
Abstract
Classic psychedelics, including lysergic acid diethylamide (LSD), psilocybin, mescaline, N,N-dimethyltryptamine (DMT) and 5-methoxy-N,N-dimethyltryptamine (5-MeO-DMT), are potent psychoactive substances that have been studied for their physiological and psychological effects. However, our understanding of the potential interactions and outcomes when using these substances in combination with other drugs is limited. This systematic review aims to provide a comprehensive overview of the current research on drug–drug interactions between classic psychedelics and other drugs in humans. We conducted a thorough literature search using multiple databases, including PubMed, PsycINFO, Web of Science and other sources to supplement our search for relevant studies. A total of 7102 records were screened, and studies involving human data describing potential interactions (as well as the lack thereof) between classic psychedelics and other drugs were included. In total, we identified 52 studies from 36 reports published before September 2, 2023, encompassing 32 studies on LSD, 10 on psilocybin, 4 on mescaline, 3 on DMT, 2 on 5-MeO-DMT and 1 on ayahuasca. These studies provide insights into the interactions between classic psychedelics and a range of drugs, including antidepressants, antipsychotics, anxiolytics, mood stabilisers, recreational drugs and others. The findings revealed various effects when psychedelics were combined with other drugs, including both attenuated and potentiated effects, as well as instances where no changes were observed. Except for a few case reports, no serious adverse drug events were described in the included studies. An in-depth discussion of the results is presented, along with an exploration of the potential molecular pathways that underlie the observed effects.
Keywords
Introduction
Classic psychedelics include lysergic acid diethylamide (LSD), psilocybin (psilocin as an active agent), mescaline, N,N-dimethyltryptamine (DMT) and 5-methoxy-N,N-dimethyltryptamine (5-MeO-DMT) (Calvey and Howells, 2018). LSD has a high affinity for several serotonin (5-HT) receptors (such as 5-HT1A/B and 5-HT2A) and also has an affinity to dopaminergic D1–5 receptors (Halberstadt and Geyer, 2011). The affinity for 5-HT2A is important as the main mechanism behind the behavioural and psychological effects of LSD and other psychedelics is thought to be mediated through the activation of 5-HT2A receptors in cortical and subcortical structures (Vollenweider and Smallridge, 2022). LSD is metabolised by cytochrome P450 (CYP) enzymes such as CYP1A2, CYP3A4, CYP2C9, CYP2C19 and CYP2D6, with particular emphasis on the primary contribution of the former two (Wagmann et al., 2019).
While LSD is chemically synthesised, psilocybin is a naturally occurring psychedelic compound found in various genera of mushrooms, including Psilocybe, Panaeolus, Conocybe, Gymnopilus, Stropharia, Pluteus and Panaeolina (Dinis-Oliveira et al., 2019). Once ingested, the body metabolises psilocybin to psilocin, which is the primary psychoactive compound (Dinis-Oliveira, 2017). The metabolism of psilocybin involves several enzymes, including aldehyde dehydrogenase, monoamine oxidase (MAO) and UDP-glucuronosyltransferase (UGT) enzymes such as UGT1A9, UGT1A10, UGT1A6, UGT1A7 and UGT1A8 (Dinis-Oliveira, 2017). Similar to LSD, psilocin acts as an agonist at 5-HT2A receptors in the brain (Rickli et al., 2016), where it exerts its psychological effects (F. X. Vollenweider et al., 1998).
Mescaline (3,4,5-trimethoxyphenethylamine) is a psychedelic compound found in the North American peyote cactus (Lophophora williamsii), the South American San Pedro cactus (Echinopsis pachanoi), as well as other cacti such as the Peruvian torch cactus (Echinopsis peruviana), Bolivian torch cactus (Echinopsis lageniformis) and the leaf cactus (Pereskia aculeata) (Dinis-Oliveira et al., 2019). It has a high affinity for 5-HT1A and 5-HT2A/C receptors but is less potent than LSD, psilocin and DMT (Rickli et al., 2016). Mescaline is metabolised in the liver and broken down into several inactive compounds, with oxidative deamination occurring via MAO or diamine oxidase (Dinis-Oliveira et al., 2019).
DMT is a naturally occurring psychoactive compound found in several plants and is also endogenously produced in mammals, including humans (Jiménez and Bouso, 2022). Recreationally, DMT is consumed either in a pure form (mostly smoked) or as a key ingredient in an orally active brew called ayahuasca (Cakic et al., 2010). Ayahuasca is made by mixing a DMT-containing plant with a vine containing β-carboline alkaloids (Brito-da-Costa et al., 2020). Although the exact plant combinations can vary, a frequent mixture is made of Psychotria viridis (source of DMT) and Banisteriopsis caapi vine, whose stem and bark contain β-carbolines harmine and harmaline (Brito-da-Costa et al., 2020).
DMT is a partial agonist primarily of the 5-HT1A, 5-HT2A and 5-HT2C receptors (Carbonaro and Gatch, 2016). Oral consumption of DMT does not produce psychotropic effects due to rapid metabolism by MAO enzymes (Riba et al., 2015). However, when consumed orally as part of the ayahuasca brew, DMT becomes bioavailable due to the MAO-A inhibiting effects of harmine and harmaline, which protect DMT from deamination in the gut (Brito-da-Costa et al., 2020). DMT alone has a short half-life of 9–12 min and is rapidly metabolised by MAO-A, while CYP2D6 and, to a lesser extent, CYP2C19 are also involved in its metabolism (Good et al., 2022).
Finally, 5-MeO-DMT is a psychedelic that has been detected in numerous plant and fungal sources, as well as in the gland secretions of the Incilius alvarius toad (Ermakova et al., 2022). 5-MeO-DMT is a non-selective serotonin receptor agonist, exhibiting a strong affinity for 5-HT1A/1B/1D/6/7 but a significantly lower affinity for 5-HT2A receptor subtype (Halberstadt et al., 2012; Ray, 2010). Both receptors, 5-HT1A and 5-HT2A activation, are involved in 5-MeO-DMT behavioural effects (Ermakova et al., 2022). 5-MeO-DMT has been observed to robustly inhibit the reuptake of serotonin but also dopamine and norepinephrine while demonstrating minimal activity in terms of releasing these three compounds (Nagai et al., 2007). The primary process by which 5-MeO-DMT is metabolised involves oxidative deamination catalysed by MAO-A, leading to the formation of 5-methoxyindoleacetic acid. In addition, a minor portion of 5-MeO-DMT undergoes O-demethylation via CYP2D6 to produce bufotenine (5-hydroxy-DMT) (Halberstadt, 2016) which is a potent 5-HT2A agonist (Egan et al., 2000).
Drug–drug interactions (DDIs) can be categorised as either pharmacokinetic or pharmacodynamic interactions. Pharmacokinetic interactions occur when one drug influences the absorption, distribution, metabolism or elimination of another drug. On the other hand, pharmacodynamic interactions involve the modification of the pharmacological effect of one drug by another. These interactions can exhibit synergistic, additive or antagonistic characteristics. Additivity refers to the overall effect of a drug combination which is the sum of the effects of each individual drug, while synergy occurs when the combined effect of the drugs is greater than additive. Antagonism arises when the combined effect is less than additive (Niu et al., 2019).
One common mechanism of pharmacodynamic drug interaction is competition at the receptor level. When two drugs interact with the same receptor, they can compete for binding, leading to alterations in their pharmacological effects (Lambert, 2004). For instance, blocking the receptors where LSD, psilocin, mescaline or DMT exert their effects, such as 5-HT2A, could impede their psychological effects.
An example of a pharmacokinetic DDI is the inhibition of drug-metabolising enzymes, such as cytochrome P450, which are responsible for metabolising a broad range of drugs (Zhao et al., 2021). Inhibition of these enzymes by concomitant drugs or circulating metabolites can lead to altered drug metabolism and impact the drug’s effects and influence treatment outcomes (Zhao et al., 2021). Additionally, there is the potential for interaction with P-glycoprotein (P-gp), a membrane transporter that facilitates the efflux of various drugs and is present in the kidneys, liver, gastrointestinal tract and blood–brain barrier (Amin, 2013). Similar to CYP enzymes, reducing the activity of P-gp can increase the concentration of its substrates in the blood, whereas increasing its activity can decrease the concentration, leading to inadequate therapeutic effects (Amin, 2013).
