
Abstract
Models of atherosclerosis are used in preclinical studies but often fail to translate to humans. A model that better reflects human atherosclerosis is necessary. We recently engineered the ExeGen™ low-density lipoprotein receptor (LDLR) miniswine, in which the LDL receptor gene is modified to drive hypercholesterolemia and atherosclerosis, and showed diet-related exacerbation of these phenotypes. Five groups of animals, either wild type (+/+) or heterozygous (+/−), were fed either a normal or high-fat diet for 6 months. One group of heterozygous pigs fed a high-fat diet was also administered atorvastatin at 3 mg/kg/day. Clinical chemistry and anatomic pathology parameters were measured biweekly and at termination. The high-fat diet resulted in increased adiposity and interspersion of adipocytes within the salivary glands. The heterozygous pigs on the high-fat diet gained more weight and had significant increases in total cholesterol, high-density lipoprotein, and LDL compared to wild-type animals or heterozygous animals fed a normal diet. Atorvastatin attenuated these parameters, indicating the statin had a beneficial effect, even in a high-fat diet scenario. Atorvastatin treatment also reduced the intensity of Oil Red O staining in pigs on high-fat diet. Atorvastatin-related amelioration of several indices of cardiovascular pathophysiology in this model underscores its utility for drug discovery.
Introduction
Atherosclerosis is the primary cause of cardiovascular disease, which is the most common cause of death in the United States. Atherosclerosis is associated with the accumulation of proliferative smooth muscle cells, macrophages, lipids, cholesterol, calcium deposits, and cellular debris in vessel walls, eventually leading to plaque formation and arterial obstruction (Lusis, Mar, and Pajukanta 2004; Sanz, Moreno, and Fuster 2013; Libby, Lichtman, and Hansson 2013). These plaques can rupture and lead to occlusive thrombosis, resulting in myocardial infarction or stroke (Little et al. 1988; Stone et al. 2011; Nichols 2013).
Current treatment strategies are directed at changing patient lifestyle and diet and decreasing cholesterol via pharmacological methods. Surgical interventions such as balloon angioplasty and stent placement are used for advanced cases. While these therapeutic approaches have benefited many patients with this disease, they are far from ideal. One of the biggest challenges in drug or medical device development is the insufficiency of the model systems used in these programs (Schwartz et al. 2008; Onuma et al. 2010).
Animal models are the primary means to test the preclinical performance of new therapeutics and medical devices, and the use of inappropriate animal models is inefficient and costly. Several mouse models were generated with mutations in genes important for lipoprotein metabolism, and while these models have been informative, they fail to develop atherosclerotic disease that is typical of the human disease (Bentzon and Falk 2010; Benvenuti et al. 2005). In addition, the small size of the mouse limits the use for the development of human devices and new imaging technologies.
In contrast to mice, the physiology and anatomy of the porcine cardiovascular system closely resembles that of humans. Pigs are frequently used as models of human cardiovascular disease, primarily due to similarities of their cardiovascular systems and their more human-like size (Gerrity et al. 2001) and thus are the preferred species for testing cardiovascular devices (Schwartz et al. 2008; Onuma et al. 2010). Spontaneous atherosclerosis in conventional swine is rare (Skold, Getty, and Ramsey 1966; Bijlenga et al. 1967), but pigs fed a diet high in saturated fat and cholesterol will develop advanced atherosclerotic lesions similar in type and location to those seen in humans (Reitman, Mahley, and Fry 1982; Kobari, Koto, and Tanigawa 1991; Mohiaddin et al. 2004; Dixon et al. 1999). However, the costs and slow disease progression make studies with these diets cost-prohibitive.
The advancement of gene engineering technologies in large animals, particularly miniature swine, has led to the development of new models of human diseases. In cases where the existing models fail to recreate the human disease, genetically engineered miniature swine are an attractive alternative. Several genetically engineered miniature swine have been developed to model diseases such as cancer and cardiac arrhythmia (Al-Mashhadi et al. 2013; Sieren et al. 2014; Park et al. 2015).
In humans, loss-of-function mutations in the low-density lipoprotein receptor (LDLR) gene led to familial hypercholesterolemia, which is associated with elevated plasma concentrations of total and LDL cholesterol (Catapano 2009). Approximately 1 in 500 people carries a mutation in 1 LDLR gene. A recently developed genetically engineered miniature pig with targeted disruptions in one or both LDLR alleles, the ExeGen™ LDLR miniswine, displays human-like atherosclerosis (Davis et al. 2014). We have endeavored here to assess the ability of this pig to serve as a translational model by testing the effect of a statin on the development and progression of atherosclerosis.
Methods
All animal studies were approved by the MPI Research IACUC. Wild-type miniature swine and ExeGen™ LDLR heterozygotes were divided 4 per group into 5 groups as shown in Table 1. All pigs were raised on a standard diet until the age of 5 months, at which time groups 2, 4, and 5 were transitioned to a diet containing ∼40% saturated fat and ∼1% cholesterol from natural sources including eggs and butter (Davis et al. 2014). In addition, group 5 received a daily oral dose of atorvastatin at 3 mg/kg. Individual assigned animal numbers, microchip implant numbers, ear tags, and the study number comprised a unique identification for each animal. Each cage was identified by the animal number, study number, group number, and sex. Animal identification was verified in all procedures during the study.
Design of the Study Groups.
