
Abstract
Serum supplementation is crucial in in vitro cell culture to provide all the essential nutrients needed for cellular processes. Fetal bovine serum (FBS) is considered the ‘gold standard’, but its production raises serious ethical concerns. Human-derived alternatives to FBS exist in the form of human platelet lysates (hPLs) or human AB serum (ABS). However, these serum products are usually pooled from several donors, in order to have a standardised product without patient-specific deviations. Nevertheless, the use of patient-specific serum in cell culture might be the key to successful transplantation of the cultured cells in medical applications, particularly as it avoids the transmission of infectious components or xenogenic proteins. In addition, the production of non-pooled hPL from single donors is likely to be a cost-effective and time-saving method. The current study used hPL units isolated from single donors and tested their performance as medium supplements for cell culture in comparison with FBS or ABS. This proof-of-concept study aimed to assess the potential of non-pooled hPL for personalised serum supplementation, and thus optimise in vitro models by making them more relevant to human physiology. We showed that A549, HepG2 and Caco-2 human cell lines were generally able to adapt to the new culture conditions and maintain viability, morphology and certain cell-specific characteristics. These results indicate that non-pooled, single patient-derived hPL could be a suitable alternative for in vitro serum supplementation.
Introduction
The practice of adding serum supplements to the culture media has always been part of in vitro cell culture methodology. These supplements crucially provide all of the essential nutrients needed for the attachment, metabolism, growth, proliferation and differentiation of cells cultured in vitro. 1,2 Fetal bovine serum (FBS) is the serum ‘gold standard’, with a worldwide consumption of around 800,000 litres per year. 3,4 For the production of one single litre of FBS, two to three bovine fetuses are needed, thus amounting to around two million fetuses being used each year for this sole purpose. 3,5 Furthermore, the fetus harvesting required for the production process raises serious ethical concerns, because of the potential suffering of the bovine fetuses. Since the use of in vitro methods in research and testing is steadily growing year on year, in order to comply with the Three Rs principles of reduction and replacement, the demand for FBS is likely to increase significantly in the near future. 3,4 At the moment, a total ban on FBS use in cell culture would be nearly impossible, due to the lack of suitable alternatives.
Although FBS is used in almost every cell culture laboratory, it represents the most critical raw material in the cell culture process. It poses various contamination risks, 6 –8 with concerns raised over the biosafety of FBS due to its potential to introduce endotoxins, mycoplasma, viral contaminants, prion proteins and other bovine infectious agents into in vitro cell cultures. 9 –11 In addition, FBS is a highly complex mixture, providing a large number of low and high molecular weight biomolecules, such as growth factors, hormones, serum and transport proteins, vitamins and trace elements. 2,12 Due to the fact that FBS is a xenogenic compound, cytokine-related responses in cell cultures, or even in patients after transplanting cells that were expanded in serum-containing medium, can be triggered. 4,13 Therefore, its use for the development of new approaches in tissue engineering, or for the expansion of stem cells for therapeutic use, is undesirable. For these reasons, limiting or avoiding the use of FBS in human cell cultures would both comply with the Three Rs and improve in vitro models in general, while also contributing to improved patient safety. 2
Over the past few years, many research groups have worked on the development of various serum replacements, which are referred to as ‘serum-free’, ‘xeno-free’ or ‘chemically defined’ supplements. 3,6,14 A number of studies have shown that serum supplements from various human origins have the potential to replace FBS in cell culture medium and are the better choice for in vitro cultures intended for cell-based human therapies. 15 A human-derived alternative to FBS consists of a lysate of thrombocytes (platelets), called human platelet lysate (hPL). Platelets are a rich source of many cytokines, chemokines and growth factors, such as platelet-derived growth factor (PDGF), epidermal growth factor, basic fibroblast growth factor (b-FGF), transforming growth factor-β (TGF-β), insulin-like growth factor (IGF-1), vascular endothelial growth factor (VEGF) and so on. 11,16 Once platelets are activated, they release growth factors, mitogenic factors and hormones at sites of injury through exocytosis, 17 promoting the recruitment, growth and morphogenesis of various cells. 18 For hPL preparation, platelet-rich plasma (PRP) is required, which can be directly obtained from the patient, in the form of thrombocyte concentrates, buffy-coats or whole blood. 19 –21 The activation and lysis of platelets is mostly performed through freeze–thaw cycling, which leads to a high amount of protein in the supernatant. 22 The use of hPL has many advantages in comparison to the use of FBS — for example, its human origin and the absence of xenogenic proteins or viruses prevents the risk of infections and immunogenicity in patients and cell culture models.
The use of hPL as a promising source of growth factors for cell culture has been promoted for nearly 40 years. 23 During the last few years, the suitability of hPL for cell culture has been shown for several cell types, such as mesenchymal stem cells, 10,24 fibroblasts, 20 keratinocytes, 25 head and neck cell lines, 1 endothelial cells 26 and kidney cells. 19 hPL is commercially available, but each preparation usually consists of a pool of at least three different blood donors (phPL), to get a standardised product and prevent undesired donor-dependent characteristics. Because of this donor pooling, these existing alternatives (e.g. phPL) are not suitable for personalised medical approaches.
As the number of potential applications for human-induced pluripotent stem cell (hiPSC) technology increases (e.g. for personalised, cell-based therapies or physiologically relevant drug screening), so does the need for patient-specific serum. hiPSCs are patient-derived somatic cells, which have been reprogrammed to a pluripotent state by, for example, the expression of four transcription factors first defined by Takahashi and Yamanaka. 27 This technology enables the direct differentiation of pluripotent cells to multiple cell lineages such as mature neurons, cardiomyocytes and epithelial intestinal or alveolar cells. 28,29 It represents an unlimited source of specialised cells that can be used for cell therapy, research into several diseases (e.g. Parkinson’s disease or cardiovascular diseases) or to address toxicological questions where more-physiological in vitro models are urgently required to replace animal testing. 30 The main advantage of hiPSC-derived cell types intended for cell therapy is the minimal immune response elicited when they are transplanted into patients 30 and, in the case of in vitro models for research, the human and non-carcinogenic origin of the cells. This argument raises the question of why many of the protocols for hiPSC differentiation have been established featuring the use of xenogenic FBS instead of avoiding it. 31,32
In this study, hPL units were produced from blood bank waste products (so-called buffy-coats), each isolated from a single patient (single patient-derived hPL (shPL)), and the efficacy of each unit was assessed in various in vitro cell culture models and compared to that of FBS and human AB serum (ABS). ABS is another suitable alternative for use as a xeno-free protein supplement in cell culture medium, but its potential for widespread use is reduced by the limited volumes produced. 6 In contrast to a number of existing studies, 1,10,19,20,23 –26 in which hPL was generally used as pooled hPL, the main goal of this study was to investigate the suitability of non-pooled hPL from individual blood samples, for use as a serum alternative with specific potential for use in personalised medicine.
Material and Methods
Preparation of hPLs
The preparation procedure was based on a previously published protocol, 24 with slight modification. Human donor buffy-coats, obtained by density gradient centrifugation of 500 ml anticoagulated blood samples, were provided by the blood donor centre Saar-Pfalz GmbH (Saarbrücken, Germany). Each 100 ml buffy-coat sample was transferred, under sterile conditions, into T75 cell culture flasks, diluted with 25 ml phosphate-buffered saline without Ca2+ or Mg2+ (Fisher Scientific, Schwerte, Germany), transferred to sterile 50 ml tubes and centrifuged at 340 × g, at 22°C for 12 minutes. The supernatant containing the platelets (called PRP) was collected and transferred to a fresh 50 ml tube. A 10 μl aliquot of PRP was diluted in 1 ml of ThromboPlus solution (Sarstedt, Nümbrecht, Germany) and transferred into a C-Chip counting chamber (VWR, Darmstadt, Germany) so that the number of platelets could be counted: 1.0 × 109 platelets/ml were required for further processing of the PRP samples into platelet lysates. The PRP samples were frozen at −20°C for 24 hours and then thawed in a water bath at 37°C. This freeze–thaw step was repeated once more in order to disrupt the platelets by hypo-osmotic shock. 22 The cell debris was pelleted by centrifuging at 4000 × g for 15 minutes (at 4°C), and the supernatant was filtered through a 0.2 µm PES syringe filter (VWR) to remove any remaining fragments. The volume of platelet lysate obtained from each buffy-coat sample was 70 ml. Platelet lysates were stored at −20°C until use.