For instance, CYP enzymes have a known role in LSD metabolism (Luethi et al., 2019) and therefore can affect LSD’s effects (Straumann et al., 2023; Vizeli et al., 2021). DDIs can occur even when drugs are not taken concurrently, allowing for days or even weeks between their administration. Some drugs, like fluoxetine, have prolonged inhibitory effects on CYP activity that may persist for several weeks following its discontinuation due to the extended half-life of fluoxetine and its metabolite norfluoxetine (Hemeryck and Belpaire, 2002).
Currently, there is limited literature available on the DDIs between classic psychedelics and other drugs. While two review articles have been published on this subject, the first one was limited in scope (Wyatt et al., 1976) and the second one focused solely on 3,4-methylenedioxymethamphetamine (MDMA) and psilocybin interactions with psychiatric medications (Sarparast et al., 2022). To fill this gap, this systematic review aimed to provide a comprehensive overview of the current state of research on DDIs between classic psychedelics and any other drugs. We conducted a thorough literature search and reported on both physiological and subjective outcomes. This review offers valuable insights into the potential risks and benefits of combining classic psychedelics (LSD, psilocybin, mescaline, DMT and 5-MeO-DMT) with other drugs, thereby guiding researchers and clinicians in this field.
Methods
This review was registered in PROSPERO (CRD42022336092) and followed the latest Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) (2020) guidelines (Page et al., 2021). A keyword search for articles pertaining to classical psychedelics (LSD, psilocybin, mescaline, DMT and ayahuasca) was initially conducted on June 5, 2022, in three primary scientific databases: PubMed, PsycINFO and Web of Science (with no year restriction). These databases comprise a general journal articles database and a database specialised in biomedicine and psychology research. Search terms included keywords (including synonyms) related to classic psychedelics in the title, abstract, keywords, full text (where available) and MeSH terms, as well as keywords related to drug interactions, side effects and adverse reactions. The search was not limited by the time period to capture all relevant articles. A total of 2151 articles were identified during the first search. The search was repeated on September 2, 2023, resulting in the identification of 262 additional articles published since the initial search. The full search terms for each database are provided in the Supplemental Material S1. Furthermore, on September 2, 2023, an additional search was carried out using the same search parameters to identify articles detailing drug interactions involving 5-MeO-DMT (full search terms are provided in Supplemental Material S2). A total of 507 records relating to 5-MeO-DMT were found.
Additionally, to ensure comprehensive coverage, a search was conducted in the Multidisciplinary Association for Psychedelic Studies (MAPS) comprehensive online ‘Psychedelic Bibliography’ database (https://bibliography.maps.org), which contains scientific but also non-scientific articles specifically about psychedelics, dating back to 1841. At the time of the search (November 20, 2022), the database contained 13,237 records. All records were downloaded, followed by keyword searches in all titles and abstracts (targeting ‘LSD’, ‘psilocybin’, ‘mescaline’, ‘DMT’ and ‘ayahuasca’, including synonyms and variations), resulting in a total of 6336 records.
Furthermore, we manually searched a registry of clinical trials at ClinicalTrials.gov on April 20, 2023, to find any articles containing results from clinical trials that were missed in the database search and conducted reference lists checking of included studies to find additional missing articles. An additional search of ClinicalTrials.gov was conducted on September 2, 2023 to find clinical trials involving 5-MeO-DMT.
Records were screened in three phases: first, those found via a scientific database search; second, those from the MAPS bibliography database and finally, records from a search for 5-MeO-DMT. In all cases, a systematic review application Catchii (Halman and Oshlack, 2023) was used. Duplicate removal was performed using Catchii’s duplicate detection method and the results were manually verified before removal. Two researchers independently conducted title and abstract screening, both of whom were blinded to each other’s decisions. The inclusion criteria were as follows: (1) any article on human participants, (2) any article describing the usage of any classical psychedelics with another drug, (3) studies describing physiological and/or psychological effects of classical psychedelics with another drug (or the lack of), (4) studies that were either randomised controlled trials, observational studies (cohort, case–control, cross-sectional) or case reports/studies, and (5) studies published in English. Disagreements between reviewers were discussed until a consensus decision was reached. Records that did not meet the population, intervention and outcome criteria were excluded. After removing duplicates, a total of 7102 records (1667 from the first phase, 5065 from the second phase and 370 from the third phase) underwent the first stage of screening, which involved title and abstract analysis. Subsequently, the full text of the 77 reports that passed this stage was assessed in the second stage, including articles identified from citation searches. Due to the small number of records retrieved from the updated scientific database search, each title and abstract was assessed directly within the search results. The summary of results is shown on the PRISMA flow diagram (Figure 1). Data from all eligible records were extracted by the authors of this review, who corroborated each other’s findings. Outcome measures included physiological and psychological outcomes.
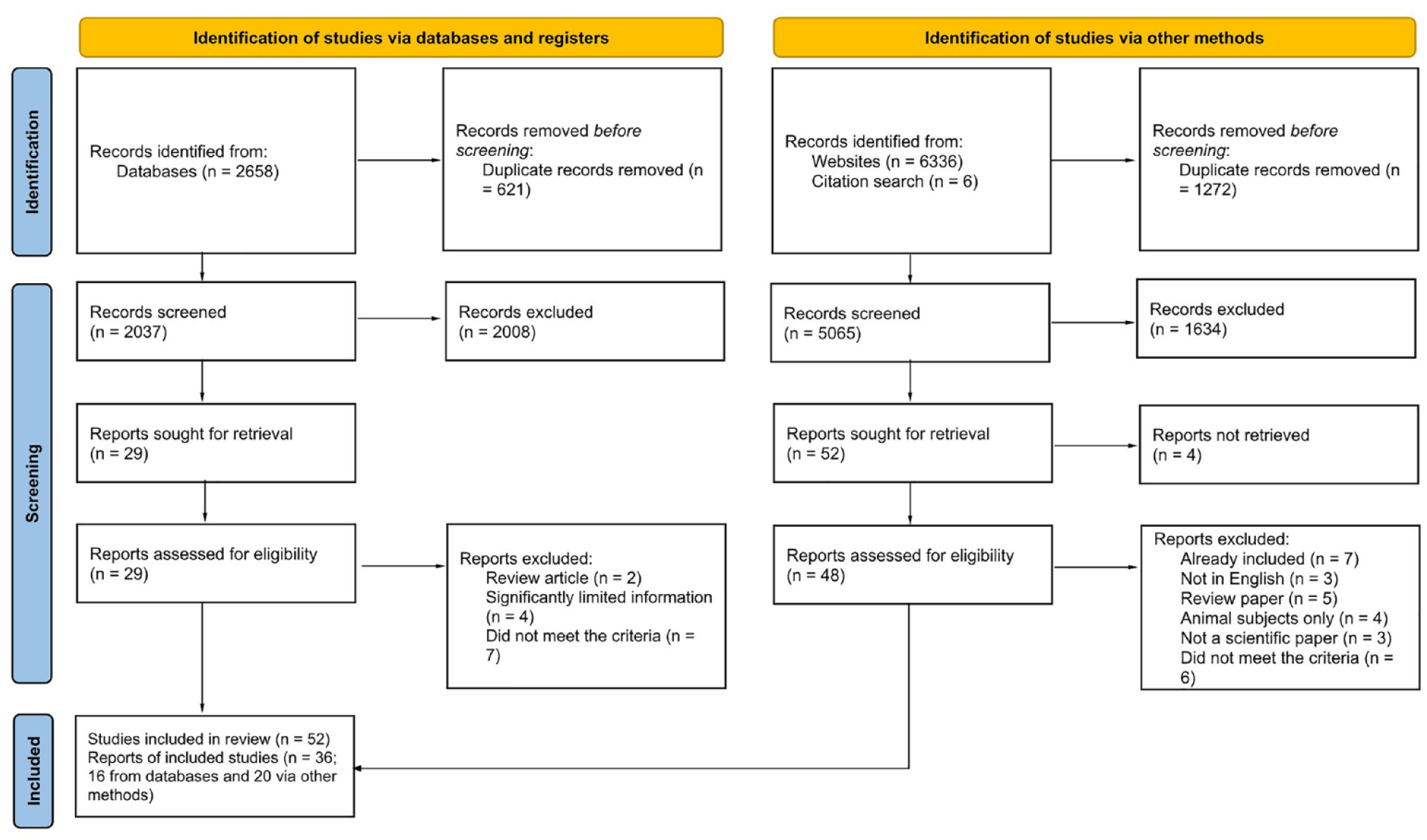
PRISMA flow chart depicting the identification of studies in the systematic review process.