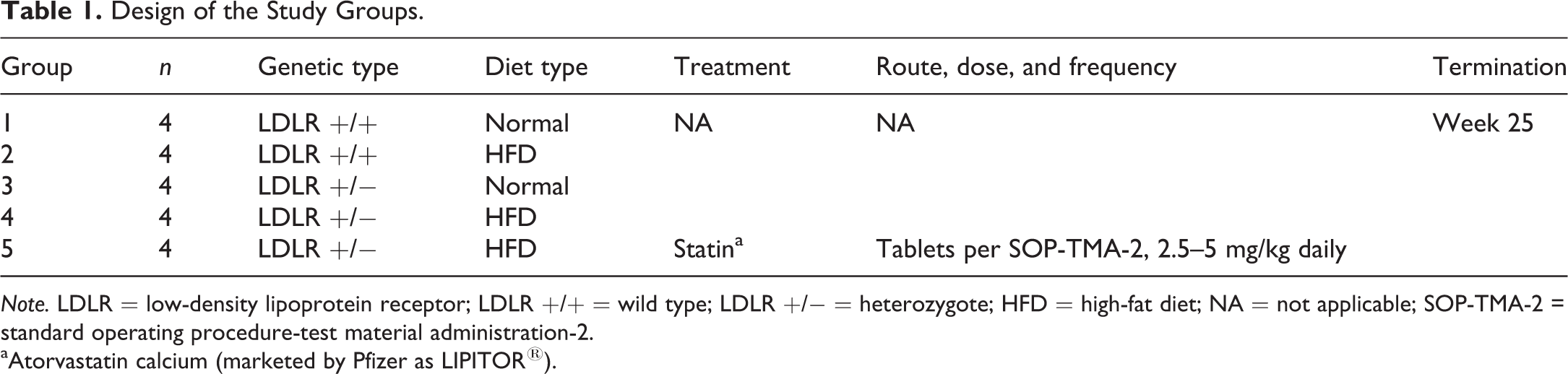
Note. LDLR = low-density lipoprotein receptor; LDLR +/+ = wild type; LDLR +/− = heterozygote; HFD = high-fat diet; NA = not applicable; SOP-TMA-2 = standard operating procedure-test material administration-2.
aAtorvastatin calcium (marketed by Pfizer as LIPITOR®).
Animals were individually housed in pens with raised flooring that provided adequate room for exercise. Animal enrichment was also provided. Fluorescent lighting was on a 12-hr on–off cycle. Temperature and humidity were continuously monitored, recorded, and maintained to the maximum extent possible within the ranges of 61°F–81°F and 30–70%, respectively. All animals were observed for morbidity, mortality, injury, and the availability of food and water, at least twice daily.
The high-fat swine diet was offered via limited feedings, except during designated periods. The lot number from each diet lot used for this study was recorded. Certification analysis of each diet lot was performed by the manufacturer. Tap water was available ad libitum via an automatic watering system.
Lipid profiling was performed every 2 weeks using either a Beckman Coulter Olympus AU2700 or AU640 instrument (The Beckman Coulter Company, Melville, NY). The animals were sacrificed and select arteries collected, stained for lipid deposits, and quantitated after 25 weeks. Data were analyzed as a repeated measures analysis of covariance, using PROC GLIMMIX of SAS/STAT 12.3. Fixed effect terms included sex, treatment group, study week, and all possible interactions. The pretreatment measurement nearest to treatment initiation was included as a covariate. Potential covariance structures for the repeated factor of study week included first-order autoregressive, heterogeneous variance first-order autoregressive, compound symmetry, and heterogeneous compound symmetry. The autoregressive error structures also included a random effect for animal. The covariance structure that yielded the smallest value for Akaike’s information criterion was selected. Marginal means were compared within group, depending on the interactions found significant. All p values for these comparisons were adjusted using the simulation methods given by Edwards and Berry (1987) as implemented in SAS as the SIMULATE= option.
The following artery sections were collected: 2 inches of the aortic arch, the femoral artery include 1 inch above and below the bifurcation to deep femoral artery, and the coronary artery to include 1 inch of mid to distal left anterior descending artery. Aorta, femoral, and coronary arteries were fixed in formalin for 24–48 hr and transferred to phosphate-buffered saline before staining the lumina using Oil Red O and trimmed open for imaging. Images were captured with a digital camera using a National Institute of Standards and Technology–certified ruler in all image fields for calibration. Color measurements were performed using Aperio, a validated image analysis system. Digital images were converted from .jpg format to .svs format and imported for quantification analysis. Image analysis measurements were accomplished using a trained Genie Classifier. Tissues designated for histopathology were fixed in neutral-buffered formalin. Microscopic examination of fixed H&E-stained paraffin sections was performed on aorta, femoral, and coronary arteries from all animals by a board-certified veterinary pathologist.
Results
Body weights were obtained at weekly intervals to compare weight gain in the different groups (Figure 1). Heterozygous animals fed a normal diet (group 1) gained the least weight compared to the high-fat diet groups including both the wild-type and heterozygous animals (groups 2, 4, and 5). At study termination, group 4 animal body weights were 88% greater than the normal diet group, while in groups 2 and 5 the body weights were 72% greater compared to the normal diet-fed animals.