ABS from a human male (sourced in the United States) and FBS (sourced in Colombia) were purchased from Sigma-Aldrich (Munich, Germany) and stored at −20°C until use. These ABS and FBS supplements were used as controls in this study, in order to compare the suitability of in-house-prepared hPL batches with well-established and commercially available serum products for cell culture.
Protein measurement
The amount of total protein contained in three batches of FBS, five batches of shPL and one batch of ABS was determined by ultraviolet–visible (UV-VIS) microvolume photospectrometry (Nanodrop 2000/2000c; Thermo Fisher Scientific, Waltham, MA, USA). FBS, shPL and ABS protein separation was performed by asymmetric flow field-flow fractionation (AF4) (Postnova Analytics GmbH, Landsberg am Lech, Germany), with a refractive index detector, a multi-angle light scattering (MALS) detector with 21 angles and a UV detector measuring at 254 nm. The samples were prepared by diluting the FBS, shPL and ABS 1:9 in 0.9% w/v NaCl. The thermostat was set to 25°C to stabilise the channel temperature. The autosampler temperature was kept constant at 4°C. The data acquisition and MALS calculations were performed by the AF2000 Control Unit software (Postnova Analytics GmbH). The screening method began with a focus step, with a delay time of 3 minutes, an injection flow of 0.2 ml/min for 4 minutes and a crossflow of 2.0 ml/min. The elution profile for the protein fractions consisted of a 30-minute linear decreasing crossflow at 2.0 ml/min, followed by a 20-minute runtime with no applied crossflow.
Cell culture
The Caco-2 (human colon epithelial), A549 (human lung epithelial) and HepG2 (human liver epithelial) cell lines were obtained from DSMZ (Deutsche Sammlung für Mikroorganismen und Zellkulturen GmbH, Braunschweig, Germany). The Caco-2 cells were cultured in DMEM with high glucose (4.5 g/l), supplemented with 2 mM
Preparation of the cell culture media
DMEM and RPMI cell culture media with FBS, shPL or ABS were prepared in sterile 100 ml flasks. Medium supplemented with either FBS or ABS was prepared by adding 10% v/v FBS or ABS, respectively. shPL-supplemented medium was prepared by adding 10% v/v or 6% v/v shPL and 10 units/ml heparin sodium salt (Serva, Heidelberg, Germany) reconstituted in ultrapure water. All cell culture media were supplemented with 1% v/v penicillin/streptomycin (Invitrogen).
Adaptation of cell lines to new culture conditions
In order to adapt cell lines to new culture conditions, Caco-2, A549 and HepG2 were cultured in cell culture media with increasing concentrations of shPL or ABS, and decreasing concentrations of FBS, over a period of 6 weeks. The cells were seeded at a density of 0.5 × 106 per T25 flask and cultured for 1 week in the respective adaptation medium. The cells were passaged up to 2 times a week, counted and seeded again at 0.5 × 106 per T25 flask. The cell culture media were prepared as described earlier and mixed with different FBS:shPL or FBS:ABS ratios, starting with 80% v/v FBS-containing medium and 20% v/v shPL or ABS-containing medium (80:20). The subsequent media ratios used were: 60:40, 50:50, 25:75 and 0:100. Over the course of the 6-week adaptation process, various individual non-pooled shPL units were used from our collected samples, due to limitations in the volume of blood available for use in experiments from one individual donor at the blood donation centre.
Morphological assessment of the cells
During the 6-week adaptation outlined above, the morphology of the cells was monitored via bright field and phase-contrast imaging with an inverted microscope (Olympus IX70; Olympus, Hamburg, Germany).
Metabolic activity and proliferation assays
Trypsinised cells were seeded at a density of 0.1 × 105 per well in 96-well plates and grown for 24 hours at 37°C and 5% v/v CO2. The test media were added to the cells and the plates were incubated for 24 hours at 37°C and 5% CO2. For the metabolic activity assay, WST-1 cell proliferation reagent (Roche, Mannheim, Germany) was diluted to 10% v/v in culture medium and added to the cells according to the manufacturer’s instructions. The plates were incubated for a maximum of 3 hours, prior to quantification. For the proliferation assay, a BrdU colorimetric cell proliferation ELISA (Roche) was performed according to the manufacturer’s instructions. For both assays, the absorbance was determined with a multi-plate reader (Infinite F200; Tecan, Crailsheim, Germany) at 450 nm (reference wavelength 690 nm) and the values were blank-corrected. The values of the control samples (cells cultured with FBS as the standard condition) were considered to be 100%.
Spheroid formation with A549 and HepG2 cells
Single cell suspensions of 0.25 × 105 cells/ml of A549 or HepG2 cells were prepared in different cell culture media and transferred (100 μl/well) to low attachment spheroid plates (Corning Inc., Corning, NY, USA). The cells were cultured for 7 days at 37°C, 5% v/v CO2. Twice a week, two-thirds of the medium was replaced with fresh medium. Bright field images were taken with an inverted microscope (Olympus IX70) to capture the shape of the spheroids.
TEER measurement and fluorescein sodium permeability assay
Caco-2 cells were seeded at a density of 0.15 × 106 cells, on Transwell® inserts with a pore size of 3.0 µm and a growth area of 1.12 cm2 (Corning Inc.), and cultured for 7 days at 37°C, 5% v/v CO2. The medium was exchanged every second day (apical volume: 0.5 ml, basolateral volume: 1.5 ml). The transepithelial electrical resistance (TEER) was measured with a cellZscope® device (nanoAnalytics, Münster, Germany) as an indicator of the extent of cell polarisation and differentiation. To determine barrier permeability, translocation experiments with fluorescein sodium salt were performed. After 7 days in culture, 5 μM fluorescein was added to the cell culture medium in the apical compartment. After a 3-hour static incubation, samples of medium from the basolateral compartment were taken and the fluorescence intensity was measured at 485 nm/535 nm (ex/em) with an Infinite F200 multi-plate reader (Tecan).
Statistical analysis
The data in this study were presented as the mean ± standard deviation (SD). The data were statistically analysed by one-way analysis of variance (ANOVA). The p values are marked as follows: * if p ≤ 0.05 and ** if p ≤ 0.01 when compared to the FBS control, and # if p ≤ 0.05 and ## if p ≤ 0.01 when compared to the ABS control.
Results
In this study, shPL was prepared and used in the maintenance of various in vitro models, in order to assess whether there were strong donor-dependent effects, as compared to commercial pooled ABS and FBS. Commonly used in vitro models were tested, to assess the potential of shPL for use as a culture medium supplement in these systems, particularly for application in the field of stem cell technology for personalised medicine. The cell lines chosen for this study — A549, Caco-2 and HepG2 — are widely used human cancer cell lines of different origin (alveolar, colon and hepatocellular, respectively). They represent important organs of interest for many research topics and applications and are usually cultured with FBS-supplemented culture medium. To the best of our knowledge, this is the first study to assess the growth of these various cell lines with shPL as the medium supplement and to compare the results obtained with those from cultures supplemented with FBS (representing standard conditions) and ABS (a commercial human serum alternative).