Results
The results of this systematic review include studies investigating DDIs between psychedelic drugs (LSD, psilocybin, mescaline, 5-MeO-DMT, DMT and ayahuasca) and other drugs. Regarding LSD, the review examined interactions with antipsychotics (chlorpromazine), mood stabilisers (lithium), various antidepressants including selective serotonin reuptake inhibitors (SSRIs; fluoxetine, sertraline, paroxetine and trazodone), tricyclic antidepressants (TCAs; imipramine, desipramine and clomipramine), as well as monoamine oxidase inhibitors (MAOIs; phenelzine, isocarboxazid, nialamide and iproniazid) and other substances (azacyclonol). Recreational substances (alcohol and MDMA), as well as other substances such as ketanserin, reserpine, niacin, scopolamine and phenoxybenzamine, were also included.
Regarding psilocybin, the review contains reports on its interactions with anxiolytics (buspirone), antipsychotics (chlorpromazine, haloperidol and risperidone), SSRI antidepressants (escitalopram) and recreational drugs (alcohol). Other investigated substances included ketanserin and ergotamine. In the case of mescaline, the review examined interactions with antipsychotics (chlorpromazine and promazine), antidepressants (azacyclonol) and the compound 2,4,5-trimethoxyphenethylamine (2C-O). For DMT, the review explored interactions with MAOI antidepressants (iproniazid), as well as other substances like racemic pindolol and methysergide. Lastly, one report of the SSRI antidepressant fluoxetine was found for ayahuasca and two case reports for 5-MeO-DMT involving the use of β-carbolines.
Results of studies included in this review are provided as separate tables for LSD (Table 1), psilocybin (Table 2), mescaline (Table 3), DMT together with ayahuasca (Table 4) and 5-MeO-DMT (Table 5), grouped by drug class. More detailed narrative descriptions of the results from each study are shown in Supplemental Material S3. Additionally, a combined table (Supplemental Table S1) summarises all the results from Tables 1 to 5, along with listing the potentially relevant molecular targets and actions of each drug.
Summary of studies and case reports describing interactions (or the absence thereof) between LSD and other drugs.
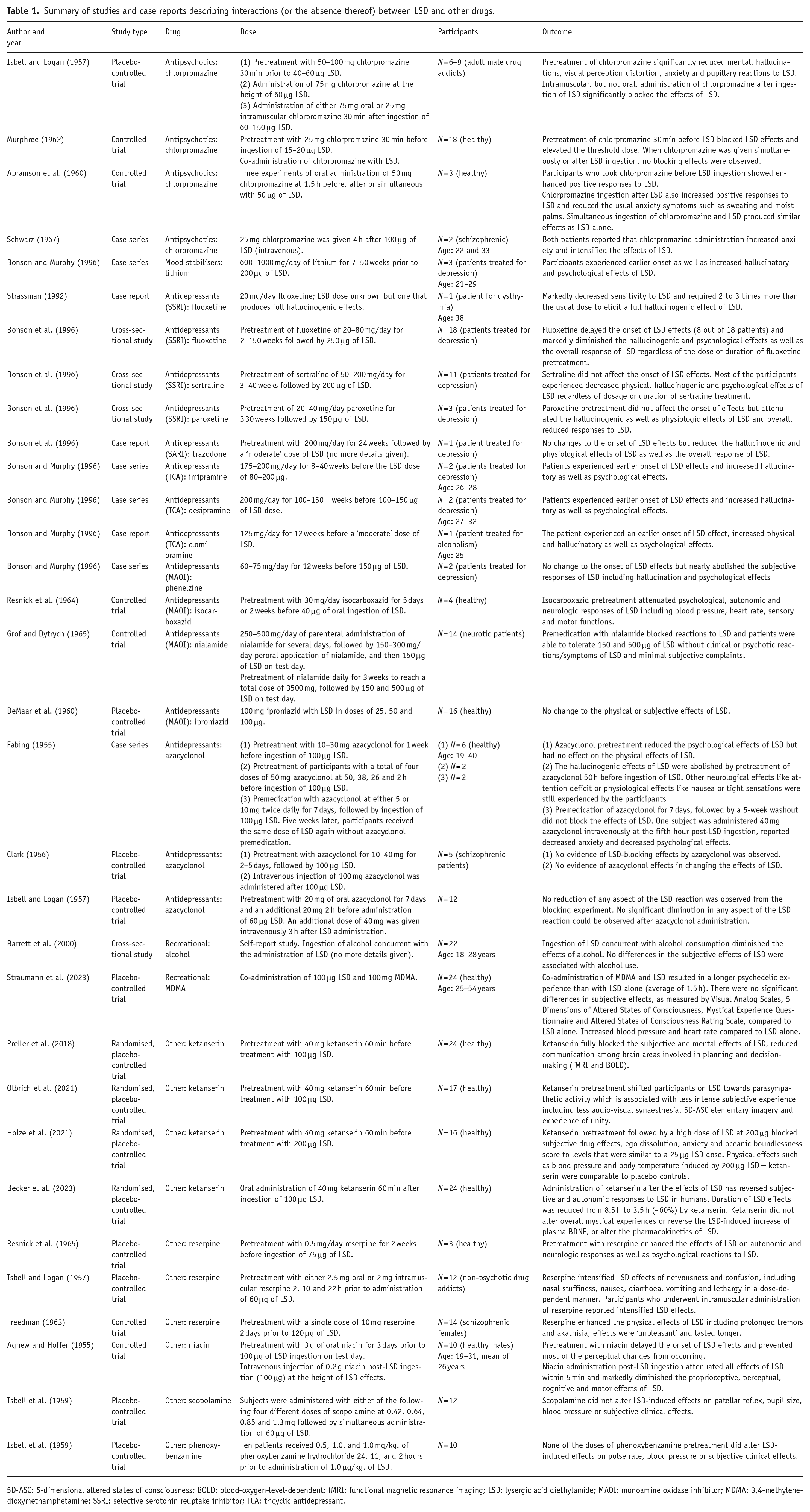
5D-ASC: 5-dimensional altered states of consciousness; BOLD: blood-oxygen-level-dependent; fMRI: functional magnetic resonance imaging; LSD: lysergic acid diethylamide; MAOI: monoamine oxidase inhibitor; MDMA: 3,4-methylenedioxymethamphetamine; SSRI: selective serotonin reuptake inhibitor; TCA: tricyclic antidepressant.
Summary of studies and case reports describing interactions (or the absence thereof) between psilocybin and other drugs.