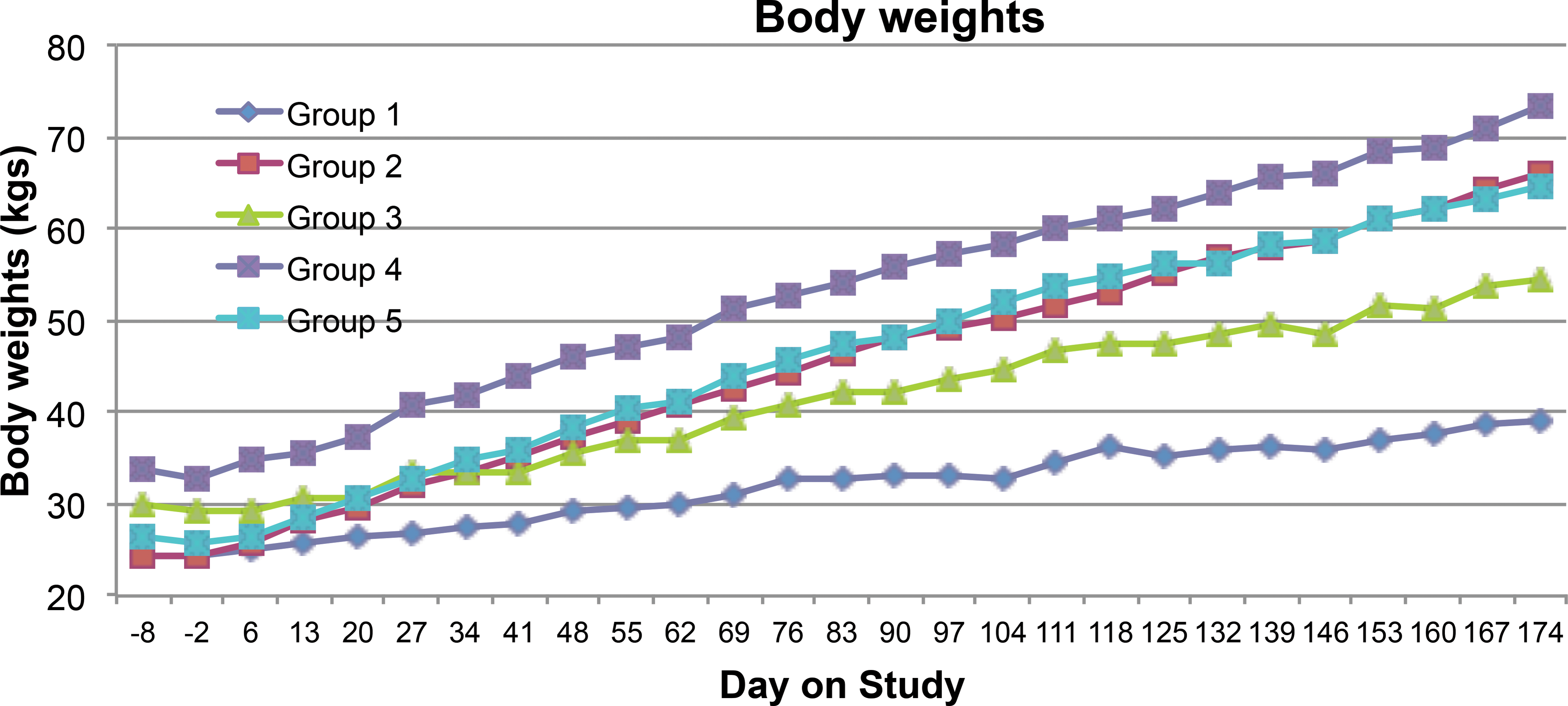
The average body weight of pig measured weekly. Baseline values represented day 6 measurements prior to starting the high-fat diet on day 1 in those groups receiving it. Statin dosing of group 5 was initiated on day 1.
Cholesterol levels were determined every 2 weeks during the study (Figure 2). The rate to steady state circulating levels of total cholesterol varied between the groups. Introduction of the high-fat diet did not result in significant differences in circulating total cholesterol for the wild-type pigs. However, the LDLR mutation resulted in a 2-fold elevation in circulating total cholesterol compared to values in wild-type pigs fed either the normal or high-fat diet. Furthermore, the inclusion of the high-fat diet to heterozygous pigs resulted in a 5-fold increase in circulating total cholesterol compared to wild-type animals fed either diet, and a 2-fold elevation in total circulating cholesterol compared to heterozygous pigs on the normal diet. Statin treatment of the heterozygous animals fed the high-fat diet markedly reduced the elevated total cholesterol to levels that were comparable to the normal diet-fed heterozygotes. A significant decrease in group 5 as compared to group 4 was first detected at day 34 and continued for the remainder of the study.
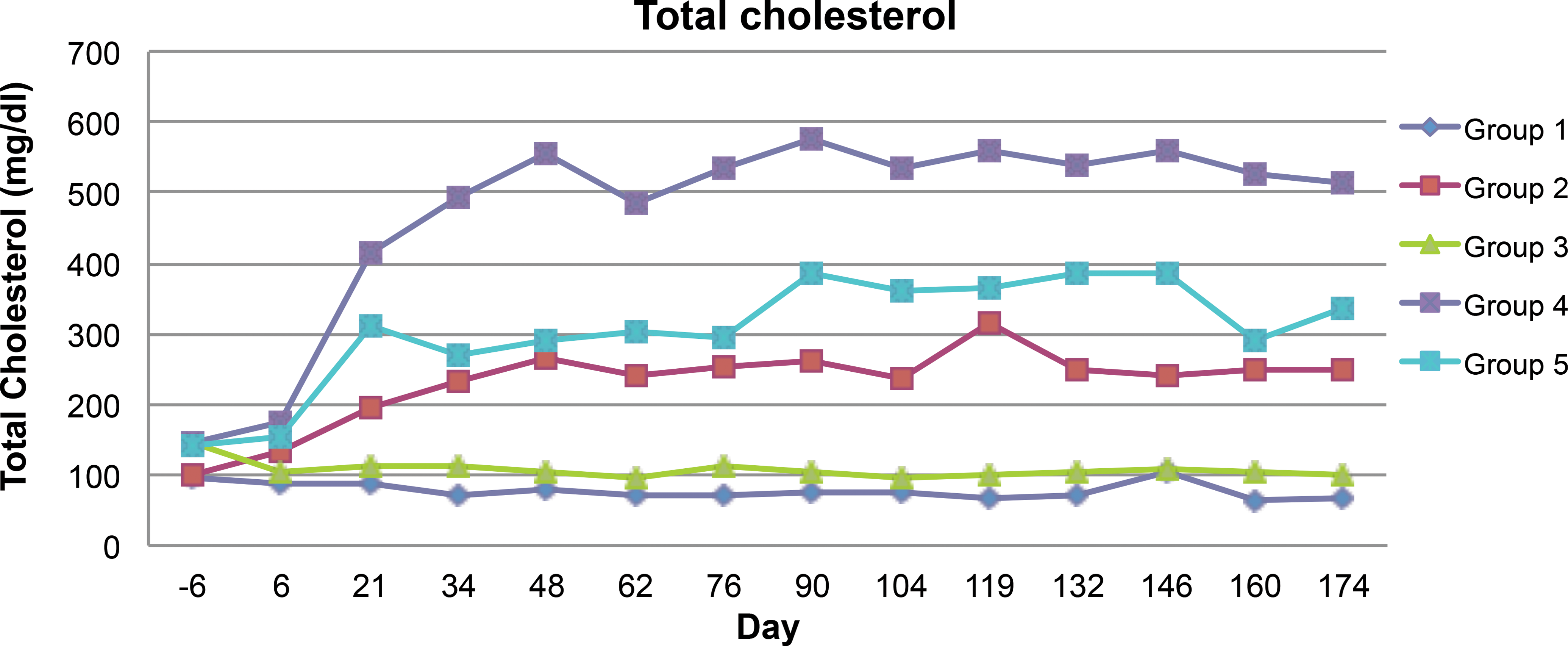
Time course of total cholesterol in the blood during the duration of the study. Samples were obtained every 2 weeks. Baseline values represented day 6 measurements prior to starting the high-fat diet on day 1 in those groups receiving it. Statin dosing of group 5 was initiated on day 1. A significant decrease in group 5 as compared to group 4 was first detected on day 34 and continued throughout the study.
