time between stimulus onset and an individual’s response on cognitive tasks. Higher levels of attention result in a shorter RT and vice versa. Inhibition further modulates cognitive control by enabling the suppression of irrelevant stimuli and the regulation of appropriate responses.
The test for measuring RT in the analyzed experimental studies were the Psychomotor Vigilance Test (PVT; Dinges and Powell, 1985), the Digit Symbol Substitution Test (DSST; McLeod et al., 1982), the motor praxis (mpraxis) task (Gur et al., 2010), the Frankfurt Attention Inventory (FAIR; Moosbrugger and Oehlschlägel, 1996), a multiple-object tracking task (Pylyshyn and Storm, 1988), the Cambridge Neuropsychological Test Automated Battery (CANTAB) reaction time (RTI) and rapid visual information processing (RVP) test (Cambridge Cognition, 2016), the Attentional Blink task (Shapiro et al., 1997), the Stroop Test (Stroop, 1935), the Trail Making Test (TMT; Reitan and Wolfson, 1995), the AX continuous performance test (AX-CPT; Rosvold et al., 1956) and the Go/No-Go task (Nosek and Banaji, 2001) (see Table 1). Studies examining the effects of psychedelics on RT employed a range of dosages across various substances. Research with LSD used microdoses of 5, 10 and 20 μg (Hutten et al., 2020), 5, 10, 20 μg (Family et al., 2020), 6.5, 13 or 26 μg of LSD (Bershad et al., 2019) and 13 or 26 μg (de Wit et al., 2022). Medium doses of LSD were also used (50 μg (Wießner et al., 2022) and 100 μg (Schmidt et al., 2018). Psilocybin studies tested low to high doses of the drug—115, 215 and 315 μg/kg (Vollenweider et al., 2007), 10, 20 and 30 mg/70 kg (Barrett et al., 2018) and 45, 115, 215, 315 μg/kg of psilocybin (Hasler et al., 2004). Other studies used medium doses—215 μg/kg of psilocybin (Carter et al., 2005), 0.28 mg/kg of psilocybin (Umbricht et al., 2003) and 0.5 g of dried mushrooms, which is representative of the upper range used for microdosing (Cavanna et al., 2022). A high dose of psilocybin (260 μg/kg) was also used by Quednow et al. (2012). Fifteen milligram of psilocybin (medium dose) was used by Mallaroni et al. (2023).
Summary of studies investigating the effects of psychedelics on reaction time, attention and inhibitory control.
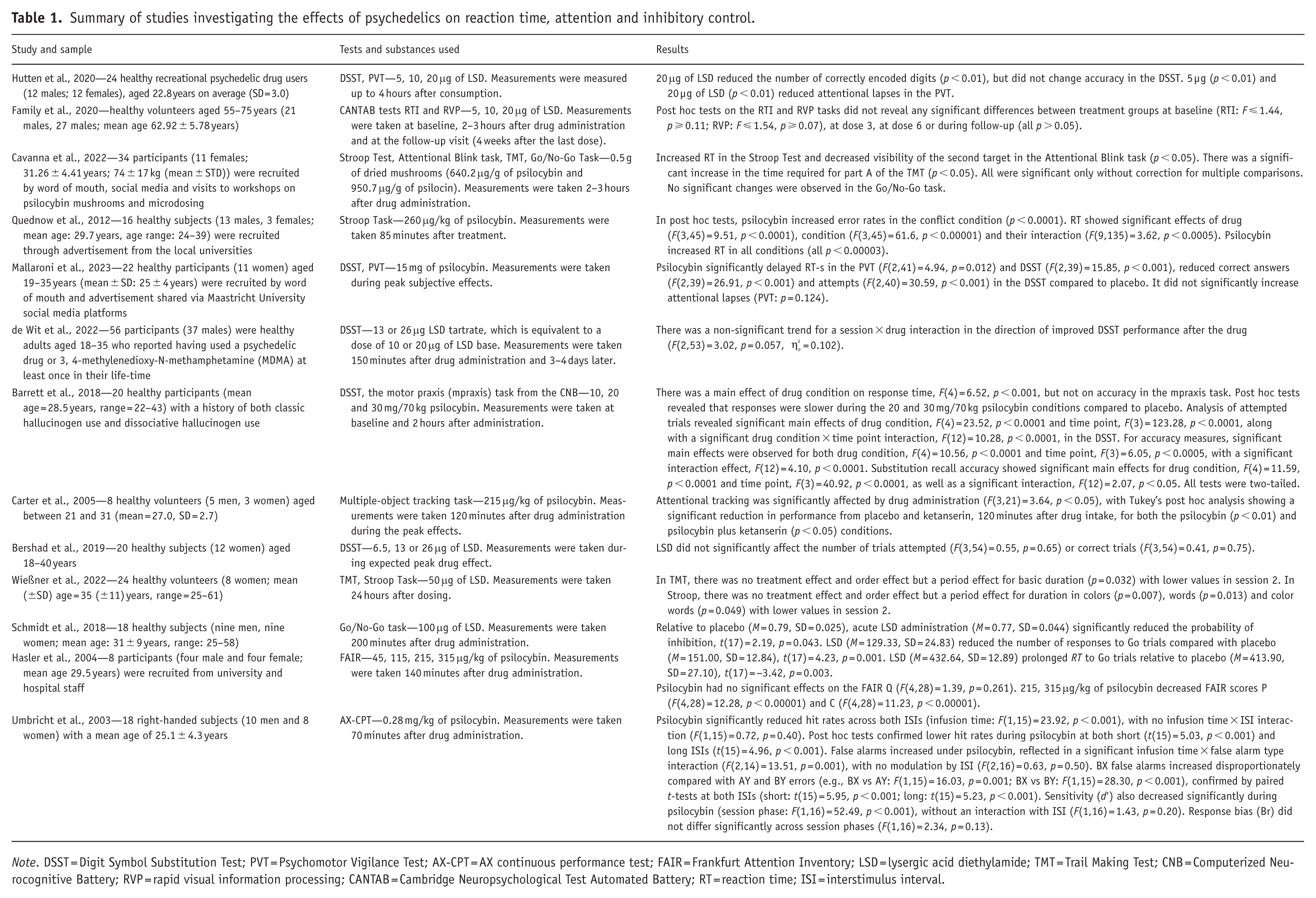
Note. DSST = Digit Symbol Substitution Test; PVT = Psychomotor Vigilance Test; AX-CPT = AX continuous performance test; FAIR = Frankfurt Attention Inventory; LSD = lysergic acid diethylamide; TMT = Trail Making Test; CNB = Computerized Neurocognitive Battery; RVP = rapid visual information processing; CANTAB = Cambridge Neuropsychological Test Automated Battery; RT = reaction time; ISI = interstimulus interval.
Reaction time
A moderate dose of psilocybin (15 mg) was found to delay RT in both the DSST and the PVT (Mallaroni et al., 2023). Similarly, psilocybin (20 and 30 mg/70 kg) significantly slowed RT in the mpraxis task, which assesses psychomotor speed (Barrett et al., 2018). However, accuracy in the mpraxis task remained unaffected.
In contrast, studies with low doses of LSD (5, 10 and 20 μg) reported no significant changes in RT in the RTI test (Family et al., 2020). Similarly, Wießner et al. (2022) found that 50 μg of LSD did not significantly alter Stroop Test performance or RT when measured 24 hours post-administration. A microdosing study using 0.5 g of dried mushrooms (containing approximately 0.32 mg of psilocybin at 640.2 μg/g) did not significantly increase RT in the Stroop Test (Cavanna et al., 2022). There was a significant increase in the time required for part A of the TMT, that measures attention (sustained and shifting) and coordination, where the subject must connect numbers 1–25 in numerical order. This result was not significant after correction for multiple comparisons.