Characterisation of the FBS, shPL and human ABS
To compare the composition of FBS, shPL and ABS in terms of protein content, UV-VIS spectroscopy and asymmetrical AF4 were performed. The results of these spectroscopic measurements (Figure 1(a)) indicated that shPL had the highest average protein content (35.22 ± 3.1 mg/ml), which was significantly higher compared to the protein content of both FBS and ABS (22.85 ± 1.6 mg/ml and 23.53 ± 1.9 mg/ml, respectively). The protein separation pattern of FBS, ABS and shPL, as determined by AF4, is shown in Figure 1(b). A multi-modal size distribution, with three MALS 90° signal peaks for FBS and four signal peaks for ABS and shPL, was identified. The peaks represent different protein fractions of increasing molecular weight and increasing elution time. Peak 1 has an average molecular weight of 55.8 ± 6.6 kDa and is present in all samples, together with peak 3 (mean 433 ± 79.5 kDa) and peak 4 (mean 1800 ± 105 kDa). In the case of ABS and shPL, there is one further peak at a retention time of 10 minutes (peak 2) with an average molecular weight of 141 ± 15.0 kDa, which is not present in the FBS samples.
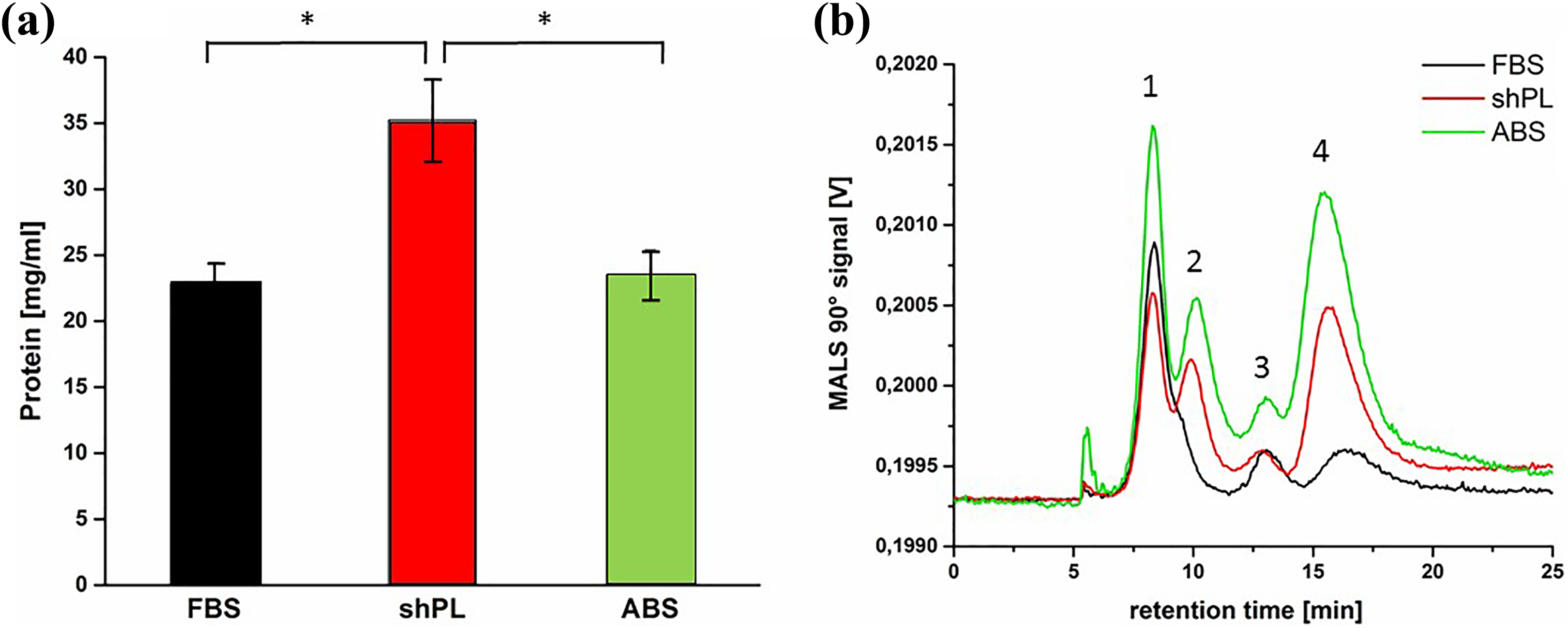
Characterisation of the protein content in samples of FBS, ABS and shPL. (a) The total protein content in FBS, shPL and ABS measured with UV-VIS spectroscopy. The mean values of single measurements for three independent batches of FBS and five independent donors of shPL ± SD are shown; for ABS, the mean values of three measurements from a single batch are shown. One-way ANOVA was performed to compare the protein content (*p < 0.05). (b) MALS 90° signals corresponding to FBS (black), shPL (red) and ABS (green). The graph is representative of three independent measurements. FBS: fetal bovine serum; ABS: human AB serum; shPL: single patient-derived human platelet lysate; ANOVA: analysis of variance; UV-VIS: ultraviolet–visible; MALS: multi-angle light scattering.
Viability and morphology of the cultured cells
Due to the higher protein content of shPL compared to FBS (Figure 1(a)), a lower concentration of shPL (6% v/v) was tested in addition to 10% v/v, to maintain a consistent protein content across all tested supplements (FBS, shPL and ABS). The three epithelial cell lines used in this study were adapted to their new culture media in weekly steps, whereby the ratio of FBS was gradually reduced and the ratio of shPL or ABS was increased. After this process of adaptation, the cells were shown to proliferate well under all culture conditions. Phase-contrast images were used to confirm the typical morphology of each cell line grown in shPL units from three different donors (Figure 2). The images shown are representative of each of the cultures grown in the individual tested shPL units. A549 has an epithelial-like growth pattern, which did not change after the transition from FBS to shPL or ABS supplementation. HepG2 cells, which also have an epithelial-like growth pattern, formed more clusters in shPL and ABS than in FBS. This cell clustering tends to occur when the cells grow on top of each other. However, despite the increase in clustering, the morphology per se did not change. Caco-2 cells kept their epithelial-like growth pattern during and after the transition, with no obvious changes.
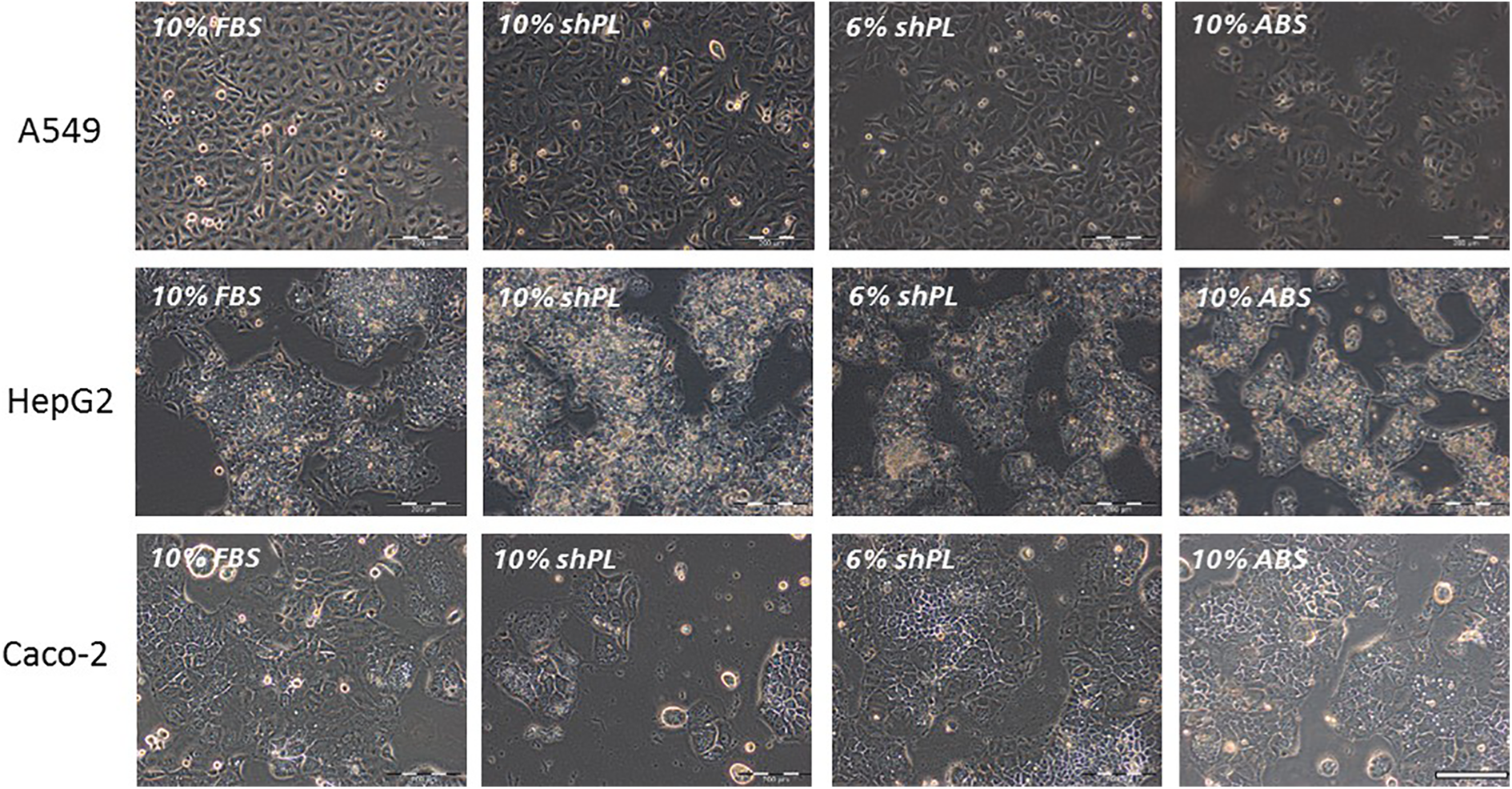
The morphology of A549, HepG2 and Caco-2 cells after a 6-week adaptation to new culture conditions. The cell culture media supplements were: 10% FBS, 10% shPL, 6% shPL and 10% ABS (all v/v). The cell morphology shown in the microscopic phase-contrast images is representative of the morphology of the whole culture and the images shown are representative images, as there was no variation between the cultures supplemented with various individual shPL units. Scale bar = 200 μm. FBS: fetal bovine serum; ABS: human AB serum; shPL: single patient-derived human platelet lysate.