High-density lipoprotein (HDL) cholesterol levels over the course of the study are shown in Figure 3. As was the case with total cholesterol levels, HDL cholesterol approached steady state in all groups between days 6 and 34 and was not altered by the high-fat diet in the wild-type pigs. Similar to total cholesterol, LDLR heterozygotes on normal diets displayed a 2-fold elevation of HDL cholesterol when compared to the wild-type pigs on either diet. The addition of the high-fat diet to LDLR heterozygotes resulted in about 3-fold increase in circulating HDL cholesterol compared to the wild type fed either the normal or high-fat diet, and a 1.5-fold elevation in HDL cholesterol compared to heterozygotes on the normal diet. The statin treatment of heterozygous animals on the high-fat diet markedly reduced the HDL cholesterol to levels comparable with heterozygotes on the normal diet. A significant decrease in group 5 as compared to group 4 was first detected at day 48 and continued for the remainder of the study.
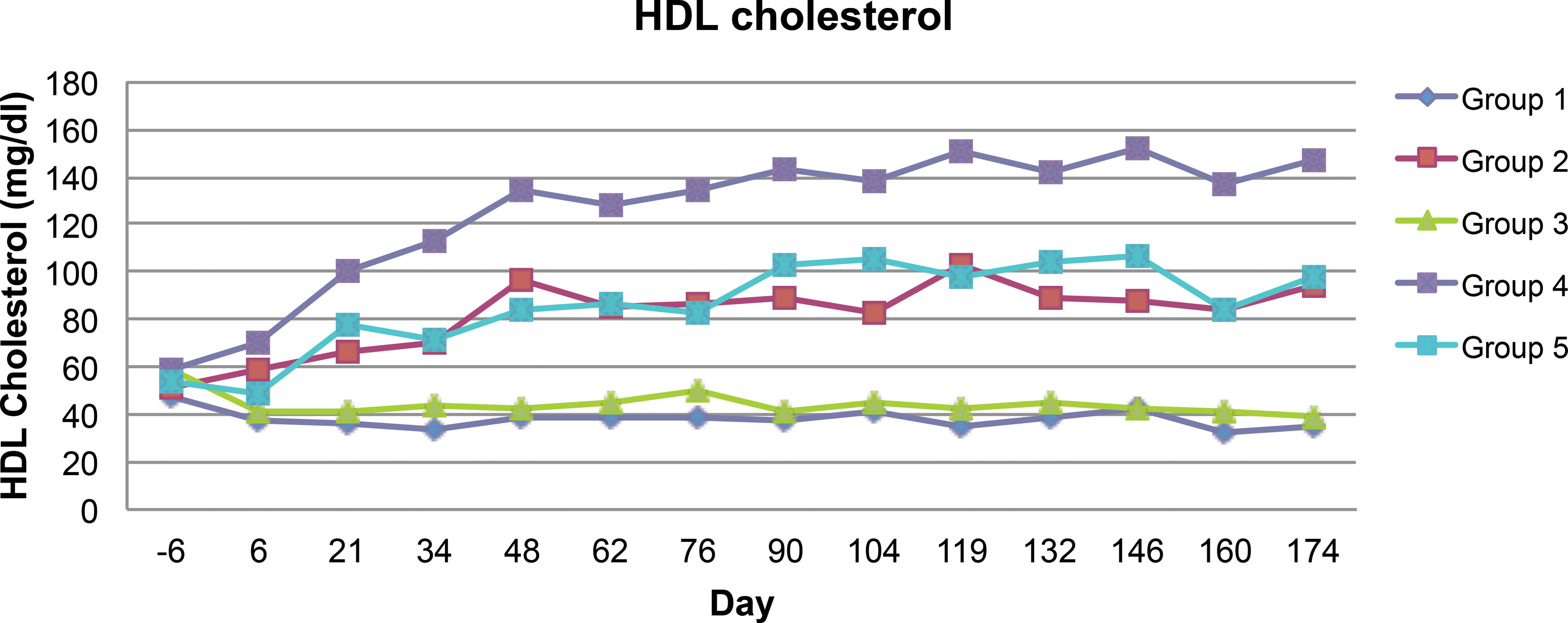
Time course of high-density lipoprotein cholesterol in the blood during the duration of the study. Samples were obtained every 2 weeks. Baseline values represented day 6 measurements prior to starting the high-fat diet on day 1 in those groups receiving it. Statin dosing of group 5 was initiated on day 1. A significant decrease in group 5 as compared to group 4 was first detected on day 48 and continued throughout the study.
LDL cholesterol levels are shown in Figure 4. LDL cholesterol approached steady state in all groups around the same time as HDL and total cholesterol. Following the same pattern as total and HDL cholesterol, no differences in the LDL cholesterol levels were observed between the wild-type animals on either the high-fat or normal diets. LDLR heterozygotes on a normal diet had a 3-fold elevation of LDL cholesterol compared to wild-type pigs on the normal diet. The addition of the high-fat diet to LDLR heterozygous animals resulted in a 6-fold increase in circulating LDL cholesterol compared to wild type on either diet. At study termination, the statin treatment of heterozygous animals reduced the 6-fold elevation of LDL cholesterol to about 4-fold of the levels observed in the wild-type animals. Thus, the statin treatment decreased all the cholesterol parameters. Liver function was unchanged based on total bilirubin and aspartate aminotransferase (AST), while alanine aminotransferase (ALT) measurements were minimally altered in some of the groups (data not shown).