Attention
Sustained attention, as measured by the PVT, improved under microdoses of LSD. Specifically, both 5 and 20 µg (but not 10 µg) reduced attentional lapses, indicating enhanced vigilance for up to 4 hours after ingestion (Hutten et al., 2020). Twenty microgram of LSD was found to reduce the number of correctly encoded digits measured up to 4 hours after consumption, but did not change accuracy in the DSST. In contrast, 15 mg of psilocybin did not significantly affect attentional lapses in the PVT (Mallaroni et al., 2023).
In studies using the DSST, low doses of LSD (6.5–26 μg) did not significantly influence performance (Bershad et al., 2019; de Wit et al., 2022). However, higher doses of psilocybin (20–30 mg/70 kg) significantly reduced the number of attempted trials, accuracy and recall (only for 30 mg) in the DSST in participants with a history of classic psychedelic use (Barrett et al., 2018).
A total of 0.5 g of dried mushrooms caused a decrease in the visibility of the second target in the Attentional Blink task (Cavanna et al., 2022). However, this was not significant after correction for multiple comparisons. All of the measures were taken 2–3 hours after drug administration. Conversely, Wießner et al. (2022) found no significant changes in either part of the TMT following 50 μg of LSD, which could be due to the 24-hour delay in assessment.
Sustained attention has also been assessed using the FAIR, where psilocybin was shown to impair performance in a dose-dependent manner at doses of 115, 215 and 315 μg/kg (Vollenweider et al., 2007). Specifically, psilocybin significantly reduced the FAIR attentional performance capacity score P and the attentional continuity score C across all doses, while the FAIR score Q—reflecting the proportion of attentively made decisions relative to the total number of decisions—was significantly reduced only at the highest dose. In another study, psilocybin (45, 115, 215, 315 μg/kg) had no significant effects on the FAIR score Q (Hasler et al., 2004). However, 215 and 315 μg/kg of psilocybin decreased the FAIR scores P and C. Further support for psilocybin’s impairing effects on task-based attention comes from a multiple-object tracking task, where a dose of 215 μg/kg significantly impaired participants’ ability to track four targets, both when administered alone and following pretreatment with the 5-HT2A antagonist ketanserin (Carter et al., 2005). This impairment was observed 120 minutes post-administration. The lack of attenuation by ketanserin suggests that psilocybin’s impact on attentional tracking may not be primarily mediated by 5-HT2A receptor mechanisms. Although participants reported that they could still comprehend the task instructions, they generally found the attention task substantially more challenging under the influence of psilocybin.
Further evidence for psilocybin’s impact on attention comes from a study using the AX-CPT (Umbricht et al., 2003). Administration of 0.28 mg/kg psilocybin significantly reduced hit rates (correct detection of AX sequences) at both short and long interstimulus intervals (ISIs), with no differential effect across ISIs. Psilocybin also led to a significant decrease in sensitivity, again independent of ISI, while leaving response bias unchanged. In addition, psilocybin increased false alarms, with the most pronounced rise occurring for BX trials—those requiring inhibition of a prepotent response and reliance on contextual information. Taken together, the findings from this and the previous studies indicate that psilocybin impairs attention across multiple assessment paradigms.
Inhibition
Inhibitory control was assessed with the Go/No-Go task and the Stroop Test. A high dose of LSD (100 μg) impaired motor response inhibition in the Go/No-Go task taken 200 minutes after administration (Schmidt et al., 2018). This impairment was correlated with subjective reports of cognitive dysfunction, which, according to the authors, may have exacerbated the failure to inhibit responses during No-Go trials. In one of the previous studies (Hutten et al., 2020), 63% of participants reported a perceived decline in performance, although performance enhancement was observed in 74% of participants on the PVT in the same study. These findings suggest that at higher doses, subjective beliefs about cognitive impairment may play a more pronounced role in influencing actual performance outcomes compared to lower doses. The effects of high-dose psilocybin (260 μg/kg) in the Stroop Test were measured 85 minutes after the drug administration in another study (Quednow et al., 2012). Psilocybin selectively increased errors in high-conflict (incongruent) trials in the Stroop Test and slowed RTs across all conditions and worsened Stroop interference, indicating reduced ability to inhibit distractions. These alterations were attenuated by ketanserin pretreatment. Thus, higher doses of psychedelics may be needed to cause significant changes in the Stroop Test.
However, inhibitory control is less impacted by microdoses. Cavanna et al. (2022) found no significant effects of psilocybin microdosing on the Go/No-Go task, and although increased RTs were observed in the Stroop Test, they were not statistically robust after correction for multiple comparisons. Similarly, Wießner et al. (2022) reported no changes in Stroop Test performance following 50 μg LSD, measured 24 hours post-administration.
Psychedelics’ effects on cognitive flexibility
Cognitive flexibility (CF) has been defined as the ability to appropriately adjust one’s behavior according to a changing environment (Dajani and Uddin, 2015) and has been found to act as a buffer between stress and negative psychological outcomes (Gloster et al., 2017). CF was assessed in analyzed studies by using tasks such as the Intra/Extra-Dimensional Shift Task (IED; Owen et al., 1991), the Wisconsin Card Sorting Test (WCST; Heaton, 2005), the Cognitive Control Task (CCT; de Wit et al., 2012), part B of the TMT (Reitan and Wolfson, 1995) and the probabilistic reversal learning task (Rostami Kandroodi et al., 2021) (see Table 2). The IED is based on the WCST paradigm, which requires participants to sort cards according to a rule that changes unpredictably. Studies investigating the effects of psychedelics on cognitive flexibility used microdoses (5, 10, 20 μg; Hutten et al., 2020; 0.5 g of dried mushrooms; Cavanna et al., 2022), low doses (50 μg; Wießner et al., 2022) and medium doses (75 and 100 μg; Kanen et al., 2023; Pokorny et al., 2020).
Summary of studies investigating the effects of psychedelics on cognitive flexibility.

Note. IED = Intra/Extra-Dimensional Shift Task; LSD = lysergic acid diethylamide; WCST = Wisconsin Card Sorting Test; CCT = Cognitive Control Task; RT = reaction time; TMT = Trail Making Test.
In a randomized, double-blind, placebo-controlled crossover study of 24 healthy participants, LSD (50 μg) reduced CF as measured by the WCST at baseline (2 hours before drug administration) and 24 hours post-administration (Wießner et al., 2022). More specifically, LSD reduced categories achieved, conceptual level responses percentage and increased errors, particularly perseverative errors, indicating a tendency to stick to incorrect rules despite feedback. Similarly, a separate randomized, placebo-controlled within-subject study with 25 healthy participants demonstrated an acute reduction in CF following LSD (100 μg) administration, measured 220 minutes post-administration using the IED as there were more errors in the LSD condition compared to placebo or LSD+ketanserin (Pokorny et al., 2020). LSD also increased latency in the extra-dimensional shift (EDS) stage (where participants had to shift attention to a previously irrelevant dimension) of the IED task compared to placebo. Thus, according to these two studies, medium doses of LSD impair CF acutely and post-acutely.
Contrary to previous findings, 75 μg dose of LSD enhanced aspects of cognitive flexibility in the probabilistic reversal learning task conducted 5 hours after administration (Kanen et al., 2023). Participants showed increased learning rates from rewards (biggest effects) and punishments, as well as exploration over rigid stimulus-response patterns, measured by decreased stimulus stickiness. Microdoses of LSD have been found not to influence CF as measured with the CCT 2.5 hours after LSD consumption, which was used to assess participants’ level of cognitive control, that is, the ease of shift between habitual and goal-directed behavior (Hutten et al., 2020). Psilocybin microdoses did not significantly impact performance in part B of the TMT, where it is required to alternate between numbers and letters (e.g., 1-A-2-B-. . .) while connecting circles, indicating a lack of effect on CF (Cavanna et al., 2022).