The percentage of viable cells within the cell populations was determined by passaging and counting (Figure 3). Overall, the A549 and HepG2 cell lines maintained high viability (> 90%). It is widely known that Caco-2 cells tend to lose some viability after passaging, even when maintained under standard conditions. Unsurprisingly, Caco-2 cells cultured in 10% FBS and 6% shPL had similar viability (73.3 ± 1.7% and 72.2 ± 0.7%, respectively), whereas their viability in 10% shPL and 10% ABS was significantly lower after the transition from FBS (63.3 ± 0.9% and 61.4 ± 1.6%, respectively).
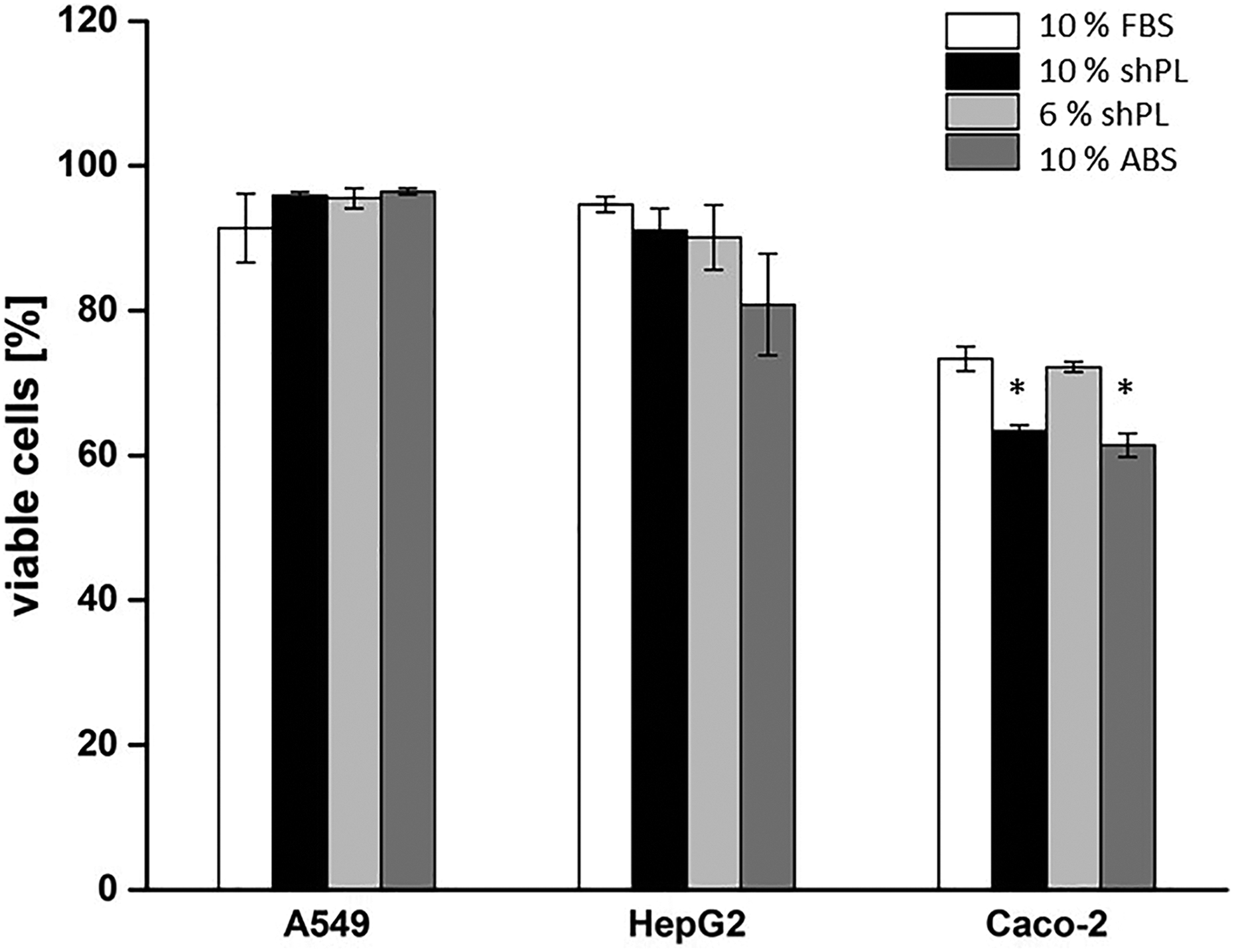
The percentage of viable cells in the different cell populations. The values are expressed as the mean ± SD of three passages after the adaptation of the cell lines to new culture conditions. The data are presented as the mean results of three passages ± SD during culture with the non-pooled shPL units. One-way ANOVA was performed to compare the results of shPL or ABS supplementation with those of FBS (*p < 0.05). FBS: fetal bovine serum; ABS: human AB serum; shPL: single patient-derived human platelet lysate; ANOVA: analysis of variance.
Cell proliferation and metabolism
The cell proliferation rate and metabolic activity of cells adapted to shPL or ABS supplementation were determined after a 24-hour incubation period. The data are presented as the mean of three independent experiments, with one single donor per experiment (in total, three different donors were tested). This was compared to data from cells cultured with FBS (standard conditions, set at 100%) and ABS (Figure 4). As shown in Figure 4(a), the relative proliferation of A549 cells decreased when cultured in the presence of 10% shPL (74.5 ± 10.1%), 6% shPL (74.0 ± 11.0%) and 10% ABS (78.7 ± 13.4%). For HepG2 cells, the highest proliferation was achieved with 6% shPL supplementation (107.8 ± 7.0%). However, a lower proliferation rate was seen with 10% shPL and 10% ABS (83.3 ± 9.3% and 78.4 ± 11.2%, respectively). Caco-2 cells proliferated more rapidly in shPL-supplemented medium (166.8 ± 8.4% and 152.9 ± 8.9% (10% v/v and 6% v/v, respectively)) as compared to FBS (100%) and ABS (99.2 ± 7.3%).
