
Abstract
Animal models of human diseases are critically necessary for developing an in-depth knowledge of disease development and progression. In addition, animal models are vital to the development of potential treatments or even cures for human diseases. Pigs are exceptional models as their size, physiology, and genetics are closer to that of humans than rodents. In this review, we discuss the use of pigs in human translational research and the evolving technology that has increased the efficiency of genetically engineering pigs. With the emergence of the clustered, regularly interspaced, short palindromic repeat (CRISPR)/CRISPR-associated (Cas) protein 9 system technology, the cost and time it takes to genetically engineer pigs has markedly decreased. We will also discuss the use of another meganuclease, the transcription activator–like effector nucleases , to produce pigs with severe combined immunodeficiency by developing targeted modifications of the recombination activating gene 2 (RAG2). RAG2 mutant pigs may become excellent animals to facilitate the development of xenotransplantation, regenerative medicine, and tumor biology. The use of pig biomedical models is vital for furthering the knowledge of, and for treating human, diseases.
Keywords
Introduction
Medical technology is advancing the knowledge and treatment of human diseases. New medical treatments are developed every day, but before clinical trials can be approved for treatment on humans, preclinical animal experimentation must be completed. Animal models are undeniably essential to understand the mechanisms of disease, develop treatments, identify preventive measures, and even provide cures for human diseases. The Nuremberg Code states that human experimental treatment should only be completed based on the results of preclinical studies using animals as models (Shuster 1997). Therefore, it is crucial that the most accurate physiological animal model of the specific human disease is created. There is an urgency to identify animal models that are best suited to reflect or mimic human disease, although there are limitations to each model.
The most common animal species used for preclinical human trials is the mouse. The mouse is advantageous because it is small, inexpensive, has a short gestation period, and reaches maturity relatively quickly. Researchers are also capable of obtaining embryonic stem cells from mice that can improve the efficiency of genome modification procedures. However, there are a few limitations that make the mouse model not as efficient to replicate and study the human diseases. One of the main limitations is the obvious size difference between mice and humans. The size of the mouse is dramatically smaller, which makes surgical and clinical monitoring almost impractical. Another limitation for the use of mice to model human diseases is the life span of a mouse is significantly shorter than that of a human. This makes mice an inefficient model for age-dependent diseases such as Huntington’s disease (HD). HD is neurodegenerative disease that is caused by an expansion of a cytosine-adenine-guanine repeat greater than 36 repeats in exon 1 of huntingtin (HTT; Pouladi, Morton, and Hayden 2013). This age-dependent neurodegeneration is identified by the formation of aggregates caused by misfolding of proteins due to the mutated HTT gene (Li and Li 2012). There are many rodent models of HD but because of their short life span, they have not become useful models to study the age-dependent neurodegenerative effects seen in humans. Also, their small brain size is an issue for studies involving medical device–delivered drugs and imaging. However, an HD transgenic pig model has been created that exhibits the neurodegeneration similar to what is seen humans (Yang et al. 2010). Although the mouse model is useful in studying the pathogenesis of HD, this pig model is beneficial to study the age-related aspects of HD; imaging of the development of HD; and drug dosage, delivery, and efficacy in a human-sized model.
The ease of creating transgenic mouse models is another advantage to using mice; however, some transgenic mouse models fail to present the same symptoms of human diseases. One example of this is mice with mutations in the chloride ion channel, cystic fibrosis (CF) transmembrane conductance regulator (CFTR) , to model CF. These transgenic mice possess a defective chloride ion channel but do not present the characteristic features of human CF such as abnormalities of the pancreas, lung, intestine, liver, and other organs (Wilke et al. 2011; Rogers et al. 2008). Transgenic pigs with a mutated CFTR gene were created and exhibit the characteristic meconium ileus, exocrine pancreatic destruction, and focal biliary cirrhosis which are also seen in newborn children with CF (Rogers et al. 2008).
Although the species divergence between pig and human occurred around the same time as the species divergence between the mouse and human, the pig sequence is more closely related to human sequence (Wernersson et al. 2005). Pigs have been valued as an excellent animal model for human diseases, since they have similar physiology to humans and have been found useful in surgical, anatomical, nutritional, and toxicological testing (Swindle et al. 2012; Swindle et al. 1994). Minipigs have also shown value and utility as a model for humans and biomedical research (Bode et al. 2010; Vodicka et al. 2005). Pigs possess similar sized organs to humans, have relatively short gestation periods, and give birth to large liters. For each of the aforementioned reasons, pigs may lend themselves to mimic the human diseases more closely than rodent models. The National Swine Resource and Research Center at the University of Missouri (see http://nsrrc.missouri.edu/) has been established to assist with the growing need of pig models.
Methods to Produce Genetically Engineered Pigs
One of the first genetic engineering approaches used in pigs was pronuclear injection of DNA into pig zygotes (Hammer et al. 1985). This technique is very inefficient, with only about 1% of injected zygotes producing transgenic piglets (Prather, Shen, and Dai 2008). Additionally, while large constructs can be injected, there is no control over the site of integration(s). Generally, large numbers of founders are created to determine where the transgene has integrated, tissue specificity and level of expression.
Genetic modification of somatic cells followed by nuclear transfer (SCNT; Whyte and Prather 2011) offers the advantages of knowing the site of integration, the number of copies integrated, and expression (at least in vitro) prior to creating the animal. The SCNT procedure involves removing the metaphase plate and polar body from oocytes arrested at the metaphase II stage of meiosis. Next, a donor cell that has been genetically modified is transferred to the oocyte, and fusion of the donor cell with the oocyte cytoplasm is induced. This transfers the nucleus into the cytoplasm of the oocyte. Finally, the oocyte is activated. Many reconstructed embryos do not progress past the early stages of development; therefore, many embryos are transferred to a recipient at one time. The SCNT process is inefficient and piglets may be born with abnormal phenotypes that may be due to epigenetic reprogramming errors (Prather et al. 2013).