Psychedelics’ effects on memory
Memory is defined as the faculty of encoding, storing and retrieving information. Memory can be categorized into short-term memory, working memory and long-term memory (Cowan, 2008). Most of the studies here looked at the effects of psychedelics on working memory, which refers to a system that provides temporary storage and manipulation of the information necessary for cognitive tasks.
The test for measuring memory in the analyzed experimental studies were the visual-manual delayed response task (DRT; Park and Holzman, 1992), the Spatial Span test (SSP) from the CANTAB (Cambridge Cognition, 2016), the letter N-back task (Jaeggi et al., 2008), Rey-Osterrieth Complex Figure (ROCF; Rey, 1999), the 2D Object-Location Memory Task (OLMT; Rasch et al., 2007), the spatial working memory (SWM) and the paired associates learning (PAL) from the CANTAB (Cambridge Cognition, 2016), the Rey Auditory-Verbal Learning Test (RAVLT; Bezdicek et al., 2013), the Groton maze learning task (GMLT; Snyder, 2005), the Paired Associative Learning Test (PALT; Dudysová et al., 2016), the Tower of London test (TOL; Phillips et al., 2001), the Spatial Memory Test (SMT; Mallaroni et al., 2023) and the Emotional Episodic Memory Task (Doss et al., 2024) (see Table 3). Some of these tests also involve other executive functions, for example, the letter N-back task also requires the use of selective attention and cognitive inhibition. Psilocybin dosages included low doses such as 115 μg/kg (Wittmann et al., 2007), 10 mg/70 kg (Barrett et al., 2018) and 15 mg (Doss et al., 2024), medium doses like 0.25 mg/kg (Vollenweider et al., 1998), 20 mg/70 kg (Barrett et al., 2018), 15 mg (Mallaroni et al., 2023) and 215 μg/kg (Carter et al., 2005), and high doses including 0.26 mg/kg (Nikolič et al., 2023), 30 mg/70 kg (Barrett et al., 2018) and 250 μg/kg (Wittmann et al., 2007). LSD was administered in dosages such as 5, 10, 20 μg of LSD (microdoses; Family et al., 2020), 13 or 26 μg (microdoses; de Wit et al., 2022), 50 μg (low dose; Wießner et al., 2022), 75 μg (medium dose; Speth et al., 2016) and 100 μg (medium dose; Pokorny et al., 2020).
Summary of studies investigating the effects of psychedelics on memory.
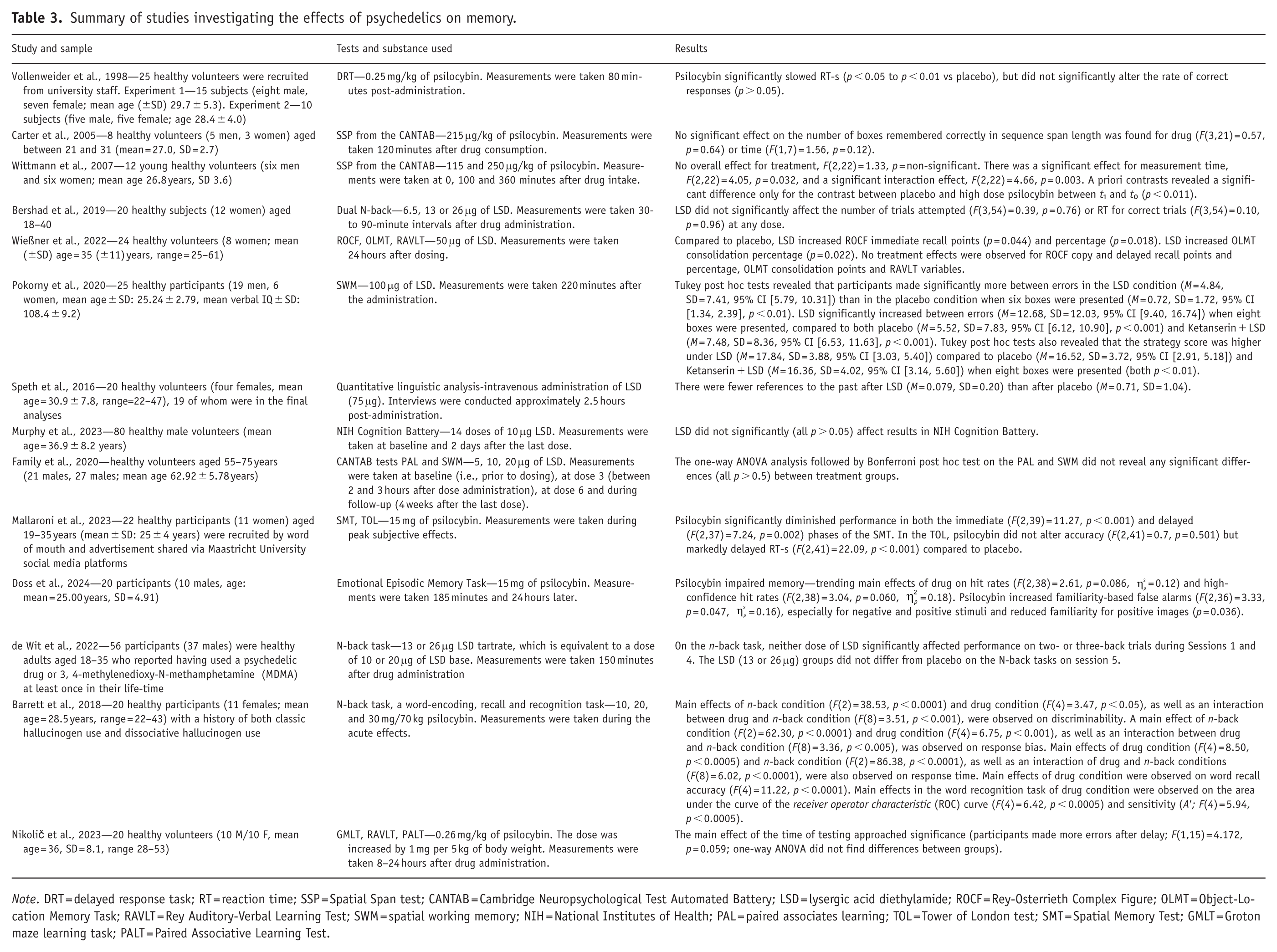
Note. DRT = delayed response task; RT = reaction time; SSP = Spatial Span test; CANTAB = Cambridge Neuropsychological Test Automated Battery; LSD = lysergic acid diethylamide; ROCF = Rey-Osterrieth Complex Figure; OLMT = Object-Location Memory Task; RAVLT = Rey Auditory-Verbal Learning Test; SWM = spatial working memory; NIH = National Institutes of Health; PAL = paired associates learning; TOL = Tower of London test; SMT = Spatial Memory Test; GMLT = Groton maze learning task; PALT = Paired Associative Learning Test.
Visuospatial working memory and SWM
Visuospatial working memory was commonly assessed using the SSP. In a study by Carter et al. (2005), psilocybin at a moderate dose (0.215 μg/kg) did not impair SSP performance when administered 120 minutes post-ingestion, during peak subjective effects. Specifically, no significant changes were observed in the span length, that is, the number of boxes remembered correctly in sequence. However, a subsequent study using a higher dose of psilocybin (250 μg/kg) reported impairments in the SSP performance when measured at 0, 100 and 360 minutes post-administration, whereas a medium dose (115 μg/kg) did not produce such effects (Wittmann et al., 2007). These results suggest that only higher doses of psilocybin significantly disrupt visuospatial working memory as measured by the SSP.