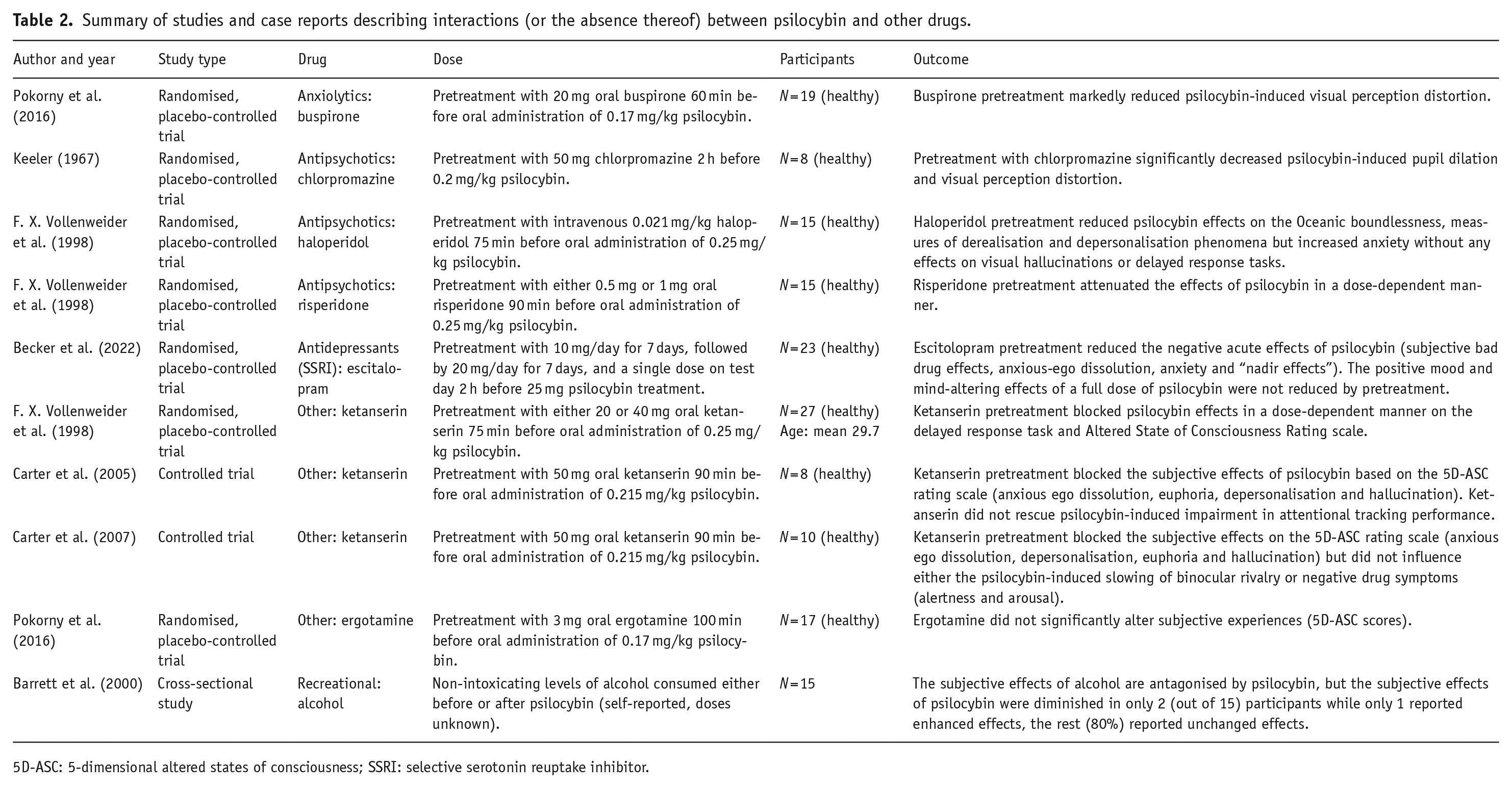
5D-ASC: 5-dimensional altered states of consciousness; SSRI: selective serotonin reuptake inhibitor.
Summary of studies and case reports describing interactions (or the absence thereof) between mescaline and other drugs.
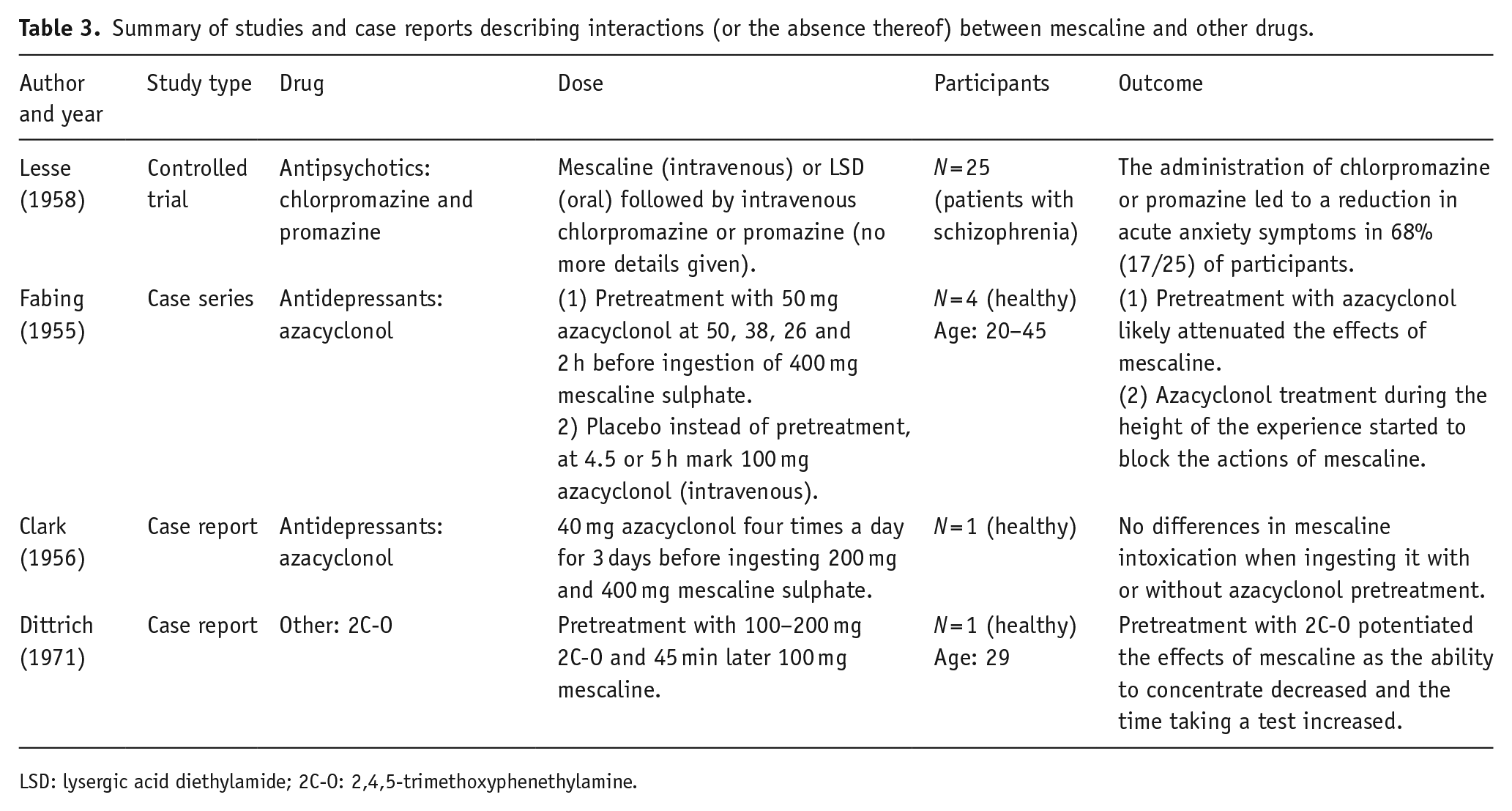
LSD: lysergic acid diethylamide; 2C-O: 2,4,5-trimethoxyphenethylamine.
Summary of studies and case reports describing interactions (or the absence thereof) between DMT, ayahuasca and other drugs.