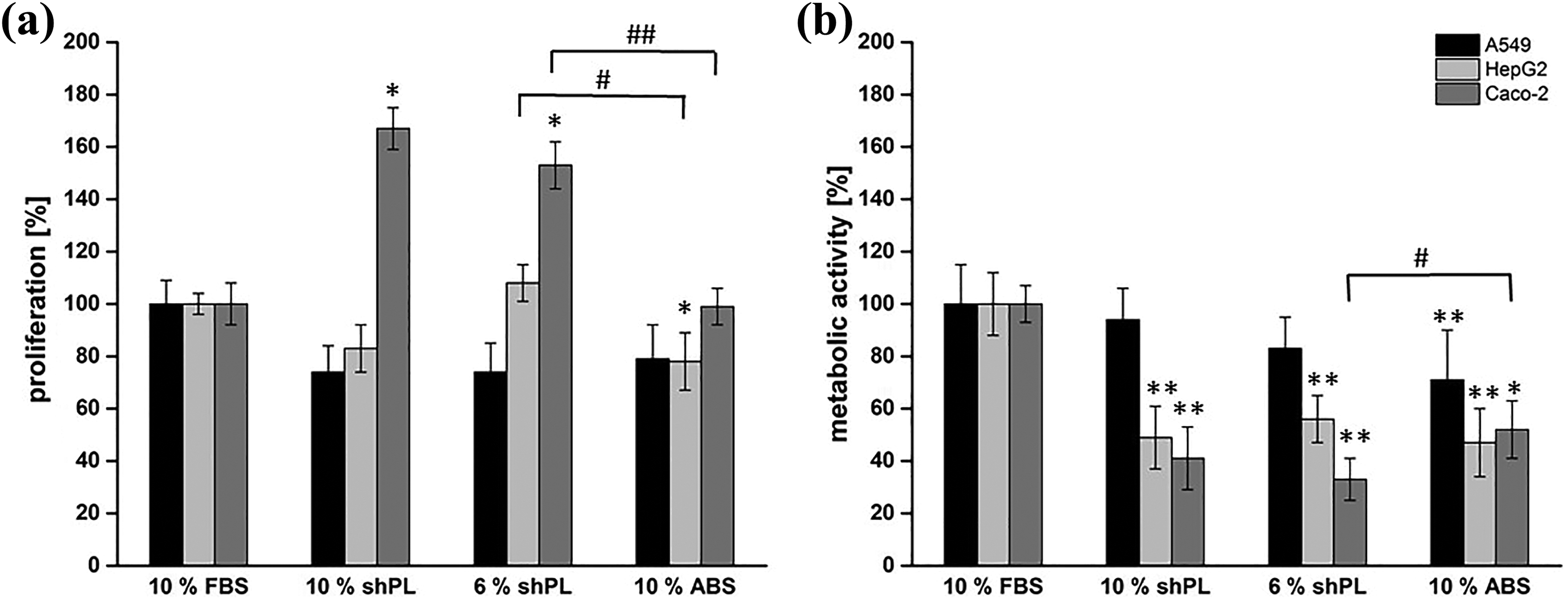
The proliferation rate and metabolic activity of A549, HepG2 and Caco-2 cells after a 6-week adaption to new culture conditions. (a) The proliferation rate was determined after culturing for 24 hours with different serum supplements (10% FBS, 10% shPL, 6% shPL and 10% ABS). (b) The metabolic activity was determined after culturing for 24 hours with different serum supplements (10% FBS, 10% shPL, 6% shPL and 10% ABS). Results obtained under standard conditions (10% FBS supplementation) were considered as 100%. The values are expressed as the mean ± SD of three independent experiments with individual shPL units. A one-way ANOVA was performed to compare the results of shPL and ABS to FBS (*p < 0.05; **p < 0.01) and shPL to ABS results (# p ≤ 0.05; ## p < 0.01). FBS: fetal bovine serum; ABS: human AB serum; shPL: single patient-derived human platelet lysate; ANOVA: analysis of variance.
The metabolic activities of all the cell lines tested were, to some extent, detrimentally affected by the transition from FBS supplementation (Figure 4(b)). For all media supplements, the metabolism of HepG2 cells decreased significantly to around half of that of cells maintained in FBS. Similarly, Caco-2 cells cultured in 10% shPL and 6% shPL showed decreased metabolic activity, down to 41.2 ± 11.8% and 32.9 ± 7.7%, respectively; the corresponding value for Caco-2 cells in 10% ABS was 52.1 ± 11.4%. However, A549 cells maintained a metabolic activity comparable to that of cells grown under standard conditions for all supplements except for 10% ABS.
In addition, in order to confirm that shPL units from different individual donors are suitable for their intended use, we compared the metabolic activities and proliferation rates of cells cultured in shPL and in pooled hPL (phPL). No significant differences between the cells maintained in individual shPL or in phPL were identified (Figure 5).
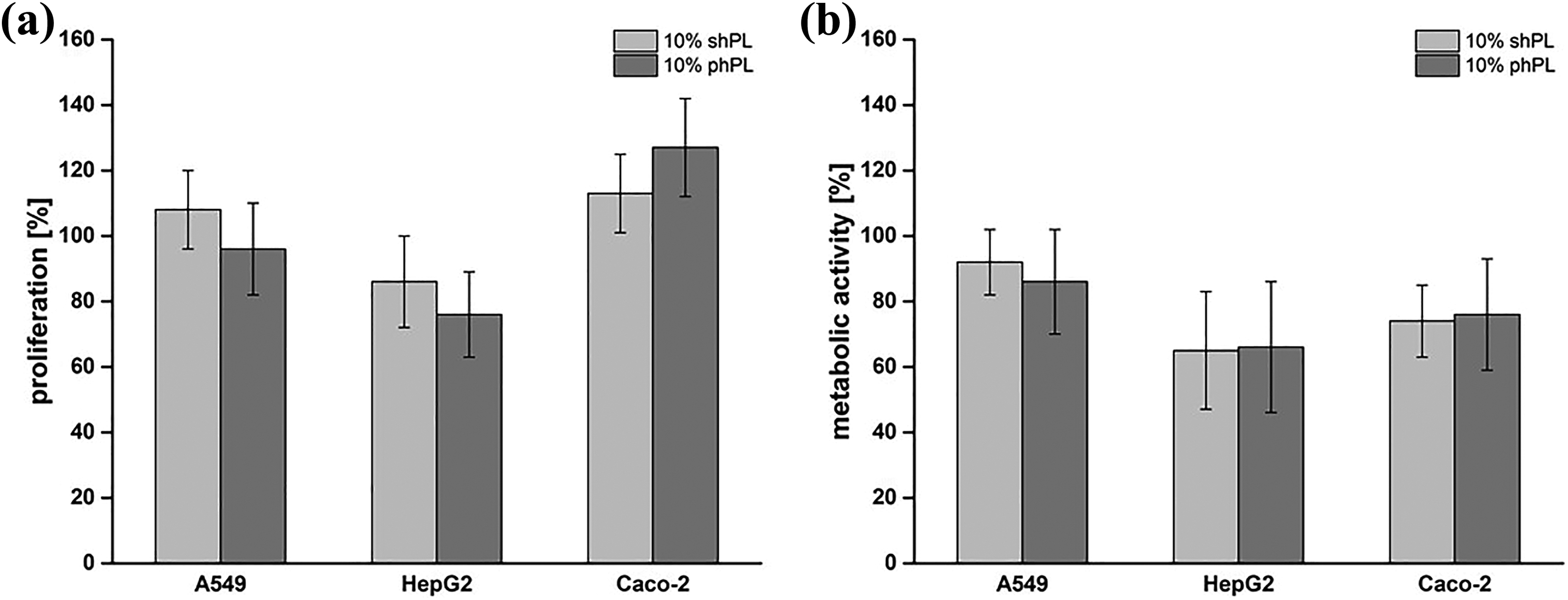
A comparison of the effects of three individual non-pooled shPL and the same three pooled shPL supplementation on the metabolic activity and proliferation rate of A549, HepG2 and Caco-2 cells. The cells were cultured in cell culture medium supplemented with 10% v/v shPL or 10% v/v phPL for 24 hours. The proliferation rate (a) and metabolic activity (b) were determined after a 24-hour culture period under the indicated conditions. Results obtained under standard conditions (10% FBS supplementation) were considered as 100%. The values are expressed as the mean ± SD of three independent experiments. One-way ANOVA was performed to compare the FBS results with the shPL and ABS results. No statistically significant differences were found (p < 0.05). FBS: fetal bovine serum; ABS: human AB serum; shPL: single patient-derived human platelet lysate; ANOVA: analysis of variance; phPL: pooled hPL.