The SMT, which measures visuospatial memory and reasoning, also revealed psilocybin-induced deficits. Specifically, a dose of 15 mg significantly reduced immediate and delayed recall performance (+30 minutes) (Mallaroni et al., 2023). In contrast, sub-acute enhancement of visuospatial memory has been observed following LSD administration (Wießner et al., 2022). LSD at a dose of 50 μg significantly improved performance on the ROCF and the OLMT when assessed 24 hours after drug intake.
Spatial working memory was assessed using the SWM. Microdoses of LSD were found not to significantly alter SWM performance (Family et al., 2020). The PAL, which measures visual memory and learning, did not also change significantly. However, higher doses of LSD (100 μg) led to significant impairments, including increased between errors and reduced strategic planning (Pokorny et al., 2020). Importantly, these effects were only present when the cognitive load was high. These effects emerged 220 minutes post-administration and were reversed by ketanserin, implicating the 5-HT2A receptor in LSD-induced SWM deficits.
Working memory and executive function
In a study using the DRT, psilocybin (0.25 mg/kg) significantly slowed RT without reducing accuracy (Vollenweider et al., 1998). This effect was not correlated with subjective intensity of psychedelic experience measured with the Altered State of Consciousness rating scale but was prevented by ketanserin, again indicating 5-HT2A receptor mediation. More complex tasks such as the N-back have revealed dose-dependent impairments under psilocybin. Barrett et al. (2018) found that psilocybin at doses of 10, 20 and 30 mg/70 kg impaired performance particularly in the 2-back condition, where both reaction times and accuracy were reduced. These effects were not observed in the 0-back condition, suggesting that the drug selectively affects working memory rather than attention per se. Similarly, psilocybin disrupted episodic memory tasks involving word-encoding, free recall and recognition, impairing both recollection and familiarity.
In contrast, microdoses of LSD (13–26 μg) did not significantly affect N-back task performance when measured during peak drug effects (150 minute post-ingestion) (de Wit et al., 2022). Interestingly, when subjects were asked to rate (on a 7-point scale) how well they thought they performed on the task, subjects in the high-microdose LSD group self-reported performing significantly above average relative to other participants. A separate study using the dual N-back task, which requires the ability to manage two n-back tasks simultaneously, also found no significant effect of LSD microdoses (6.5–26 μg) on working memory performance (Bershad et al., 2019).
Psilocybin did not significantly affect performance in the TOL, which measures planning, inhibition, impulsivity and working memory, although delayed RT-s were observed during peak effects (Mallaroni et al., 2023). Similarly, in a home-administered randomized controlled trial, LSD did not produce significant post-acute effects on the NIH Cognition Battery, that measures episodic memory, language (vocabulary and reading), processing speed, working memory, attention, inhibitory control and attention and the set-shifting component of executive function (Murphy et al., 2023).
Episodic memory
Psilocybin (10–30 mg/70 kg) was found to impair episodic memory during encoding and retrieval, affecting both free recall and recognition accuracy (Barrett et al., 2018). Psilocybin (15 mg) impaired the encoding of episodic memory, particularly for neutral and positive stimuli in a study by Doss et al. (2024). This was evidenced by reduced hit rates and high-confidence hit rates in the Emotional Episodic Memory Task conducted the day after drug administration. Contrary to the hypothesis that psychedelics would enhance familiarity, the study found that both psilocybin and 4-bromo-2,5-dimethoxyphenethylamine (2C-B; a synthetic psychedelic drug) impaired estimates of familiarity. However, this impairment was likely due to a misattribution of heightened familiarity, as both drugs increased familiarity-based false alarms, especially for negative and positive stimuli. Memory performance was generally best for negative stimuli under placebo conditions, but psilocybin disrupted this pattern. Psilocybin selectively increased false alarms for emotional (negative and positive) stimuli, suggesting a distortion in how emotional memories are processed.
Autobiographical memory and memory consolidation
While LSD appears to modulate autobiographical memory, as evidenced by Speth et al. (2016) who reported that a 75 μg dose significantly reduced mental time travel to past events—indicating decreased attentional focus on autobiographical content assessed through interviews 2.5 hours post-administration—multiple objective cognitive tests have consistently found no significant effects on memory consolidation. Specifically, Nikolič et al. (2023) observed no impact of psilocybin (0.26 mg/kg) on memory performance across several measures, including the RAVLT, the PALT and the GMLT. Likewise, Wießner et al. (2022) reported no changes in auditory-verbal memory consolidation under LSD, as assessed by the RAVLT.
Psychedelics’ effects on emotional processing
Emotional processing refers to perceiving, expressing and managing emotions. Several tests were used in the current studies, such as the Facial Emotion Recognition Task (FERT; Bedi et al., 2010), the Reading the Mind in the Eyes Test (Baron-Cohen et al., 2001), the Emotional Go/No-go Task (Kometer et al., 2012), the Emotional Images Task (Lang et al., 1997; Wardle, 2012), the Emotional Faces Task (Griffiths et al., 2015), the Cyberball Task (Williams and Jarvis, 2006), an emotional face discrimination task (Grimm et al., 2018), two facial affect discrimination tasks (Schmidt et al., 2013) and paradigm of facial stimuli (Mueller et al., 2017) (see Table 4). Thus, these tests primarily focused on the perception of emotions. For psilocybin, low doses included 0.16 mg/kg (Grimm et al., 2018), 115 μg/kg (Schmidt et al., 2013) and medium doses included 215 μg/kg (Kometer et al., 2012). In LSD studies, low doses included 13 μg (de Wit et al., 2022), 26 μg (de Wit et al., 2022) and medium doses included 100 μg (Dolder et al., 2016; Mueller et al., 2017) and high doses included 200 μg (Dolder et al., 2016).
Summary of studies investigating the effects of psychedelics on emotional processing.

Note. FERT = Facial Emotion Recognition Task; LSD = lysergic acid diethylamide; RT = reaction time.
Dolder et al. (2016) demonstrated that 100 and 200 μg doses of LSD impaired the recognition of fearful faces in the FERT, measured 5 and 7 hours post-administration (during the acute effects of the drug). In contrast, Mueller et al. (2017) found that 100 μg of LSD had no significant effect on RT, accuracy or omission rates (absence of button presses) in an fMRI paradigm conducted 2.5 hours after consumption—timed to coincide with the subjective and pharmacological peak effects. During fMRI scanning, participants viewed a standardized set of facial stimuli consisting of 10 unique identities, each displaying neutral, 50% fearful and 100% fearful expressions (totaling 30 stimuli). Brain activity, accuracy and RT-s were recorded. The discrepancy between these studies may stem from differences in the tasks employed (FERT vs fMRI-based emotion processing) or the timing of assessments relative to LSD’s pharmacokinetic profile.
Kometer et al. (2012) found that a dose of 215 μg/kg psilocybin attenuated the recognition of negative facial expressions in the Reading the Mind in the Eyes Test (administered 130 minutes post-ingestion). In the Emotional Go/No-Go Task, where cues were defined by emotional valence (unlike the standard shape-based version), psilocybin disproportionately increased RT-s for negative and neutral words compared to positive words. It also significantly elevated error rates for neutral—but not positive or negative—words. Notably, error rates were higher for negative versus positive stimuli under psilocybin, an effect absent under placebo. These valence-specific effects on error rates were consistent across Go/No-Go conditions and unaffected by ketanserin pretreatment. Grimm et al. (2018) observed that a low psilocybin dose (0.16 mg/kg) slowed RTs across all affective categories (negative, neutral, positive) in an emotional face discrimination task during acute effects, though accuracy remained intact and no drug-by-face-type interaction emerged. Schmidt et al. (2013) reported that 115 μg/kg psilocybin impaired the discrimination of fearful (but not happy) faces relative to neutral ones. This suggests psilocybin’s behavioral effects are selectively modulated by stimulus valence. Collectively, low-to-medium psilocybin doses acutely impair the processing of negative stimuli, an effect that appears partially independent of 5-HT2A receptor activation.