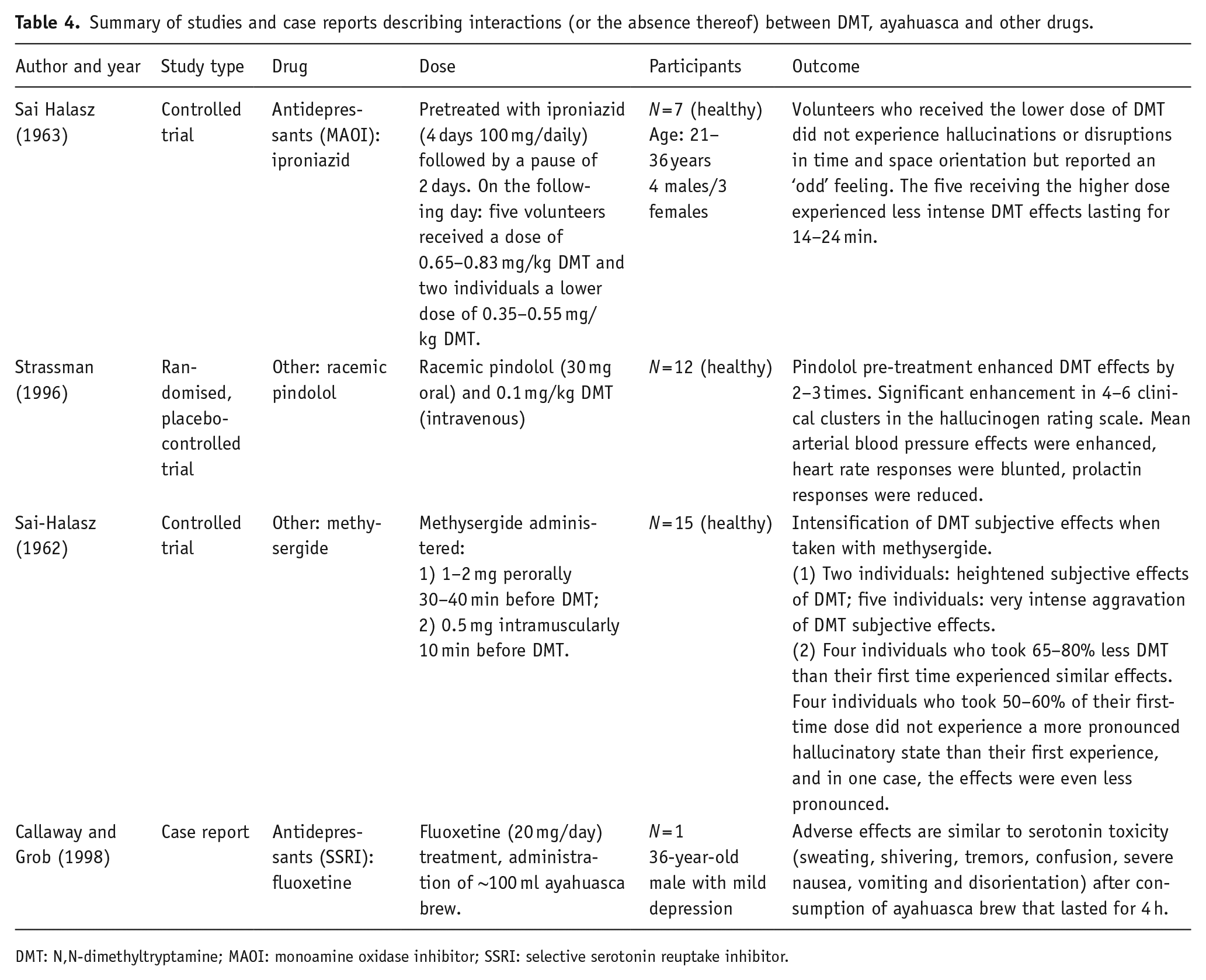
DMT: N,N-dimethyltryptamine; MAOI: monoamine oxidase inhibitor; SSRI: selective serotonin reuptake inhibitor.
Summary of studies and case reports describing interactions (or the absence thereof) between 5-MeO-DMT and other drugs.
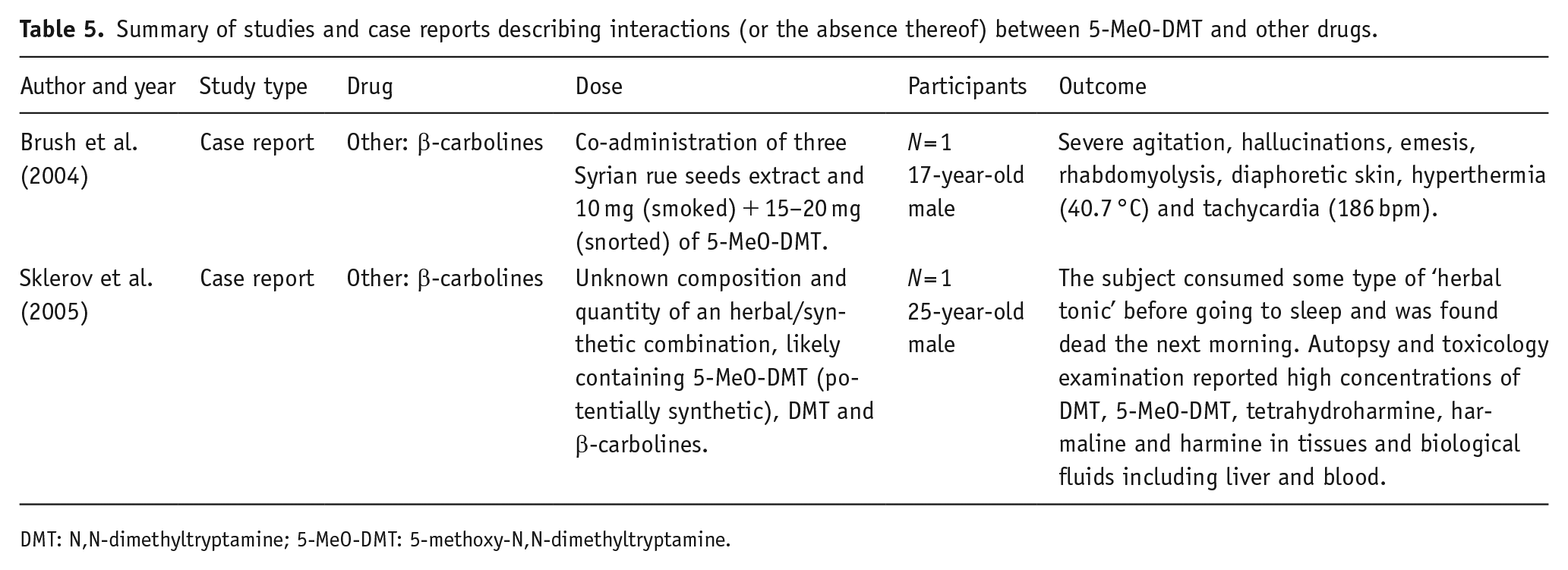
DMT: N,N-dimethyltryptamine; 5-MeO-DMT: 5-methoxy-N,N-dimethyltryptamine.
Discussion
In this systematic review, a total of 7102 records were screened and 36 reports from 52 studies were included, providing information about the potential DDIs (or the lack of) of classic psychedelics in the scientific literature. Considering the scarcity of studies addressing DDIs involving classic psychedelics, case reports were included to provide more insights into DDIs. Although anecdotal in nature, these case reports highlight potential DDIs and areas for future research. In the following sections, all drugs are discussed, grouped by their properties and/or class, where possible. Groups with blocking or reduced effects on psychedelics are generally addressed first, followed by those with potentiating effects, and concluding with those that have mixed evidence or no effects.
Serotonin 2A receptor antagonists
Classic psychedelics are known to bind to 5-HT2A receptors where it is thought that their hallucinogenic properties are mediated (Vollenweider and Smallridge, 2022). The results of multiple studies on drugs that are 5-HT2A receptor antagonists, such as ketanserin, trazodone, risperidone and chlorpromazine, indicated that they were all effective in attenuating the effects of LSD and psilocybin. Specifically, ketanserin, a 5-HT2A antagonist with a strong affinity (Becker et al., 2023) was found in all studies to either fully block or reduce the subjective effects of LSD (Becker et al., 2023; Holze et al., 2021; Olbrich et al., 2021; Preller et al., 2018) and psilocybin (Carter et al., 2005, 2007; F. X. Vollenweider et al., 1998). Similarly, decreased physical effects were also observed, including lowered blood pressure, body temperature and heart rate (Holze et al., 2021). Similarly,
The findings from studies using chlorpromazine, which is also a 5-HT2A receptor antagonist but has an affinity for D2 and adrenoceptor α1A and α1B subtypes as well (Boyd-Kimball et al., 2019; Gillman, 1999), are inconsistent. While two studies (Isbell and Logan, 1957; Murphree, 1962) found that chlorpromazine reduced the intensity of LSD-induced effects as well as anxiety, another two independent studies (Abramson et al., 1960; Schwarz, 1967) reported the opposite. In addition, while Isbell and Logan (1957) reported that oral pre-administration of chlorpromazine blocked the effects of LSD and not post-administration, then Abramson et al. (1960) observed enhanced effects regardless of whether it was administered before or after LSD intake. However, the inconsistent findings regarding the effects of chlorpromazine on LSD response may be due to differences in study design, dosing regimens and individual variability. Moreover, chlorpromazine was reported to attenuate the effects of psilocybin and mescaline, where its pretreatment significantly decreased psilocybin-induced visual perception distortion (Keeler, 1967) and abolished the increased anxiety observed in schizophrenic patients following mescaline ingestion (Lesse, 1958). Overall, considering the available evidence, treatment with potent 5-HT2A antagonists, including medicines, is anticipated to diminish the effects of psychedelics (e.g. antipsychotics risperidone, olanzapine and pipamperone, antidepressants mirtazapine, mianserin and etoperidone as well as migraine prevention drug pizotifen).