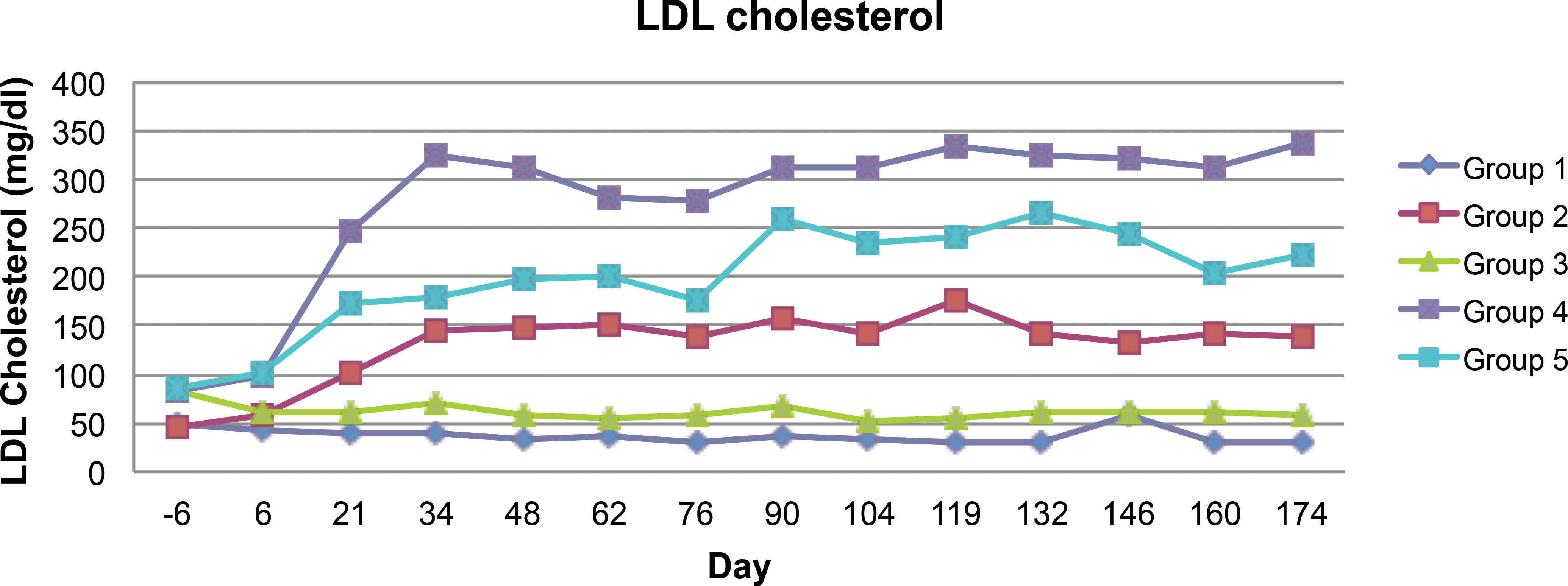
Time course of low-density lipoprotein cholesterol in the blood during the 6-month duration of the study. Samples were obtained every 2 weeks. Baseline values represented day 6 measurements prior to starting the high-fat diet on day 1 in those groups receiving it. Statin dosing of group 5 was initiated on day 1. Significant differences between groups 4 and 5 only occurred at days 36 and 44.
Oil Red O staining was used to assess lipid deposition within arterial lumina (Figures 5 and 6). Both the qualitative (Figure 5) and quantitative (Figure 6) data indicate that LDLR heterozygotes on high-fat diet had intense staining with Oil Red O, consistent with the marked elevations of circulating total cholesterol, HDL, and LDL. Quantitatively, the increase in Oil Red O staining intensity was significantly (p < .05) higher, especially in the aorta and femoral artery, than the same arteries in wild type on any diet and heterozygotes on normal diet (Figure 5). The addition of atorvastatin to heterozygous pigs on high-fat diets reduced Oil Red O staining intensities for the aorta and femoral artery to levels that were statistically comparable to the wild type on high-fat diet and/or normal diets. It is noteworthy that all pigs on high-fat diet generally had a higher Oil red O staining intensity when compared to those on normal diet, regardless of genetic background.