A study by de Wit et al. (2022) examined the effects of LSD microdoses (13 or 26 μg) on emotional and social processing using the Cyberball Task, the Emotional Images Task and the Emotional Faces Task. In the Emotional Images Task, neither LSD dose significantly affected ratings of positive or negative images during compared to placebo. However, in the Emotional Faces Task, 26 μg of LSD reduced false alarm rates for fearful faces on the first and last days of administration, without altering hit rates for any emotion. During the Cyberball Task, the same dose (26 μg) attenuated negative mood ratings in the social rejection, though it had no effect during social acceptance. These effects did not persist at follow-up (3–4 days post-session).
However, Bershad et al. (2019) observed no significant impact of LSD microdoses on the Cyberball Task. While their Emotional Images Task results were also unaffected, the highest dose (26 μg) slightly reduced positivity ratings for positive images. Similarly, psilocybin microdoses showed no measurable effects on emotional processing in the Emotional Go/No-Go Task (Marschall et al., 2021). Collectively, these studies suggest that LSD microdoses may transiently modulate specific aspects of emotional and social processing (e.g., fear perception, rejection sensitivity), but these effects are subtle, dose-dependent and do not endure beyond acute administration.
Psychedelics’ effects on empathy
Empathy can be categorized as cognitive or emotional. Cognitive empathy involves knowing how other people think and feel, while emotional empathy involves feeling another person’s emotions. In most of the analyzed studies, empathy was measured acutely or post-acutely with the Multifaceted Empathy Test (MET; Dziobek et al., 2008) that was developed to assess cognitive and emotional aspects of empathic functioning. Emotional empathy is categorized into two in this test: (1) implicit empathy, often referred to as “arousal,” includes the automatic, unconscious aspect of emotional empathy in which the sharing of emotions arouses the observing individual, (2) explicit empathy, which is a deliberate and conscious process requiring mental effort and conscious processing, enabling us to respond appropriately and empathetically toward other people’s emotions.
Studies investigating the effects of psychedelics on empathy employed LSD doses of 100 µg (medium dose), 200 µg (high dose) (Dolder et al., 2016), 25, 50, 100 and 200 µg (from low to high dose) (Holze et al., 2021). Psilocybin studies utilized a dose of 0.215 mg/kg (medium dose) (Jungwirth et al., 2025; Pokorny et al., 2017) and a dose of 15 mg (medium dose) (Mallaroni et al., 2023). Investigations on ayahuasca reported DMT concentrations ranging from 14.1 to 20.1 mg (medium to high dose) (Uthaug et al., 2021) (see Table 5).
Summary of studies investigating the effects of psychedelics on empathy.
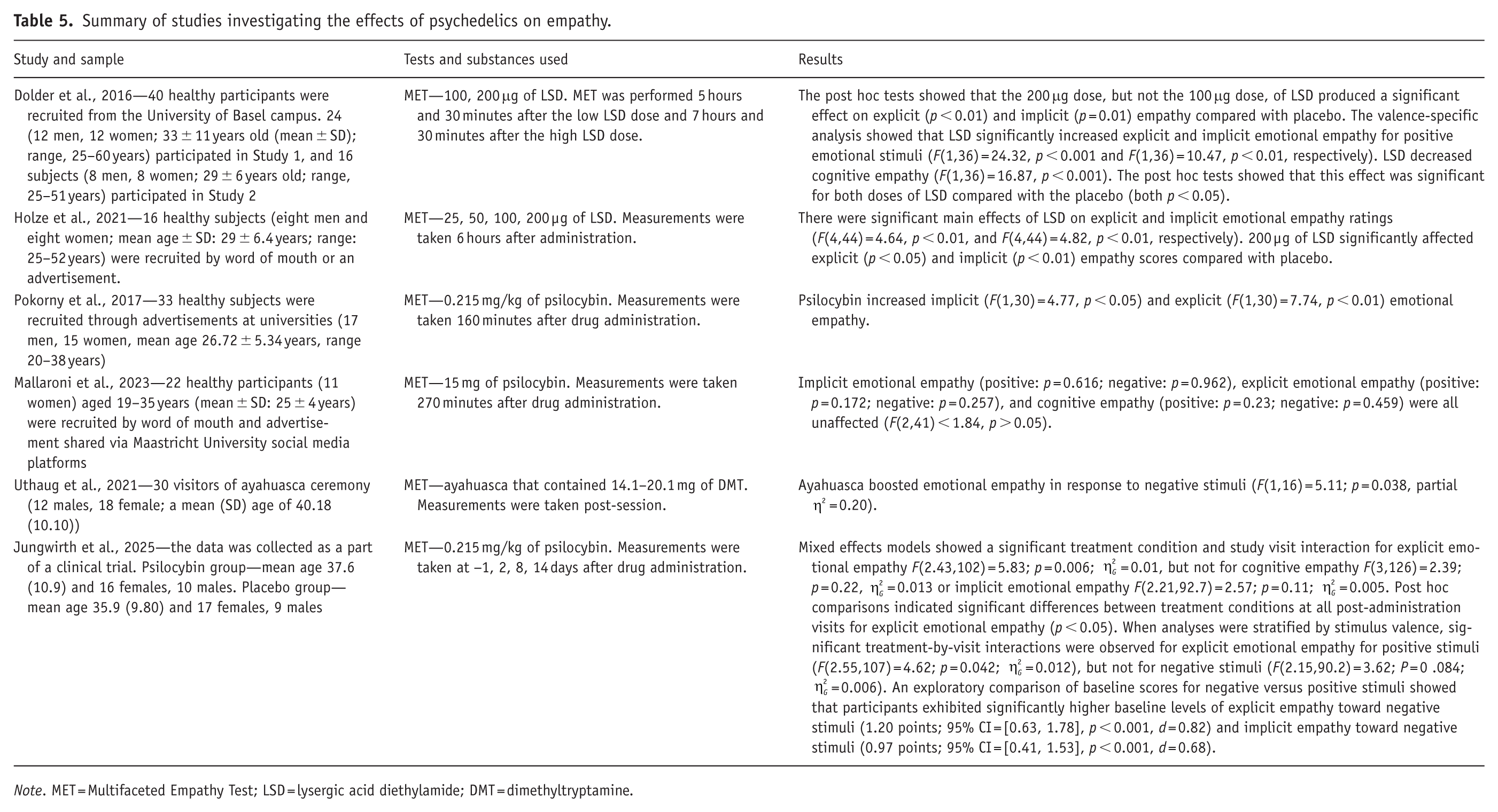
Note. MET = Multifaceted Empathy Test; LSD = lysergic acid diethylamide; DMT = dimethyltryptamine.