Serotonin 1A receptor agonists
Pretreatment with buspirone, which is a 5-HT1A partial agonist, markedly reduced the psilocybin-induced visual hallucinations (Pokorny et al., 2016). On the contrary, agonism of 5-HT1A with ergotamine did not affect any of the psilocybin effects (Pokorny et al., 2016). However, given that ergotamine has lower efficacy at receptor signal transduction when compared to buspirone and its generally low bioavailability, it was suggested that the administered dose may not have been high enough to compete with psilocybin at those receptor site (Pokorny et al., 2016). These findings indicate that 5-HT1A receptors may be also involved in the manifestation of psilocybin-induced effects such as visual hallucinations, affective alterations, derealisation and depersonalisation. The mechanism of action by which buspirone reduces hallucinations may involve direct stimulation of 5-HT1A receptors, or alternatively, interaction between 5-HT1A and 5-HT2A receptors on pyramidal cells due to co-expression of 5-HT1A and 5-HT2A receptors in cortical and visual areas (Pokorny et al., 2016). Despite both psilocin and buspirone being partial agonists at 5-HT1A receptors, the blocking effect of buspirone may be due to a more effective inhibitory impact on pyramidal neurons compared to psilocin (Pokorny et al., 2016).
Dopamine receptor antagonists
Buspirone is also an antagonist for dopamine D2 receptors (low affinity) and has shown weak affinity to the 5-HT2 receptors (Loane and Politis, 2012), which could have a role in modulating the effects of psilocybin as well. However, while LSD is known to have an effect on the dopaminergic system (found to be important in perceived selfhood and cognition) (Lawn et al., 2022) and it has a high affinity for both D1 and D2 receptors (Halberstadt and Geyer, 2011), then psilocin does not bind to dopamine receptors (Halberstadt and Geyer, 2011). Moreover, D2 antagonism by haloperidol alone did not have an effect on psilocybin-induced hallucinations (F. X. Vollenweider et al., 1998). Haloperidol did, however, diminish the feelings of oceanic boundlessness and derealisation and also increased anxiety (F. X. Vollenweider et al., 1998). In this case, psilocybin may exert an indirect impact on dopaminergic systems, which is subsequently counteracted by haloperidol (F. X. Vollenweider et al., 1998). It has been shown in rats that psilocybin administration can increase extracellular dopamine levels in the frontal cortex (Wojtas et al., 2022) and psilocin administration has also increased dopamine levels in the nucleus accumbens (Sakashita et al., 2015).
Serotonin reuptake inhibitors
Interestingly, the blocking of serotonin reuptake transporters with SSRIs, such as fluoxetine, sertraline and paroxetine, reduced the effects of LSD as reported in one study (Bonson et al., 1996). Moreover, fluoxetine also delayed the onset of LSD effects in nearly half of the participants in the same study (Bonson et al., 1996) and was shown to markedly decrease sensitivity to LSD in one case report (Strassman, 1992). SSRIs can also result in DDI at the pharmacokinetic level as some of them, such as fluoxetine, are potent inhibitors of CYP2D6 enzymes (Brøsen, 1998), which can modulate the effects of psychedelics that are being metabolised by such enzymes. For example, CYP2D6 enzymatic activity is already known to influence the effects of LSD (Straumann et al., 2023; Vizeli et al., 2021). Considering this, one might have expected a stronger and more prolonged response from LSD instead of reduced effects. However, ultimately this could involve an interplay between various molecular actions, including interactions with enzymes, receptor availability and other molecular factors.
Trazodone, which was previously described to reduce LSD effects as it is a 5-HT2A antagonist, is also a serotonin receptor antagonist and reuptake inhibitor (SARI) (Fagiolini et al., 2012) that might have a role in the observed effects as well. Potentially, SSRIs and SARIs that increase extracellular serotonin by inhibiting its uptake can then attenuate the effects of psychedelics via receptor competition with the endogenous serotonin. In addition, it has been shown that repeated administration of SSRIs desensitises 5-HT2A receptors which may reduce the cell’s response to psychedelics by binding to these receptors (Gray and Roth, 2001). Chronic SSRI use can also increase serotonin release via desensitisation of 5-HT1A raphe autoreceptors (Artigas et al., 2001).
However, one SSRI, escitalopram, did not have an effect on psilocybin-induced positive mood or mind-altering effects (depersonalisation, oceanic boundlessness or euphoria), but it did reduce ego disintegration and anxiety (Becker et al., 2022). The authors hypothesised that while LSD does not affect the serotonin transporter, psilocin has a weak inhibitory effect on it (Becker et al., 2022; Rickli et al., 2016). This distinction in pharmacology could lead to different interactions between antidepressants and LSD compared to psilocybin (Becker et al., 2022), explaining the differences in response. Although this study offers important insights into the safety of administering psilocybin to patients taking escitalopram, it is crucial to bear in mind that the treatment phase of this study was 14 days. Longer treatment periods with escitalopram, which may often last for years, could lead to distinct outcomes due to molecular changes that occur over extended periods (Faure et al., 2006).
Nonetheless, another study (not included in the results) involved 19 patients with treatment-resistant depression and who were on chronic SSRI treatment (sertraline, escitalopram, fluoxetine, vilazodone, paroxetine or citalopram). The patients were treated with synthetic psilocybin (an investigational drug named ‘COMP360’) and no serious treatment-emergent adverse events were reported (Goodwin et al., 2023). However, two cases were considered severe (both blood pressure increase) and necessitated treatment with clonidine, but the study did not provide information regarding which specific SSRI treatments these individuals were on. Secondly, although the study measured 5-dimensional altered states of consciousness (5D-ASC) scores, it also did not provide a breakdown by specific SSRIs. This absence of information makes it difficult to assess and compare the individual impacts of different SSRIs on subjective effects as well, particularly given the high standard deviations in the 5D-ASC results and instances of zero scores in three dimensions, indicating some participants did not respond to psilocybin. Improvements in depression severity were observed in 42.1% of cases at week 3, which indicates that ongoing SSRI treatment did not significantly hinder psilocybin’s therapeutic potential, at least for those participants. Based on the results, the authors hypothesise that chronic SSRI treatment may not significantly downregulate 5-HT2A/C receptors, or potentially indicate a lesser impact on the downregulated receptors on the psychedelic experience than previously theorised. It is also possible that unknown mechanisms downstream of 5-HT2A/C receptor signalling could compensate for any potential effects of chronic SSRI treatment on the psychedelic response, as they state (Goodwin et al., 2023). While the study contributes to the body of evidence supporting the safety of using psilocybin alongside SSRI treatment, it is important to note its limitations, including a small sample size and the absence of a breakdown of specific SSRIs. This latter is crucial for linking adverse events and subjective effects to individual drugs, given their distinct properties and potential for varied drug interactions. Due to this, this article was not included in the main results.
Another study by Gukasyan et al. (2023), based on online retrospective survey results, also provided generalised findings for SSRIs and serotonin and norepinephrine reuptake inhibitors (SNRIs) (thus not included in the primary results). This study reported weakened effects of psilocybin in approximately half of the participants when used concurrently with SSRI/SNRIs, also suggesting downregulation of 5-HT2A receptors (Gukasyan et al., 2023).
Monoamine oxidase inhibitors
Results from another antidepressant class, MAOIs, showed a similar outcome to those of SSRIs. In particular, phenelzine, isocarboxazid, nialamide and iproniazid are all non-selective MAO inhibitors (Entzeroth and Ratty, 2017), therefore inhibiting both MAO-A and MAO-B enzymes. The first three listed antidepressants attenuated or blocked the effects of LSD (Bonson and Murphy, 1996; Grof and Dytrych, 1965; Resnick et al., 1964). By contrast, the latter, iproniazid, did not alter the subjective or physical effects of LSD (DeMaar et al., 1960). However, iproniazid was also used in the experiments with DMT where it was shown to reduce the effects of DMT (Sai Halasz, 1963). This particular finding is interesting as iproniazid is an irreversible MAOI (Entzeroth and Ratty, 2017) that inhibits the enzyme that rapidly metabolises DMT (Barker et al., 1980). Moreover, studies in rats have shown that iproniazid pretreatment increases levels of DMT in the brain, as well as in the liver, kidney and blood (Sitaram et al., 1987). However, the reduced DMT effects could be due to increased serotonin levels after blocking MAO and it is hypothesised that higher doses of DMT are needed when serotonin levels are elevated (Sai Halasz, 1963). It is suggested that ayahuasca’s effect is mediated by MAO inhibition in the digestive system or bloodstream, which protects DMT from metabolism during its transit to the brain, where MAO inhibitors can then attenuate DMT’s effects due to elevated brain serotonin (Ott, 1996). This hypothesis for DMT can also explain the similar, attenuated response, which was observed in the studies where LSD was combined with MAOIs (Bonson and Murphy, 1996; Grof and Dytrych, 1965; Resnick et al., 1964).