Spheroid formation in A549 and HepG2 cell culture
The popularity of 3-D models, such as multicellular tumour spheroids, has been increasing over the years as they exhibit many characteristics of natural tissue and are suitable for the in vitro screening of drugs. 33,34 The 3-D growth of immortalised cells, such as A549 and HepG2, has been well established and these spheroids can be used for numerous applications. 35,36 Caco-2 cells, on the other hand, are usually used for transport studies in 2-D culture. Thus, we investigated whether A549 and HepG2 cells cultured in shPL or ABS-supplemented medium in low adhesion plates were capable of forming spheroids after 7 days of incubation (Figure 6). The images shown are representative of the cultures grown in the presence of each of the three individual shPL units tested. We found that, in the presence of 10% FBS, both cell lines formed spherical aggregates with diameters of 450–550 μm. When cultured in 10% or 6% shPL, A549 cells assembled into small, loose cell aggregates of no defined form, whereas in 10% ABS, these cells were able to form spheroids with a comparable diameter to those seen in 10% FBS. HepG2 cells formed spheroidal aggregates under all of the tested conditions. However, a looser cell layer surrounded the centre of the spheroids formed in the presence of 10% shPL and 10% ABS, but this was not the case in the presence of 6% shPL. The diameters of the stable centre core were generally comparable to those formed in the presence of 10% FBS, that is, between 450 µm and 500 µm (Figure 6).
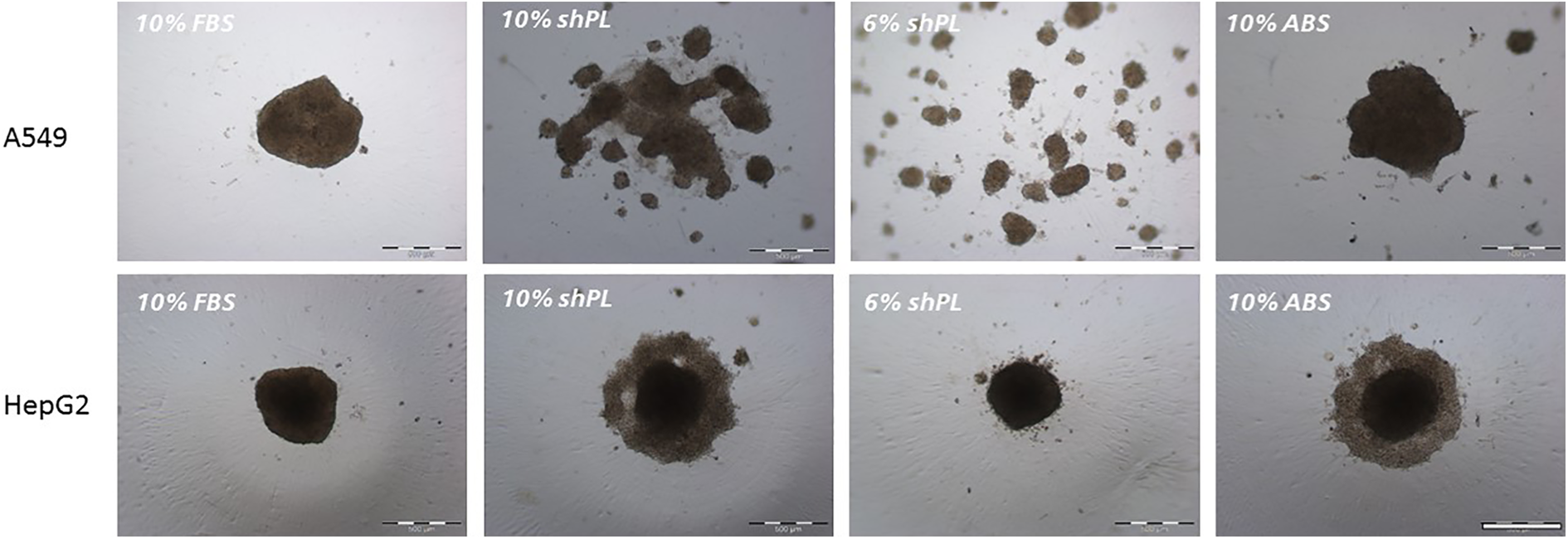
The formation of 3-D spheroids by A549 and HepG2 cells. The bright field microscopy images were taken 7 days after seeding 2500 cells/well. The images are representative of three independent experiments with three different shPL units. Scale bar = 500 μm. FBS: fetal bovine serum; ABS: human AB serum; shPL: single patient-derived human platelet lysate.
Barrier formation by Caco-2 cells
The Caco-2 cell line is widely used as a model for the human intestinal barrier. Cells cultured on porous membranes start to polarise and differentiate into enterocytes with apical brush borders and microvilli. Another characteristic of this cell model is the formation of tight junctions and the accompanying formation of a cell barrier with high electrical resistance.
37,38
In contrast, A549 and HepG2 cells do not form such tight cell layers.
39
In order to investigate these specific characteristics, TEER measurements and a fluorescein permeability assay were performed (Figure 7(a)). Caco-2 cells cultured under standard conditions (with 10% FBS supplementation) had the highest TEER values. The second highest TEER values corresponded to cells in 10% ABS (444.8 ± 109.3 Ω
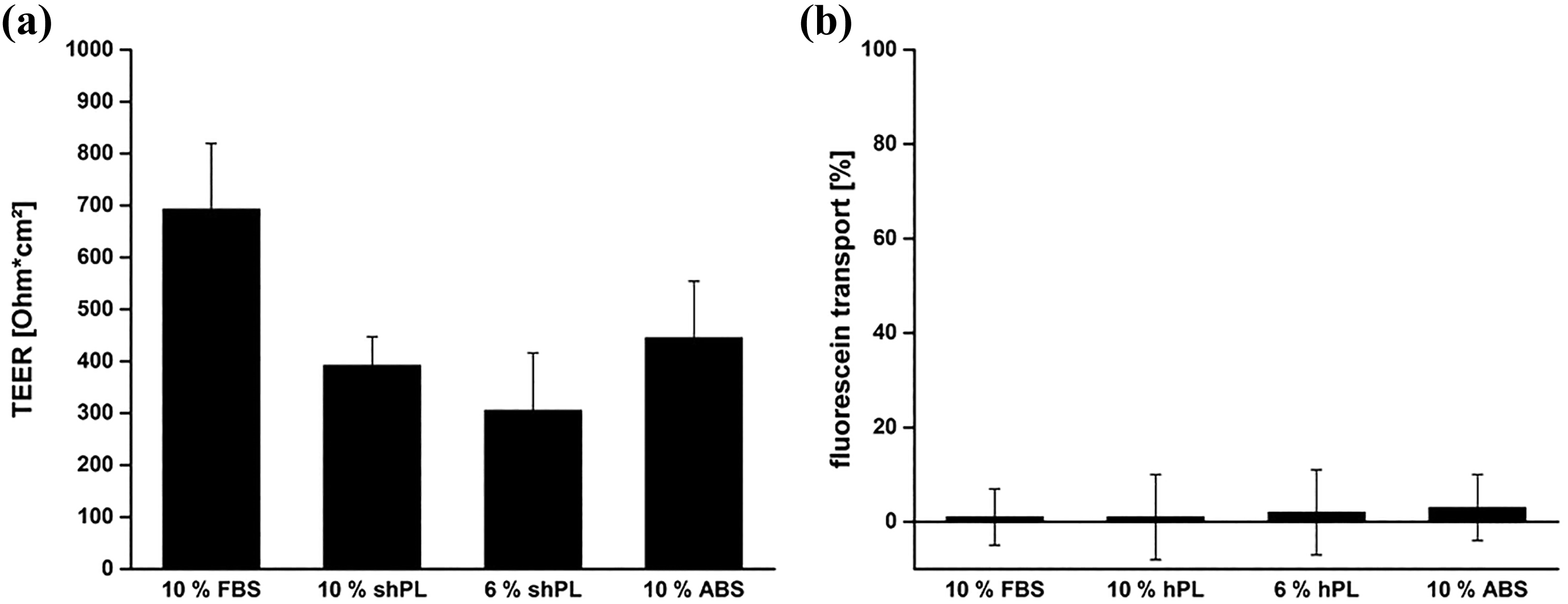
The characteristics of the cell barrier formed by Caco-2 cells after 7 days in culture with different serum supplements. (a) TEER values of the cell layers grown in the different conditions. (b) Fluorescein transport across the cell layers cultured on porous membranes, in comparison to cell-free membranes, which was considered to represent 100%. The values are expressed as the mean ± SD of three independent experiments. The results show the mean results ± SD of three independent experiments with individual shPL units. A one-way ANOVA was performed to compare the FBS results with the shPL and ABS results. No statistically significant differences were identified (p < 0.05). FBS: fetal bovine serum; ABS: human AB serum; shPL: single patient-derived human platelet lysate; ANOVA: analysis of variance; TEER: transepithelial electrical resistance.