Dolder et al. (2016) looked at the effects of 100 and 200 μg of LSD on empathy. Most significantly, 200 μg of LSD enhanced implicit and explicit emotional empathy as measured with the MET 7 hours and 30 minutes after the administration, which accounts for 50% of the peak subjective effects. The valence-specific analysis showed that a higher dose of LSD significantly increased explicit and implicit emotional empathy scores for positive emotional stimuli. Both doses of LSD significantly reduced cognitive empathy; for the lower dose the MET was performed 5 hours and 30 minutes after the administration. Similar results were found in another study where 200 μg of LSD increased implicit and explicit emotional empathy measured with the MET 6 hours after administration (Holze et al., 2021). It is interesting to note that ketanserin did not have an effect on the result—thus the effects of LSD on empathy could be 5-HT2A receptor independent. Lower doses of LSD (25, 50, 100 μg) did not significantly impact empathy, but the authors point out that the results could have been different when measured at 3 hours, which is approximately 50% of the peak subjective effects for lower doses.
An amount of 0.215 mg/kg of psilocybin was found to increase explicit and implicit emotional empathy independent of stimulus valence (Pokorny et al., 2017). The MET was performed 160 minutes after the administration, when according to the authors, the peak perceptual/visual effect of psilocybin had already markedly subsided. However, some studies suggest that 160 minutes at 0.215 mg/kg might still have considerable visual effects present (Preller and Vollenweider, 2016). Additionally, there is some indication that a moderate dose might disrupt vigilance more than a high dose of psilocybin as measured with the “Five Dimensional Altered States of Consciousness” scale (5D-ASC; Hasler et al., 2004). Psilocybin did not significantly influence cognitive empathy. 0.215 mg/kg of psilocybin was administered to 51 people with depression in another study (Jungwirth et al., 2025). Psilocybin significantly enhanced explicit emotional empathy, particularly due to increased empathy toward positive stimuli, compared to the placebo group, with effects lasting at least 2 weeks. At baseline, participants showed significantly stronger empathy towards negative compared to positive stimuli, which could be linked to depression. However, no significant correlations between increased empathy and improvement in depressive symptoms were found in the present study. In contrary to the previous studies, 15 mg of psilocybin has been found to have no effect on empathy measured with the MET 270 minutes after drug administration (Mallaroni et al., 2023).
While the impact of ayahuasca on empathy has been less extensively studied, Uthaug et al. (2021) observed a notable treatment × time interaction in implicit emotional empathy, revealing that ayahuasca (containing 14.1–20.1 mg of DMT) boosted emotional empathy in response to negative stimuli. The MET was performed post-acutely.
Quality rating
All of the 6 cross-over and 26 parallel-design studies were rated as having a high risk of bias (measured with the Cochrane RoB tool; Sterne et al., 2019; see Appendices S1 and S2). These results are connected to the unblinding effects of psychedelics and to the likelihood that measurement of outcomes differed between interventions within each sequence, given that the pronounced and distinct subjective effects of psychedelics can influence participant expectations, assessor behavior and the context in which outcomes are measured (Aday et al., 2022).
Discussion
The effects of psychedelics on cognitive and psychological functions
This systematic review included 32 placebo-controlled studies (see Table 6), mostly done in healthy volunteers. Dose-dependent impairments were seen in many tasks assessing RT, attention and inhibition (Barrett et al., 2018; Carter et al., 2005; Cavanna et al., 2022; Hutten et al., 2020; Mallaroni et al., 2023; Schmidt et al., 2018; Vollenweider et al., 2007; Quednow et al., 2012), although some studies found no effects (Bershad et al., 2019; de Wit et al., 2022; Family et al., 2020; Wießner et al., 2022). The results on CF were less clear, although the small number of studies limits strong conclusions. While moderate doses of LSD (50–100 μg) acutely and post-acutely impaired CF (Pokorny et al., 2020; Wießner et al., 2022), other studies reported enhanced CF at a 75 μg dose (Kanen et al., 2023) or no effect following microdosing (Cavanna et al., 2022; Hutten et al., 2020). Psychedelics impaired (Barrett et al., 2018; Doss et al., 2024; Mallaroni et al., 2023; Pokorny et al., 2020; Vollenweider et al., 1998; Wittmann et al., 2007), enhanced (Wießner et al., 2022) or had no effect (Bershad et al., 2019; Carter et al., 2005; de Wit et al., 2022; Family et al., 2020; Murphy et al., 2023; Nikolič et al., 2023) on memory depending on the task, dosage and timing of the assessment. Regarding emotional processing, several studies found impaired recognition of specifically negative stimuli (Dolder et al., 2016; Kometer et al., 2012; Schmidt et al., 2013). Psychedelics enhanced implicit and explicit emotional empathy, but most of the time had no effect on cognitive empathy (Dolder et al., 2016; Holze et al., 2021; Jungwirth et al., 2025; Pokorny et al., 2017; Uthaug et al., 2021). Most of the studies assessed the acute effects of psychedelics. Post-acute measurements were usually taken hours or days later.
The effects of psychedelics on tasks and functions they assess.

Many tests for measuring RT also assessed attention and inhibitory control as they are intertwined. Microdoses of LSD (5–26 μg) showed mixed results, with one study reporting enhanced sustained attention (Hutten et al., 2020) and others showing no significant changes (Bershad et al., 2019; de Wit et al., 2022). Medium (15–30 mg; 0.28 mg/kg) and high (⩾215 μg/kg) doses of psilocybin impaired performance in several attention tasks (Barrett et al., 2018; Quednow et al., 2012; Umbricht et al., 2003; Vollenweider et al., 2007). Higher doses of LSD (50–100 μg) and psilocybin also slowed RT and reduced inhibitory control (Schmidt et al., 2018; Quednow et al., 2012), whereas microdoses of psilocybin (0.5 g dried mushrooms) showed non-significant trends toward impaired RT (Cavanna et al., 2022). Overall, while low doses of psychedelics may have subtle or negligible effects on RT and attention, medium to high doses tended to impair performance.
Only five CF articles met criteria for placebo-controlled study standards. All of these studies, except one, were done with LSD (microdoses to medium doses) in healthy volunteers. Two of them found that LSD impairs CF acutely (Pokorny et al., 2020) and post-acutely (Wießner et al., 2022), whereas one study found enhanced CF during LSD subjective effects (Kanen et al., 2023). The difference in the results could have been influenced by different tests used as the IED and the WCST are based on the same principle, but the probabilistic reversal learning task can be looked at as a reinforcement learning test. Microdoses of LSD and psilocybin did not influence CF that is in line with several previous studies that have found no significant effects of microdosing on cognition (Bershad et al., 2019; Cavanna et al., 2022).
Psilocybin and LSD exerted dose- and time-dependent effects on memory, with acute impairments often observed in working and episodic memory tasks, particularly at medium to high doses (Barrett et al., 2018; Mallaroni et al., 2023; Pokorny et al., 2020; Vollenweider et al., 1998; Wittmann et al., 2007). However, some studies reported no acute effects on working memory (Bershad et al., 2019; Carter et al., 2005; de Wit et al., 2022; Family et al., 2020) or post-acute improvements in memory consolidation and visuospatial memory (Wießner et al., 2022). Changes in autobiographical memory during acute effects were also observed (Speth et al., 2016). Psilocybin was found to impair episodic memory (Barrett et al., 2018; Doss et al., 2024). Regarding memory consolidation it was found that psilocybin does not affect daytime or sleep-related declarative memory consolidation in healthy volunteers (Nikolič et al., 2023).
Research on psychedelics’ modulation of emotional processing revealed dose- and valence-dependent effects, primarily characterized by attenuated responses to negative stimuli without consistent alterations in positive emotion perception. Medium to high doses of LSD (100–200 μg) impaired fear recognition in the FERT during acute effects (Dolder et al., 2016), though discrepant fMRI findings suggest task-dependent variability (Mueller et al., 2017). Psilocybin demonstrated valence-specific blunting of negative emotion processing: medium doses (215 μg/kg) reduced recognition of negative facial expressions (Kometer et al., 2012), while low doses (0.16 mg/kg) slowed RT-s across emotional stimuli without affecting accuracy (Grimm et al., 2018). Microdoses of LSD (13–26 μg) showed limited acute effects, with isolated reductions in fear false alarms and transient mood improvements during social rejection (Cyberball Task; de Wit et al., 2022). Neither LSD nor psilocybin microdoses reliably altered emotional processing in standardized tasks (Bershad et al., 2019; Marschall et al., 2021).