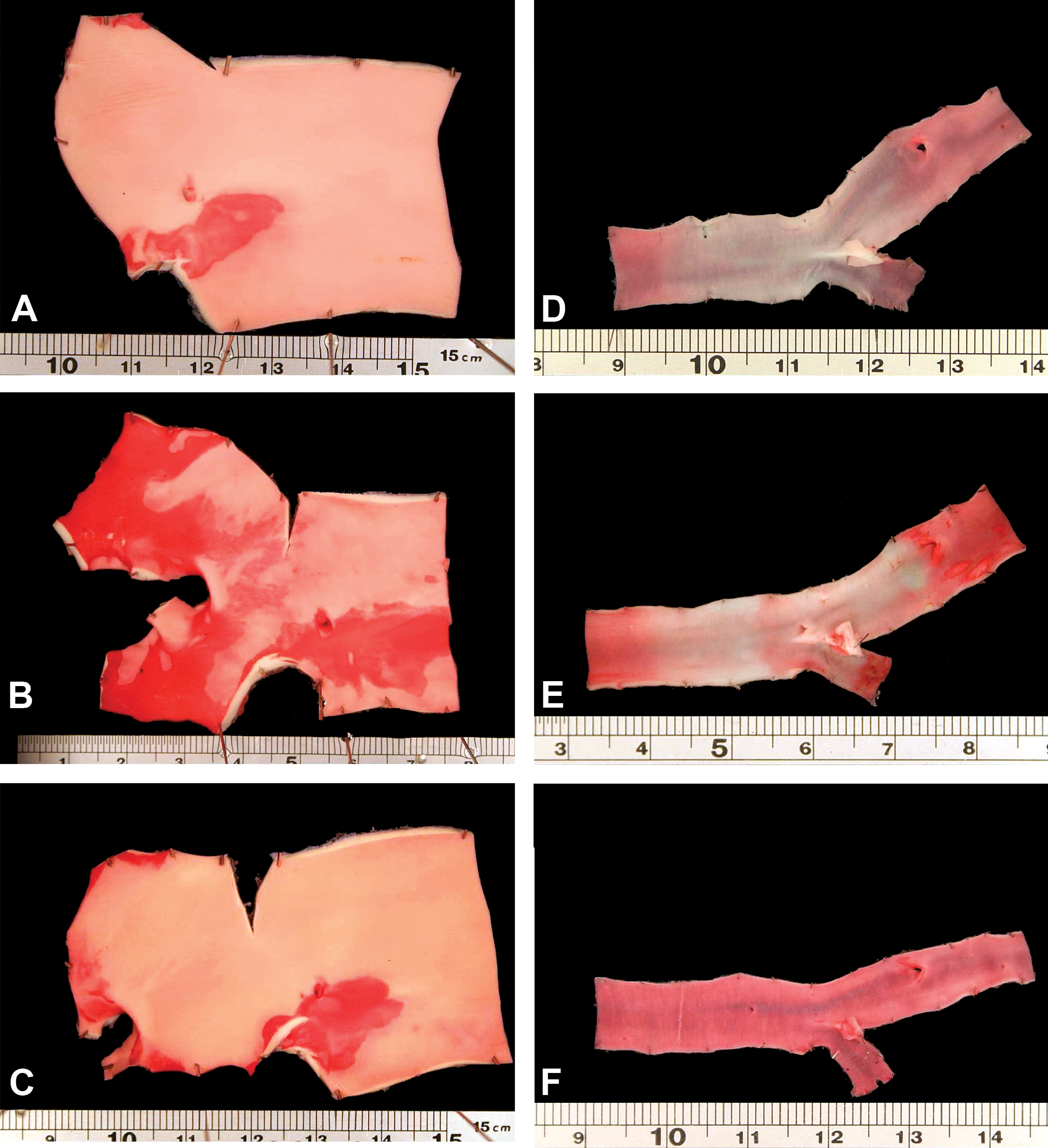
Representative images of Oil Red O staining for aorta (left column) and femoral arteries (right column) of animals on high fat diet at the end of study. Note that among animals on high fat diet, heterozygotes (B and E) had more Oil Red O staining than either the wild type (A and D) or heterozygotes that were treated with statins (C and F).
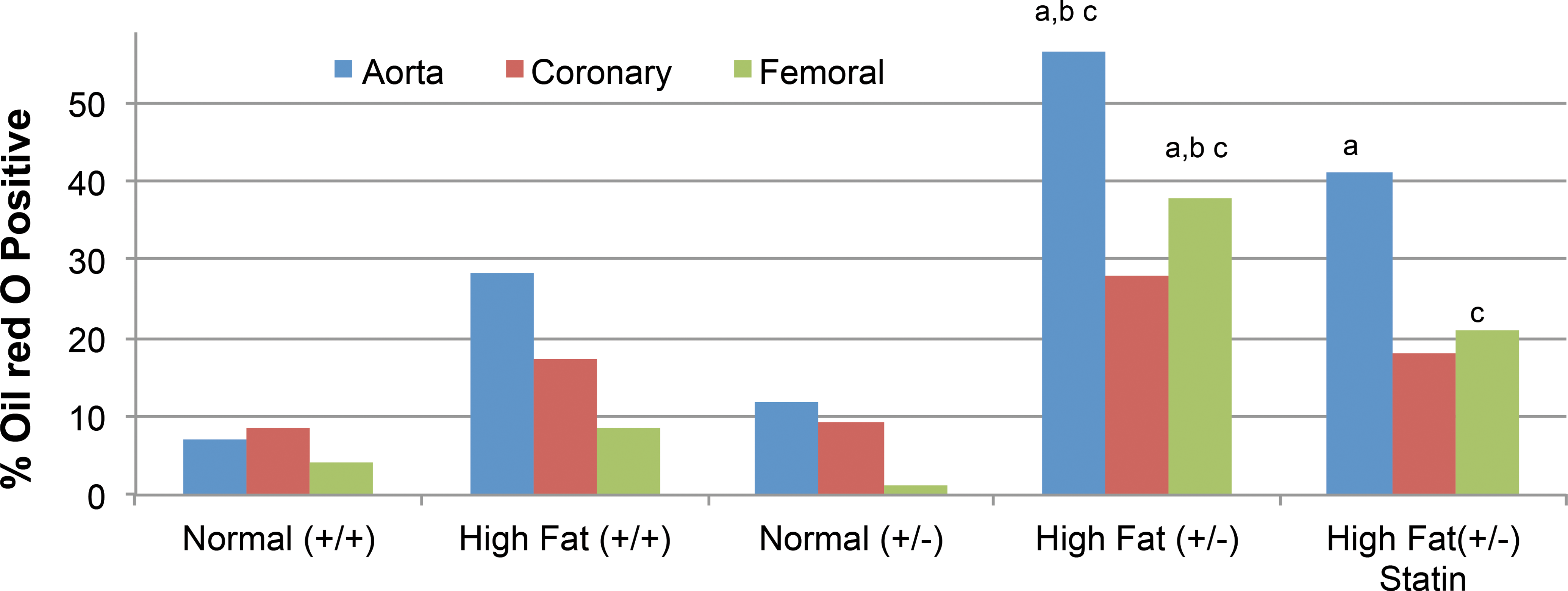
Oil Red O-positive staining on the luminal surface of aorta, femoral, and coronary arteries at the end of study. The text “a” indicates different from wild type on normal diet (p < .05), “b” indicates different from wild type on high-fat diet (p < .05), and “c” indicates different from heterozygotes on normal diet (p < .05). Note that the heterozygotes generally had more Oil Red O staining than the wild type on high-fat diet and that statin cotreatment reduced the magnitude of Oil Red O staining.
Some animals on high-fat diet had minimal to mild proliferative plaques at the intima of aorta, femoral, or coronary arteries (Figure 7). Plaques were often characterized by a discontinuity of internal elastic lamina and infiltration of mononuclear cells interspersed with pale staining extracellular matrix in the subintimal space. No consistent differences in incidence or severity of these morphological arterial changes could be detected due to the small numbers of animals in the experimental groups.