Summary of the results
In this proof-of-concept study, the production and use of shPL in cell culture was assessed with regard to its simplicity and cost-effectiveness, and for use in a personalised human serum supplementation approach, for different in vitro applications in the future. The use of shPL as a supplement in cell culture medium was investigated with three human cell lines, of different origins, and by assessing several endpoints. In comparison to ABS, a commercially available human serum, 6% shPL supplementation was shown to be suitable for in vitro culture with no apparent impairment of cell morphology, viability or proliferative capability. In terms of metabolism, HepG2 and Caco-2 cells were less active in shPL-supplemented medium than in ABS-supplemented medium, whereas the metabolic activity of A549 cells remained unaffected. In addition, the data showed that shPL could be used as a supplement when the goal is to form 3-D liver spheroids of HepG2 cells and tight intestinal barriers of Caco-2 cells (Figure 8).
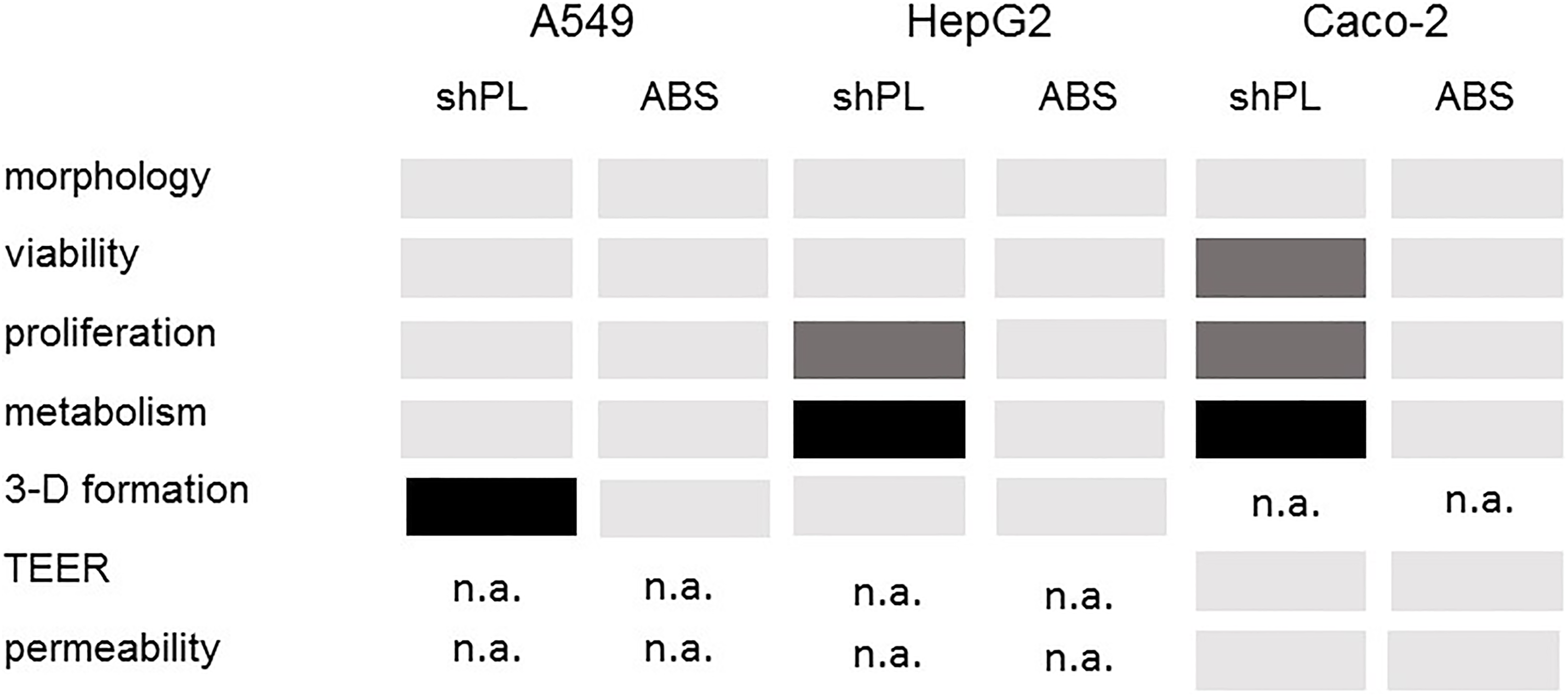
A summary of the results obtained in the current study. The results shown refer to the use of 6% shPL in comparison to 10% ABS as culture media supplements. Light grey: similar results; dark grey: significantly better results; black: significantly worse results; n.a.: not analysed. ABS: human AB serum; shPL: single patient-derived human platelet lysate.
Discussion
There are many scientific and ethical reasons to justify a ban on the use of FBS and other animal sera in in vitro work. This proof-of-concept study, based on the widely used cell lines A549, HepG2 and Caco-2, was performed to assess the potential of non-pooled shPL for use as a serum supplement in cell culture growth media in contrast to commercially-available pooled hPL (phPL).
The three different cell types were cultured in medium supplemented with non-pooled shPL, which was obtained from a blood bank as a waste product. To assess the suitability of shPL for widespread cell culture use, and compare its performance to that of commonly used serum supplements, the cells were also cultured in medium supplemented with FBS and with ABS, which is a commercially available pooled serum product. With a view to using shPL as serum supplement with hiPSC-based cell systems and technologies, we tested several cell characteristics, including morphology, metabolic activity, proliferation, 3-D aggregate formation and barrier formation.
The shPL batches prepared for this study had higher total protein content in comparison to FBS and ABS (Figure 1(a)). Spectrometric analysis revealed an additional protein peak in shPL which was not present in FBS (Figure 1(b)). Judging by the molecular weight of this fraction, it is likely that these proteins are immunoglobulins (IgG), which have a molecular weight of approximately 150 kDa and would originate from the blood used to prepare the shPL. 40 This assumption is supported by the fact that this potential IgG peak was also detected in the ABS sample, and IgG is one of the most abundant proteins in human serum. 41 In contrast, the amount of IgG in FBS is extremely low, 22 probably because FBS is produced by drawing blood from bovine fetuses which have never been in contact with the external environment. This is in line with the results presented by Johannsson et al. 42 It can be assumed that peak 1, which occurred in all samples, is associated with serum albumin (of human or bovine origin), and that peak 3, which was also detected in all samples, could be a mixture of several higher molecular weight proteins present in blood serum (such as bilirubin, thyroxine, cholesterol, progesterone, testosterone or fibrinogen). 43,44 Peak 4, which consisted of very high molecular weight proteins (> 1000 kDa), might include protein agglomerates.