As mentioned, Grimm et al. (2018) showed 0.16 mg/kg of psilocybin slowed RTs for negative, neutral and positive faces without having a significant effect on accuracy. Research on psilocybin’s effects has shown differing results depending on the task paradigm. In an emotional go/no-go task, psilocybin significantly increased RTs more for negative and neutral words compared to positive ones (Kometer et al., 2012), but slowed RT across all conditions in a Stroop Test in another study (Quednow et al., 2012). Further research is needed to thoroughly assess whether affective psychedelic effects are dependent on emotional valence. However, these findings suggest that psilocybin’s effects could be highly dependent on the specific cognitive and emotional demands of the task.
Current studies on empathy demonstrated that psychedelics exert distinct, dose-dependent effects on cognitive and emotional empathy. All of the studies on empathy were conducted with MET, which makes it easier to compare the results and makes them more reliable. High dose of LSD (200 μg) enhanced both implicit and explicit emotional empathy, particularly for positive stimuli, when measured during the acute phase, while both medium (100 μg) and high doses simultaneously reduced cognitive empathy (Dolder et al., 2016). Similar results on emotional empathy were found by Holze et al. (2021). Psilocybin showed more variable outcomes: a dose of 0.215 mg/kg increased emotional empathy independent of stimuli valence in healthy volunteers (Pokorny et al., 2017) and depressed patients (Jungwirth et al., 2025), with the latter study showing again increased empathy toward positive stimuli. However, 15 mg of psilocybin showed no significant effects on empathy in another study (Mallaroni et al., 2023). Ayahuasca appeared to specifically enhance emotional empathy for negative stimuli when measured post-acutely (Uthaug et al., 2021). Overall, classic psychedelics enhance explicit and implicit emotional empathy without affecting cognitive empathy. A recent meta-analysis that looked at the effects of classic psychedelics on MET while also incorporating open-design studies, found similar results (Olami and Peled-Avron, 2024).
Only one placebo-controlled study looked at the effects of psychedelics in clinical populations, more precisely in people diagnosed with depression (Jungwirth et al., 2025). While these results might not generalize to the wider population they are valuable for clinical populations as cognitive and psychological functions often exhibit disturbances in several mental health disorders (Grant and Chamberlain, 2023; Millan et al., 2012; Robinson and Roiser, 2016). Future studies should assess whether psychedelics impact cognitive and psychological functions similarly in clinical and non-clinical populations.
There were only four studies (de Wit et al., 2022; Family et al., 2020; Jungwirth et al., 2025; Murphy et al., 2023) that looked at longer-lasting post-acute (>24 hours) effects of drug experience. Three of these studies that assessed the effects of 5, 10, 20 μg (Family et al., 2020), 14 doses of 10 μg (Murphy et al., 2023) and 13 or 26 μg of LSD did not find significant longer-lasting changes (de Wit et al., 2022). The post-acute measurements were taken 4 weeks, 2 days and 3–4 days later, respectively. In a study by Jungwirth et al. (2025) measurements were taken 2, 8 and 14 days after psilocybin administration. Psilocybin significantly enhanced explicit emotional empathy with effects lasting at least 2 weeks. These results leave open the possibility that higher doses could have longer-lasting effects on different functions.
While the primary consciousness-altering effect of classic psychedelics is highly dependent on 5-HT2A receptor agonism, many alterations in cognitive functions likely also require activity at other receptors. Multiple studies utilized ketanserin (a relatively selective 5-HT2A/5-HT2C receptor antagonist) to assess the role of 5-HT2A receptors finding mixed results. Ketanserin was effective in attenuating psilocybin’s effects in attention tasks (Stroop Task—Quednow et al., 2012; FAIR task—Vollenweider et al., 2007) and blocked LSD-induced impairments in CF (IED task) and working memory (SWM task) (Pokorny et al., 2020). Ketanserin also reduced psychedelic reaction time deficits in the working memory DRT task (Vollenweider et al., 1998). However, ketanserin did not completely block the effects of psilocybin in a multiple-object tracking attention task (Carter et al., 2005), in the Emotional Go/No-Go task (Kometer et al., 2012) and did not alter LSD’s effects on empathy (Holze et al., 2021). The role of the 2A-receptor on attention and inhibition seems to be task-dependent as ketanserin attenuates the impairing effects of psilocybin in the Stroop Test (Quednow et al., 2012), but does not affect performance in a multiple-object tracking task (Carter et al., 2005). These results demonstrate that 2A-receptors are likely responsible for psychedelic-induced impairments in cognitive flexibility and memory, but attention, emotional processing and empathy are affected by more complex pharmacological pathways.
Decoupling the internal and external
One of the more characteristic and general effects of psychedelics is altering the interaction between top-down (internal) and bottom-up (external) processing (Carhart-Harris and Friston, 2019; Vollenweider and Preller, 2020). This, however, strongly complicates utilizing many standard cognitive tests primarily designed for use in unperturbed states of consciousness. A deeper look into the results of several studies suggests that standard tests might not capture the intended effects but predominantly indicate impairment in interacting with the external test. For instance, in one study utilizing a custom attention task, subjects under the effects of psilocybin reported that they understood the task requirements, but it was still harder to perform (Carter et al., 2005). Similarly, performance and continuity value of the FAIR, which capture primarily task-interaction, are much more strongly affected by psychedelics than the quality value, which measures the level of attention when interacting with the test (Hasler et al, 2004; Vollenweider et al, 2007). Quality value seems to be only significantly affected at very high doses.
Similar picture emerges from the TMT paradigm. Microdoses of psilocybin increased the time required for part A of the TMT (measures attention and coordination), but did not cause significant changes in part B of the TMT (measures task-switching ability, CF, working memory; Cavanna et al., 2022). These results suggest that while attentional capacity is impaired at lower doses, cognitive or executive functioning impairment seems to require higher doses, providing tentative evidence that external stimulus processing (i.e., task-based attention) and internally-directed processing (e.g., executive functioning, task-switching) are considerably affected at different doses suggesting that true effects of psychedelics on various cognitive functions might be occluded by impairment of external stimulus processing happening at already lower doses. Various forms of increased cognitive functioning, such as increased sensory acuity, mindfulness, insightfulness, absorption, mental clarity, cognitive flexibility, autonomy and broadened attention are also commonly reported during psychedelic experiences (Amada et al., 2020; Carter et al., 2005; Day and Schmetkamp, 2022; Watts et al., 2017; Wolff et al., 2019), which could highlight increased flexibility or attention within internal mentation decoupled from sensory information.
While perfect solutions to these issues are hard to envision, some methods can be useful. First, utilizing specialized experimental designs with simplified or intuitive task interactions or using more immersive non-screen displays. Second, developing environmental settings as behavioral assays where participants naturally interact with the environment and their interactions can be measured or quantified. Third, leaning into subjective methodologies and avoiding the common objectivist tendency to undervalue or even neglect self-report. Adapting various novel phenomenological methodologies (e.g., microphenomenological interviews, precise theoretical and hermeneutical approaches) to altered states research is still vital to make the most out of report-based paradigms. In line with all this, developing a validity framework for measures designed with altered states of consciousness in mind would considerably support and streamline research in psychedelic cognitive science.