In addition to metabolising DMT, MAOIs are also important in the breakdown of serotonin as they block MAO enzymes involved in its metabolism (Foong et al., 2018). This gives rise to a possible scenario of excessive serotonin levels in the brain. In particular, when combining MAOIs with each other or with SSRI/SNRI, it has been thought to carry the greatest risk of serotonin toxicity (Foong et al., 2018). In one case report, symptoms resembling serotonin toxicity were reported after an individual was administered ayahuasca while being on a fluoxetine treatment (Callaway and Grob, 1998). However, the patient also recovered rapidly within 4 h after the administration of ayahuasca without any treatment. Moreover, another case report that was not included in the review documented an instance of serotonin toxicity in an individual who had been undergoing
Ayahuasca brew is usually made from a plant containing DMT and another plant that contains β-carbolines which mostly inhibit MAO-A, thereby making DMT orally active (McKenna, 2004). Tetrahydroharmine (THH), one of the β-carbolines present in the brew, can also act as a weak serotonin uptake inhibitor and increase brain serotonin levels (Airaksinen et al., 1980). Fluoxetine strongly inhibits the CYP2D6 enzyme, which metabolises many drugs, including serotonin. The concurrent inhibition of serotonin reuptake and serotonin-metabolising enzymes, such as CYP2D6 and MAO, can cause an accumulation of serotonin in the brain and low clearance, potentially leading to life-threatening serotonin toxicity (Dunkley et al., 2003). This explanation aligns with the reported case reports; however, further research is necessary to confirm the potential association. While more evidence is required to shed light on the possible DDI, it would be advisable to exercise caution when combining MAOIs found in ayahuasca with SSRIs and SNRIs (e.g. citalopram, escitalopram, fluvoxamine, fluoxetine, paroxetine and sertraline).
There are two case reports in which the consumption of 5-MeO-DMT alongside β-carbolines (MAO-A inhibitors) resulted in a serious adverse event. In one case (Brush et al., 2004), the ingestion of 5-MeO-DMT in combination with three Syrian rue (Peganum harmala) seeds containing harmaline and harmine led to severe agitation, hallucinations, emesis, rhabdomyolysis, diaphoretic skin, hyperthermia and tachycardia. Based on the observed time course and symptoms, the authors suggested the possibility of MAOI poisoning (Brush et al., 2004). Alternatively, they also considered serotonin toxicity but deemed it less likely. A critical assessment of the report by dos Santos (2013) suggested serotonin toxicity over MAOI poisoning (dos Santos, 2013).
In this case report, it was indicated that three Syrian rue seeds were consumed. However, considering the tiny weight (Li et al., 2023) and the average content of alkaloids in these seeds (Herraiz et al., 2010), the estimated total harmaline and harmine content would be only approximately 0.26 mg and 0.34 mg, respectively. The combined amount is at least 50 times less than the same alkaloid levels typically used in a standard ayahuasca brew for MAO inhibition (D. J. McKenna et al., 1984; Rivier and Lindgren, 1972). While it is challenging to determine the minimum dose of harmaline/harmine necessary for MAO inhibition that induces such adverse effects, it is important to approach this reported consumption of three seeds with a degree of scepticism. It might be possible that the individual may have instead consumed seed capsules, each containing over 50 seeds (Moloudizargari et al., 2013), totalling approximately over 30 mg of combined harmaline and harmine for three capsules, comparable to doses used in an ayahuasca brew.
The second case report (Sklerov et al., 2005) described the death of a 25-year-old male after he consumed herbal extracts containing β-carbolines and tryptamines. No information is available regarding the composition and dosage of the products but toxicology analysis revealed the presence of 5-MeO-DMT, DMT, MAOIs (harmaline and harmine) and THH (weak serotonin uptake inhibitor). This particular report has faced some criticism for associating toxicity with ayahuasca and for not including several important details (Callaway et al., 2006). Callaway and his colleagues suggested that the elevated levels of 5-MeO-DMT found in the deceased individual’s blood likely have a synthetic origin rather than originating from an ayahuasca brew. This conclusion arises because common ingredients in ayahuasca brew do not contain 5-MeO-DMT or, at most, contain only trace amounts of it (Callaway et al., 2006).
The toxicity observed in those two case reports remains unclear, but could also have arisen from the impact of MAOIs on the levels of 5-MeO-DMT in the blood and brain as hypothesised by Halberstadt (2016). Inhibiting MAO-A activity by β-carbolines could substantially increase concentrations and accumulation of 5-MeO-DMT in the brain and may also lead to interactions with various other targets, potentially giving rise to adverse effects and toxicity (Halberstadt, 2016; Jiang et al., 2013). Further evidence is required to confirm the link between 5-MeO-DMT and MAO-A inhibitors. However, based on the case reports, one should exercise caution when combining 5-MeO-DMT with either MAO-A inhibitors (such as harmaline and harmine) or non-selective MAO inhibitors (e.g. medicines such as tranylcypromine, isocarboxazid, phenelzine and selegiline).
Other drugs (blocking action)
One study that reported a blocking action on LSD effects was using a high dose of niacin (vitamin B3). Pretreatment with niacin resulted in delayed onset of LSD effects and prevented most of the perceptual changes from occurring, and administration post-LSD ingestion attenuated all effects of LSD within 5 min (Agnew and Hoffer, 1955). The blocking mechanism of niacin is not clear but may be due to increased serotonin levels. This is indicated by the studies where niacin has been shown to induce serotonin release from human platelets within a few minutes and to increase plasma serotonin levels in rats (Papaliodis et al., 2008). Secondly, blocking 5-HT2A receptors with ketanserin inhibits niacin-induced temperature increase (Papaliodis et al., 2008), which indicates that serotonin is involved in the effects of niacin. Finally, high-dose treatment with niacin has caused manic-like psychotic episodes that were thought to occur via stimulation of serotonin but also dopamine production (Loebl and Raskin, 2013).
Tricyclic antidepressants
Several studies have reported the potentiated effects of LSD. In one study, desipramine, imipramine and clomipramine, which are all TCAs, were shown to increase the psychological effects of LSD (Bonson and Murphy, 1996). Desipramine is a potent inhibitor of noradrenaline reuptake, while imipramine and clomipramine exhibit a lower degree of noradrenaline and serotonin reuptake inhibition (Gillman, 2007). Furthermore, clomipramine also functions as a 5-HT2A receptor antagonist and also imipramine has an affinity for this receptor subtype (Gillman, 2007).
Since TCAs were reported to enhance the effects of LSD despite some of them inhibiting serotonin uptake and are even 5-HT2A receptor antagonists (similar to SSRIs that decreased the effects of LSD), it is suggested another mechanism can contribute. For instance, chronic administration of TCAs, such as desipramine, can increase the sensitivity of certain neurons to LSD, suggesting that these medications may sensitise postsynaptic serotonin receptors in the brain and therefore be more responsive to LSD (Bonson and Murphy, 1996). Alternatively, the effects can be enhanced through a dopaminergic system (Bonson and Murphy, 1996) as chronic use of desipramine has been demonstrated to result in an elevated behavioural response to amphetamine, whereas chronic use of fluoxetine did not have the same effect (Spyraki and Fibiger, 1981). Therefore, it is possible that the observed effects are due to a modification in the sensitivity of the dopamine receptors. However, it is noteworthy that the results of TCAs originate from a single study with a very small number of participants.
Other drugs (potentiating action)
Additionally, from the same study, it was reported that chronic lithium use potentiated LSD effects and resulted in earlier onset (Bonson and Murphy, 1996). It has been suggested that while acute administration of lithium increases serotonin levels in the brain, chronic administration on the other hand reduces serotonin concentrations (Bonson and Murphy, 1996). This may explain why long-term lithium use enhances LSD effects, as LSD acts as an agonist in the absence of endogenous serotonin resulting in the observed behavioural effects (Bonson and Murphy, 1996). It is important to note that these findings and some anecdotal reports that can be found on internet forums suggest a similar outcome, there are also several reported instances of lithium causing seizures when used concurrently with LSD or psilocybin (Nayak et al., 2021).