The intensities of the MALS signals were comparable in all of the tested batches of shPL, indicating that the concentration of IgG and other compounds in the bloods of different donors did not vary strongly. Fazzina et al. 11 have shown that different batches of pooled hPL do not differ in their growth factor content (VEGF, PDGF-AB, b-FGF and TGF-β), suggesting that this parameter remains consistent across different donors. 11 In order to confirm that shPL units from different donors are compatible for their intended use, we compared the metabolic activity and proliferation rate of cells cultured in shPL and pooled hPL (phPL). No significant differences between the cells maintained in single donor or pooled hPL units were identified (Figure 5). For this reason, we propose that the culture of cells in shPL-supplemented medium is possible without incurring any result variability from the use of shPL from different donors. These results confirm the potential of shPL for use as a personalised serum supplement.
Due to the higher protein content of shPL, we tested a lower percentage of serum supplementation in the medium (6% as well as 10%). All cell lines cultured in shPL and ABS survived well during the 6-week adaptation process, whereas cells cultured without any serum supplementation died after a short time (data not shown). This demonstrates the capability of shPL and ABS to provide the nutrients and growth factors required by the cells for growth and proliferation, as has been shown with other cell lines in previous studies. 11,21,26 The characteristic morphologies of A549 and Caco-2 cells were fully maintained in shPL, whereas HepG2 cells formed more clusters when cultured in the presence of shPL, compared to FBS and ABS (Figure 2). It has been reported that when cells are cultured under new conditions, certain changes may occur. 45 However, with regard to the percentage of viable cells within the populations (Figure 3), HepG2 and A549 cells cultured in shPL and ABS had comparable viability levels, as compared to cells cultured under standard conditions (10% FBS). In the case of Caco-2 cell viability, 6% shPL was comparable to 10% FBS, whereas 10% shPL and 10% ABS resulted in a 10% decrease in viability. However, it could be shown that Caco-2 cells had a much higher proliferation rate in shPL (both 10% and 6% supplementation) than in FBS or ABS. A higher proliferation rate may be apparent because hPL is known to be a rich source of the most potent mitogenic factors derived from activated thrombocytes. 20,22,25 However, the Caco-2 cells in our study had a much lower metabolic activity in shPL and ABS (Figure 4). In this case, the increased proliferation rate could be interpreted as a defence reaction to new culture conditions or against antibodies present in the medium supplements, instead of being an indicator of viability. The same behaviour was observed with HepG2 cells cultured in the presence of 6% shPL, whereas in 10% shPL only the metabolic activity was reduced. This result is in agreement with the formation of more clusters of HepG2 cells, which occurs when the cells do not spread and instead start growing on top of each other as a response to changes in the cell culture medium. 46
A549 cells proliferated less under the adapted serum conditions, but their metabolic activity was comparable to that in the standard serum. It should be noted that an increase in proliferation rate leads to more cells in the well, which means that a concurrent drop in metabolic activity is in effect more extreme, considering the higher number of potentially metabolically active cells. Conversely, when cells proliferate less rapidly and have a higher metabolic rate in parallel, the effective metabolism per cell is, in reality, much higher. In conclusion, Caco-2 and HepG2 cells proliferated well in shPL but had a very low metabolic activity. A549 cells proliferated less rapidly in shPL, but had a more active metabolism. A possible explanation for the cell-type related differences in these endpoints could be related to the activation of independent mechanisms in the cells, which would depend both on the origin and physiology of the cells and on the individual components of FBS, ABS and shPL. It is possible that a certain component in hPL triggers lung cells to exhibit a higher metabolism, whereas another component (absent from ABS and FBS) triggers hepatic and intestinal cells to exhibit higher proliferation. Therefore, to assess the suitability of different cell culture supplements, it is very important that various endpoints and different cell types are studied.
To confirm the potential of shPL as an alternative serum supplement for cell culture, we performed more-specific analyses. Due to the need for cell models of increased physiological relevance, 3-D microtissues (also called spheroids) are often used in cell biology instead of the conventional 2-D cell layers. 35 In this study, the ability of A549 and HepG2 to form spheroids in shPL-supplemented medium was examined. Both cell lines successfully formed 3-D structures with different appearance. In the case of A549 cells, the serum supplement had a noticeable effect on the structure of the spheroids formed. In the presence of FBS or ABS, the resulting spheroids were round and dense, whereas in the presence of shPL, smaller cell bodies and clumps were visible. HepG2 cells formed round spheroids even in shPL, although 10% shPL and 10% ABS supplementation resulted in spheroids with loose cell layers around a denser centre core. During the formation of 3-D structures, the cells adhere to each other and form tightly packed structures. As expected, extracellular matrix proteins play an important role in this process. 47 A549 cells in the presence of shPL were not able to form spheroids as readily as they did in FBS or ABS, indicating that either crucial factors were absent from, or inhibitory factors were present in, shPL. If the presence of an inhibitor is considered, the most likely candidate protein would be the IgG present in the shPL. However, IgG was also present in ABS (Figure 1(b)), so this is probably not the case at all. Thus it is more likely that factors crucial for the formation of spheroids, at least for A549 cells, are absent from the shPL. The production process of the different sera might offer an explanation for this absence of factors. The shPL is produced from lysed platelets and is rich in PDGFs. ABS, in contrast, is a pooled product from human plasma of the blood group AB and has a different protein composition compared to the shPL and FBS. ABS is also considered to have a broader spectrum of growth factors than the shPL.
The Caco-2 cell line is widely used as an in vitro model for studying the transport of drugs and other substances across the intestinal barrier. Thus, it is crucial that the cell layer develops a tight barrier, as a result of tight junction protein interaction between the cells. 38 The tightness of the cell barrier is usually measured by impedance spectroscopy, which provides information on the TEER of the cell layer. 39 When cultured in shPL or ABS, the TEER values for the Caco-2 cell layers were found to be lower than when the cells were cultured in FBS. To confirm the integrity of the barrier, data on TEER values are often combined with a permeability assay to determine the extent of sodium fluorescein transport through the cell layer. It was concluded that the barrier formed by the Caco-2 cells was tight under all the culture conditions tested, because fluorescein transport was not evident. These results show that even though TEER values were lower, tight junction proteins were expressed during the differentiation process.
This proof-of-concept study suggests that shPL units from three different donors do not differ in their ability to provide the nutrients and growth factors required by the cells to maintain their physiological properties. Thus, non-pooled shPL offers a realistic option for the in vitro culture of hiPSCs for use in the field of personalised medicine.
Conclusion
This study provides useful knowledge regarding the production and use of shPL for cell culture purposes. As a proof-of-concept, it was shown that different human cell lines can adapt to shPL-based culture conditions and maintain their viability, morphology and specific characteristics, with limited exceptions. With regard to translational medicine, there is a huge need to identify animal sera alternatives, because xenogenic compounds, such as FBS, can cause severe rejection in patients. Furthermore, the replacement of FBS in cell culture models, in general, will reduce the number of sacrificed animals for in vitro research and improve the cell culture models used in toxicity and pharmaceutical screening by making them more relevant to human physiology. Our study showed that FBS can be successfully replaced by shPL and that donor-related variability is unlikely to be an issue.
Future work will focus on the use of shPL as a serum supplement for hiPSC-derived cell types. These cells, which are of non-carcinogenic origin, show promise for use in the field of personalised medicine, but they are known to be highly sensitive to the challenges of in vitro cell culture.
Footnotes
Acknowledgements
The authors would like to thank the blood donor centre Saar-Pfalz (Saarbrücken, Germany) for providing the buffy-coat samples and the team of Postnova Analytics GmbH (Landsberg am Lech, Germany) for their helpful discussions and support on AF4 data.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Ethical approval
Ethics approval was not required for this research article.
Informed Consent
Informed consent was not required for this research article.