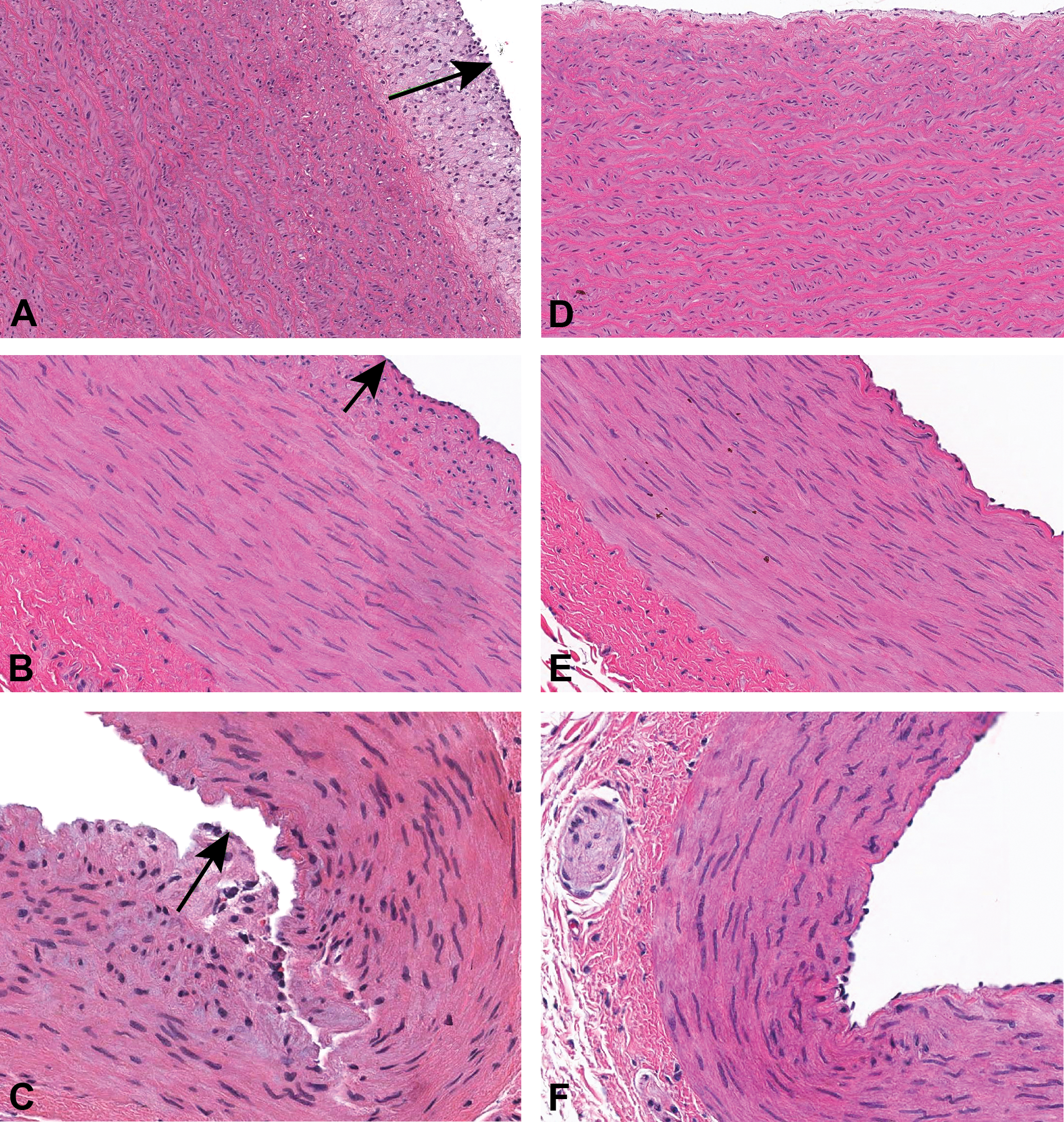
Pigs that were fed high fat diet (left column) had minimal intimal proliferation (beneath arrows) at the aorta (A and D), femoral (B and E) and coronary arteries (C and F). Intimal proliferation was characterized by infiltration of mononuclear cells that are interspersed with pale staining extracellular matrix in the subintimal space with occasional focal distortion of internal elastic lamina.
In pigs on the high-fat diet, regardless of genotype, the mandibular and sublingual salivary glands were morphologically characterized by an increase in the number of adipocytes (Figure 8). Both the mandibular and sublingual salivary glands of pigs on high-fat diet had reduction in acinar surface area by clusters of mature adipocytes that were 70–100 μm in diameter. There were no inflammatory or degenerative changes associated with these mature adipocytes. Occasionally, the increase in mature adipocytes was associated with a reduction in secretory granules of the sublingual salivary gland. The general increase in adiposity in high-fat diet fed animals also reflected as minimal to mid increase in mature adipocytes and/or fat vacuoles in the pancreas, heart, kidney, and adrenal glands of some pigs (data not shown).