Intensified and prolonged (negative) effects of LSD have also been reported in individuals being pretreated with reserpine, an antihypertensive drug, in three separate studies (Freedman, 1963; Isbell and Logan, 1957; Resnick et al., 1965). Both Freedman (1963) and Isbell and Logan (1957) reported tremors that were present after using LSD together with reserpine. Tremors were not observed during LSD use alone (Isbell and Logan, 1957), and when combined with reserpine, the experience was reported to be less pleasant and lasted longer than LSD use alone (Freedman, 1963). Furthermore, specific types of hallucinations were reported with reserpine treatment that were not observed during LSD use alone. Overall, individuals reported unpleasant experiences when using an LSD with reserpine.
Reserpine has the ability to bind to the storage vesicles of certain neurotransmitters, including dopamine and norepinephrine. This binding inhibits the catecholamine pumps and blocks the uptake of serotonin, norepinephrine and dopamine into the presynaptic storage vesicles. Ultimately, this leads to the depletion of these neurotransmitters by cytoplasmic MAO at both central and peripheral synapses (Cheung and Parmar, 2023). Lower serotonin levels can enhance LSD effects, which was also observed for lithium. However, when LSD was used after chronic administration of lithium, it resulted in positive effects and more vivid hallucinations, without any noted increase in negative psychological or physical side effects as for reserpine. Treatment with reserpine has been known to cause several neurological side effects (Cheung and Parmar, 2023), and studies have shown that in patients with anxiety it can exacerbate their symptoms (Peterfy et al., 1976; Sarwer-Foner and Ogle, 1956). Since LSD can also increase anxiety as a side effect (Strassman, 1984), it is possible that reserpine treatment could enhance this effect (or vice versa), while the modulated hallucinogenic effects of LSD are due to depleted serotonin levels.
Moreover, reserpine is also a potent inhibitor of both CYP2C19 and CYP2D6 enzymes (Englund et al., 2014), which are involved in the initial metabolic steps of LSD (Wagmann et al., 2019). This inhibition may account for the reported prolonged effects of LSD when it was used with reserpine (Freedman, 1963) as it could potentially decrease the metabolism of LSD. Finally, reserpine acts as an inhibitor of P-glycoprotein as well (Englund et al., 2014), which can lead to increased concentrations of P-gp substrates. However, no research has been conducted to investigate the impact of P-gp activity on LSD pharmacokinetics and its effects, making it difficult to assess the potential contribution of P-gp inhibition.
Racemic pindolol, which has an affinity for 5-HT1A and β-adrenergic receptors (Artigas et al., 2001), was reported to intensify the effects of DMT (Strassman, 1996). The authors suggested a buffering effect of 5-HT1A that blocked the 5-HT2-mediated psychedelic effects (Strassman, 1996). However, pindolol has been shown to accelerate, and in some cases, enhance the antidepressant effects of SSRIs. This can be mediated by antagonising 5-HT1A autoreceptors in the midbrain raphe and, as a result, preventing the inhibition of serotonin release (Artigas et al., 2001). DMT is a full agonist at 5-HT1A receptors and has a higher affinity for the receptor than it has for 5-HT2A receptors where it is a weak partial agonist (Kozell et al., 2023). Hypothetically, a similar effect as for SSRIs has been described, and can also modulate the effects of DMT, which may have enhanced effects via activating postsynaptic 5-HT1A receptors while 5-HT1A autoreceptors are blocked.
A 5-HT1 receptor agonist and 5-HT2 receptor antagonist methysergide were similarly shown to potentiate DMT’s effects (Sai-Halasz, 1962). This finding adds more weight to the 5-HT1A receptor contribution to the effects of DMT, especially considering that methysergide is also an antagonist at the 5-HT2A receptors.
With regards to mescaline use and potentiated effects, one case report involved the use of 2C-O, a structural isomer of mescaline that did not appear to have psychedelic properties on its own (Dittrich, 1971). However, when used together with mescaline, a synergistic effect may have occurred that potentiated the effects of mescaline described in the case report.
Other drugs (mixed results)
Azacyclonol, once investigated as a potential antidepressant, was experimented on individuals who administered LSD and mescaline. Two studies showed no effect of blocking LSD’s actions (Clark, 1956; Isbell and Logan, 1957) and one case report describes the same for mescaline (Clark, 1956). These results are in contrast to another study that reported the attenuated (pretreatment) or blocked (acute) effects of mescaline after azacyclonol treatment (Fabing, 1955). The mechanism of action of azacyclonol is not well understood, and the mixed results from the described studies make it difficult to draw firm conclusions and hypotheses.
Recreational drugs
Finally, two studies reported the effects of recreational drugs, including MDMA and alcohol consumption on LSD and alcohol on psilocybin. One study, which investigated the effects of alcohol, found that LSD and psilocybin (to a lesser extent) acted as antagonists to the subjective effects of alcohol, while their own psychedelic effects were mainly unaffected (Barrett et al., 2000). It has been suggested that LSD may block the subjective effects of alcohol by interacting with 5-HT1B and 5-HT1C receptors, which are implicated in the formation of the ethanol cue (Barrett et al., 2000). Evidence shows that agonists of these receptors produce responses similar to those of alcohol, whereas blockade of these receptors interferes with the discriminative stimulus properties of alcohol (Barrett et al., 2000). In the second, a placebo-controlled study (Straumann et al., 2023), LSD was co-administered with MDMA to investigate the benefits of a combination also known as ‘candy-flipping’. While the particular combination did not yield significant changes as assessed by various instruments, it did result in longer-lasting drug effects (averaging 1.5 h longer, compared to LSD alone). Also, the combination led to higher plasma concentrations and an extended plasma elimination half-life for LSD, compared to LSD alone. The prolonged drug effects could be attributed to decreased CYP2D6 activity, an enzyme known to be inhibited by MDMA (O’Mathúna et al., 2008) and involved in LSD metabolism (Vizeli et al., 2021).
Limitations
One of the limitations of this study is the inclusion of a number of old research articles, particularly those published between the 1950s and the 1970s, where many of them provided limited information about the outcomes and/or methods used. Additionally, the limited number of total studies included in this review led to the inclusion of case reports, which may be subject to bias and may provide limited generalisability to larger populations. This review may also have also missed some relevant studies that were published only in non-English languages, which were more common in the early days of research. Finally, this review focused on interactions with LSD, psilocybin, mescaline, 5-MeO-DMT, DMT and ayahuasca, while not including other psychedelics.
Conclusions
In this systematic review, we observed DDIs at both pharmacodynamic and (likely) pharmacokinetic levels that may block or decrease the response to psychedelics, or alternatively potentiate and lengthen the duration of psychological and/or physical effects. While there is strong evidence of 5-HT2A receptor involvement in the effects of psychedelics, some research included in this review suggests that other serotonin receptors, such as 5-HT1A/B and dopamine receptors, along with altered serotonin levels, may also modulate psychological and/or physical effects. Additionally, a small number of studies reviewed indicated a potential role of the 5-HT1 receptor subtype in modulating the effects of DMT. It appears that although different psychedelics may yield similar subjective effects, their pharmacological properties differ, resulting in potentially varying interaction effects when combined with other drugs. Overall, given the limited number of papers exploring DDIs associated with psychedelics and the resurgence of scientific and medical interest in these compounds, further research is needed to improve understanding of such interactions, and identify novel drug interactions and potentially serious adverse reactions not currently described in the literature.
Supplemental Material
sj-docx-1-jop-10.1177_02698811231211219 – Supplemental material for Drug–drug interactions involving classic psychedelics: A systematic review
Supplemental material, sj-docx-1-jop-10.1177_02698811231211219 for Drug–drug interactions involving classic psychedelics: A systematic review by Andreas Halman, Geraldine Kong, Jerome Sarris and Daniel Perkins in Journal of Psychopharmacology
Footnotes
Acknowledgements
The authors acknowledge Eleanor White for her contribution to an earlier version of the introduction.
Declaration of conflicting interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: JS and DP are directors of a not-for-profit research institute which has in the past received commercial funding to undertake psychedelic medicines research.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.