Contextual and Stimulus-Dependent Effects of Psychedelics on Perception
LSD and psilocybin appear to modulate emotional perception and attentional processes in ways that depend on both stimulus content and contextual factors. Dolder et al. (2016) found that moderate to high doses impaired the recognition of fearful faces but did not significantly affect the recognition of neutral, angry or happy faces. This finding could mean that the effects of LSD vary depending on the emotional content of the sensory stimulus and thus different aspects of the environment catch attention. While the impaired recognition of fearful faces might suggest a more positive subjective experience—potentially due to a diminished perception of fear—psychedelic experiences are often emotionally ambivalent (Gashi et al., 2021). Dolder et al. (2016) findings with psilocybin were not replicated in subsequent studies using LSD (Mueller et al., 2017) or psilocybin at different doses (Grimm et al., 2018). Additionally, a less explored area is how various types of open-eye visuals induced by psychedelics—such as enhancements, transformations and overlays—interact with attention allocation (Swanson, 2024). For example, transformations and overlays might have bigger effects on attentional tasks performance as they change visual stimuli more profoundly, that may result in broadened attention (Carter et al., 2005). Investigating these aspects could provide insights into the interplay between perception and attention during psychedelic states.
Limitations
Despite the growing body of placebo-controlled studies (see Table 6) investigating the cognitive and psychological effects of psychedelics, several methodological and conceptual limitations complicate the interpretation and generalization of findings. These limitations concern expectancy effects (Olson et al., 2020), the choice of comparator substances (Doss et al., 2024), timing of cognitive assessments (see Tables 1–6), participant characteristics (see Tables 1–6) and differences between psychedelic compounds (Griffiths et al., 2019; Lawrence et al., 2022). Addressing these issues is essential for improving the reliability and ecological validity of future research.
Although only placebo-controlled studies were used in this systematic review, unblinding is a common problem in psychedelic research (Aday et al, 2022; Olson et al., 2020). One key factor contributing to placebo effects is expectancy, which can significantly influence outcomes across various types of interventions. Expectancy has been linked to improved therapeutic outcomes with escitalopram (a common selective serotonin reuptake inhibitor; SSRI), whereas expectancy did not predict treatment response to psilocybin (Szigeti et al., 2024). This suggests that the effects of psilocybin, and potentially other psychedelics, cannot be fully explained by placebo alone. Self-reported expectancy may be a worse predictor of drug effects for psychedelics as they have a strong effect on cognition and consciousness (Nutt et al., 2020). Incorporating a similar approach into future studies on cognition—by asking participants to report their expectations before administration—could help disentangle the influence of expectancy on results.
Some of the studies used other drugs as comparison to classic psychedelics, for example, 2C-B, dextromethorphan and ketamine were used (Barrett et al., 2018; Doss et al., 2024; Mallaroni et al., 2023; Schmidt et al., 2013), that have somewhat similar phenomenology to classic psychedelics. As they still have considerable effect on the state of consciousness they could induce analogous expectancy to classic psychedelics while fortifying blinding. It would also be highly useful in understanding the neurochemistry of consciousness to use purported (Kehler and Lindskov, 2025) non-hallucinogenic psychedelics (e.g., 2-Br-LSD, Lewis et al, 2023; tabernanthalog, Cameron et al, 2021) for helping us understand how different receptors or even 2A-activated cellular signaling pathways affect cognitive functions. Many of the analyzed studies also had small samples, which can be especially problematic in psychedelic research due to the extreme idiosyncrasy of psychedelic experiences (Bălăeţ, 2022). Many of the participants were university students or part of an academic staff further limiting generalizability (see Tables 1–5). Many psychedelic effects are time-dependent and can follow oscillatory dynamics (Preller and Vollenweider, 2018). Future studies could benefit from measuring the temporal dynamics of aspects of psychedelic experiences (e.g., general intensity, mood alterations) with higher resolution, to better understand the fluctuations in various cognitive processes.
The majority of studies in our review researched the cognitive and psychological effects of psilocybin and LSD (Table 6). Only one study looked at the effects of ayahuasca (Uthaug et al., 2021). The scarcity of placebo-controlled studies on ayahuasca could be explained by the fact that these studies are often conducted in ceremonial settings with open-label design (dos Santos et al., 2016). DMT and ayahuasca are also generally less studied than psilocybin or LSD (Wen et al., 2024). The short duration of intravenous DMT can also complicate conducting cognitive tests during a stable level of alteration in consciousness further explaining the smaller number of studies. Although there have been some recent efforts looking at the effects of extended DMT (Luan et al., 2024). To gain a more complete picture of how consciousness can be pharmacologically altered, it is vital to also assess the cognitive effects of other classic psychedelics, DMT and mescaline. Furthermore, it would be especially enlightening to understand the effects of other synthetic serotonergic psychedelics with various pharmacological profiles (e.g., higher 5-HT2A-selectivity of TCB-2 and 25CN-NBOH or 5-HT1A-dominant 5-MeO-DMT) if their toxicity profile proves safe for human testing.
Conclusions
The recent resurgence in psychedelic research supports the potential of classic psychedelics to treat certain mental health disorders in controlled clinical settings (Andersen et al., 2020). However, their effects on cognitive and psychological functions remain less thoroughly explored. This review synthesized findings from 32 placebo-controlled studies investigating the acute and post-acute effects of psilocybin, DMT and LSD on empathy, memory, emotional processing, reaction time, attention, inhibition and cognitive flexibility. Psychedelics consistently enhanced emotional empathy, while effects on cognitive empathy were mixed or absent. Memory outcomes varied by task, dose and timing, ranging from impairments to no effect or improvement. Dose-dependent impairments were common in attention, reaction time and inhibition, though some studies reported no changes. Emotional processing was often disrupted, especially in response to negative stimuli. Evidence on cognitive flexibility was limited and inconclusive. Most studies assessed only acute effects and many had small sample sizes. Placebo effects—particularly challenging in psychedelic research—also remain a significant methodological concern. Future research should focus on developing specialized experimental paradigms for altered states, exploring how changes in cognitive and psychological functions relate to therapeutic benefits and integrating findings from trials with different active control conditions to build a more comprehensive understanding.
Supplemental Material
sj-docx-1-jop-10.1177_02698811251412012 – Supplemental material for Effects of LSD, DMT and psilocybin on cognitive and psychological functions: A systematic review of the literature
Supplemental material, sj-docx-1-jop-10.1177_02698811251412012 for Effects of LSD, DMT and psilocybin on cognitive and psychological functions: A systematic review of the literature by Marten Kase, Karl Kristjan Kaup and Jaan Aru in Journal of Psychopharmacology
Supplemental Material
sj-docx-2-jop-10.1177_02698811251412012 – Supplemental material for Effects of LSD, DMT and psilocybin on cognitive and psychological functions: A systematic review of the literature
Supplemental material, sj-docx-2-jop-10.1177_02698811251412012 for Effects of LSD, DMT and psilocybin on cognitive and psychological functions: A systematic review of the literature by Marten Kase, Karl Kristjan Kaup and Jaan Aru in Journal of Psychopharmacology
Footnotes
Acknowledgements
The author would like to thank Jaan Aru and Karl Kristjan Kaup for their valuable feedback and guidance throughout the preparation of this manuscript.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This work was supported by the Estonian Research Council grants PSG728 and Tem-TA 120, and the Estonian Centre of Excellence in Artificial Intelligence (EXAI), funded by the Estonian Ministry of Education and Research.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Declaration of generative AI and AI-assisted technologies in the writing process
During the preparation of this work the authors used ChatGPT in order to improve the readability of the text. After using this tool/service, the authors reviewed and edited the content as needed and take full responsibility for the content of the publication.
Supplemental material
Supplemental Material for this article is available online.