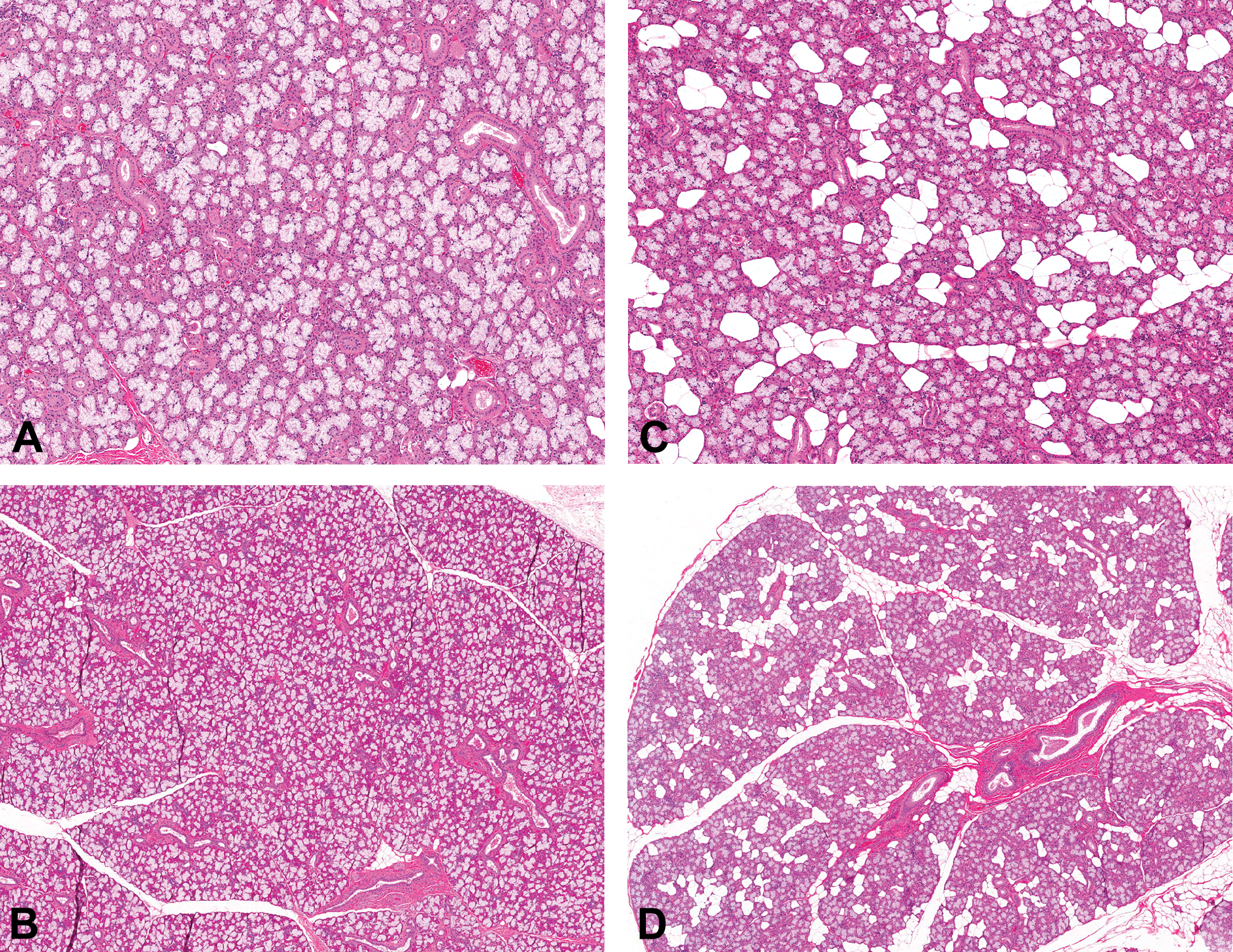
Pigs that were fed high fat diet (right column) had an increase in mature adipocytes interspersed between acini, in both the mandibular (A and C) and sublingual (B and D) salivary glands when compared to those on a normal diet (left column)
Discussion
Miniature swine fed a diet that contains natural sources of cholesterol (1%) and fat (35%) demonstrated significant increases in serum concentrations of total cholesterol, HDL, and LDL compared to a corresponding normal diet group. ExeGen™ LDLR miniature swine demonstrated stable hypercholesterolemia that developed at, or before, 34 days of feeding with high-fat diet as evidenced by total cholesterol levels over 500 mg/dl. The use of diet to elevate total LDL cholesterol on a background of a susceptible population is consistent with the model of cardiovascular disease development in humans where elevation of LDL is associated with an increased risk of coronary heart disease (Kobari, Koto, and Tanigawa 1991). The marked elevation of LDL in the model in this study is one component of its translational utility.
Pigs fed a high-fat diet gained significantly more weight at 6 months whether they were wild type or heterozygous. This weight increase was associated with an overall increase in adiposity across tissues. Minimal to mild increases in mature adipocytes and/or fat vacuoles were seen in the pancreas, heart, kidney, and adrenal glands of these pigs (data not shown). High-fat diet consumption was also associated with morphologic alterations in pigs regardless of their genotype. These included intimal plaques on the walls of different arteries and increased adipocytes within the salivary glands. The increase in adipocytes within salivary glands raises a few questions that were not specifically addressed in this study. These questions relate to the impact of adipocyte increase on the amount of functional acini, volume of salivary secretion, and other digestive functions. Interestingly, hyposalivation has been reported in association with adiposity in obese human patients (Ueda et al. 2013). Therefore, future studies should investigate the functional impact of increased adipocytes within the salivary glands of pigs on high-fat diet.
At the completion of the study, ExeGen™ LDLR heterozygotes also had a higher percentage of Oil Red O-positive staining intensity on the luminal surface of their aorta, femoral, and coronary arteries than wild-type controls on the same high-fat diet. The higher staining intensity is indicative of higher lipid deposition at the arterial luminal wall which is a precursor to other arterial degenerative changes such as atherosclerosis. The general increase in luminal staining likely includes lipids in both intracellular and extracellular locations in/around the arterial lumina, because Oil Red O stains lipids in both extra- and intracellular locations (Kruth 1987). Based on microscopic findings, these areas of increased Oil Red O staining also contained other early alterations in the pathogenesis of atherosclerosis such as endothelial lining alteration and monocyte infiltration, in addition to lipid accumulation.
Atorvastatin administration to ExeGen™ LDLR heterozygotes reduced values of total cholesterol, HDL, and LDL compared to heterozygotes on high-fat diet showing that a statin had a beneficial effect on lipoprotein levels even in a high-fat diet scenario, similar to the effect seen when administered to human patients. ExeGen™ LDLR heterozygotes administered a statin also demonstrated a reduction in the percentage of Oil Red O-positive fatty streaks in comparison to controls. Atorvastatin also attenuated the weight gain that was due to the high-fat diet. These indices of atorvastatin amelioration of disease further highlight the translational utility of this model.
In conclusion, a combination of LDL receptor modification and diet alteration resulted in a stable, consistent miniature pig model of hyperlipidemia with both functional and morphological characteristics that are suggestive of early atherosclerosis. This novel translational model is useful for understanding/altering dyslipidemia in the context of early atherosclerotic lesions, which might be beneficial for developing the next generation of medicine and devices.
Footnotes
Authors’ Note
ExeGen is a trademark of Exemplar Genetics.
Authors’ Contribution
All authors (CA, JS, CR, TV, SD, and DM) contributed to conception or design; data acquisition, analysis, or interpretation; drafting the manuscript; and critically revising the manuscript. All authors gave final approval and agreed to be accountable for all aspects of work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.