Other methods of genetically engineering pigs include oocyte transduction and sperm-mediated gene transfer. Oocyte transduction uses an inactive virus that does not replicate to deliver the transgene. The inactivated virus is injected under the zona pellucida into the perivitelline space of metaphase II stage oocytes (Cabot et al. 2001). Since these retroviruses integrate into chromosomes that are in metaphase and the oocyte is arrested in metaphase, this makes for a very efficient system. The oocyte can then be fertilized and transferred to a surrogate for gestation. Again, while retroviruses have preferred sites of integration, this method cannot be used to target where the DNA is integrated or control the number of copies that are integrated (Prather et al. 2013). Another approach used to produce transgenic pigs is the sperm-mediated transfection approach (Lavitrano et al. 2003). Here, the transgene is introduced with the sperm at the time of fertilization. The sperm facilitates bringing the transgene into the oocyte where integration can occur. Again, there is no control over the site of integration.
More sophisticated genetic modifications require a system whereby precise genome edits can be made. Such edits may disrupt a gene’s function (knockout), delete a domain (interfere with the intron exon boundary such that an exon is skipped), or cut the DNA for integration of donor DNA by homologous recombination (HR). These editing tools (meganucleases) are proving to be very valuable for the creation of animal models.
Genome Editing with Meganucleases
Recently, meganucleases have advanced gene targeting and transgenic approaches to improve the efficiency of transgenic animal production. This review will briefly describe this technology, but for an exhaustive review of each meganuclease and their application in genetically engineering livestock, see Petersen and Niemann (2015). These targeted meganucleases create site-specific DNA double-strand breaks and cell repair mechanisms can cause random mutations at that location via nonhomologous end joining (NHEJ), or if donor DNA is provided then HR can occur. When a cell repairs DNA by NHEJ, insertion/deletion mutations can be introduced, which can then disrupt the reading frame of a coding sequence. Although there are many meganucleases that have been used for gene editing, including designed endonucleases and engineered meganucleases, here we will discuss only the three most commonly used meganucleases (Menoret et al. 2013; Ashworth et al. 2006). The first, zinc finger nucleases (ZFNs) are composed of a DNA-binding zinc finger protein domain and the nuclease domain derived from the Fok1 restriction enzyme (Kim and Kim 2014). The Cys2-His2 zinc finger domain is one of the most abundant types of DNA-binding motifs found in eukaryotes (Gaj, Gersbach, and Barbas 2013). Each zinc finger is approximately 30 amino acids that generally bind to three DNA base pairs. Typically, there are 3–6 zinc fingers joined together to create a DNA-binding domain with a specificity of 9–18 base pairs per ZFN monomer (Carlson, Fahrenkrug, and Hackett 2012). To create a double-stranded DNA break, dimerization of two FokI nuclease domains is needed. This requires a pair of ZFNs to bind to opposing DNA strands, thus allowing FokI to dimerize and cut the DNA. Since two sets are required, the genome wide DNA specificity increases to 18–36 bp. The cleaved DNA can then be repaired by either HR, if donor DNA is provided, or error-prone NHEJ.
ZFNs have been used successfully to create modification of pig genomes by either electroporation of fetal fibroblast cells followed by SCNT with the edited cells or microinjecting ZFNs directly into zygotes. The first example of ZFN use in pigs was to knock out an enhanced green florescent protein transgene in pigs (Whyte et al. 2011). Later that same year, an α1,3-galactosyltransferase (biallelic knockout model was created by transfecting porcine fibroblasts with ZFNs and SCNT (Hauschild et al. 2011). While another group used ZFNs to disrupt the myostatin gene by both injection into parthenogenetic embryos and modification of fibroblast cells followed by SCNT (X. J. Huang, Zhang, et al. 2014). While efficient at inducing DNA edits, ZFNs are cumbersome and time-consuming to assemble.
The second meganuclease system to be discussed is the transcription activator–like effector nucleases (TALENs). The TALENs have been introduced as an alternative to genome editing with ZFNs. TALENs are similar to ZFNs as they use a FokI restriction enzyme domain that is fused to a tailored DNA-binding domain. However, this DNA-binding domain is comprised of highly conserved repeats from transcription activator–like effectors, which are proteins that are secreted by Xanthomonas spp. bacteria to alter transcription in plant cells (Joung and Sander 2013). TALENs have been directly injected into porcine zygotes targeting the porcine v-rel avian reticuloendotheliosis viral oncogene homolog A gene, for which a tolerance allele for African swine fever has been suggested (Carlson et al. 2012). About 25% of the modified embryos exhibited biallelic modifications (Carlson et al. 2012). Similarly, Lee and coworkers (2014) used TALENs to efficiently create biallelic modifications in the recombination-activating gene 2 (RAG2)-gene producing pigs with severe combined immunodeficiency (SCID; this will be discussed in more detail in this review).
The use of TALENs allows greater flexibility in selecting the target sequences than that of ZFNs (Cermak et al. 2011) and the constructs are easier to build. However, one limitation to ZFNs and TALENs is that they are not efficient at making biallelic modifications, and additional cloning steps or animal breeding is required to produce animals with the wanted biallelic mutations. This process of waiting for the founders to reach puberty in domestic animals is both time-consuming and expensive. In pigs, the generation interval is up to 1 year. Recently, a relatively new endonuclease (our third meganuclease system), the clustered regularly interspaced, short palindromic repeat (CRISPR)/CRISPR-associated (Cas) protein 9 system has revolutionized genome editing (Cong et al. 2013; Hwang et al. 2013; Mali et al. 2013; Cho et al. 2013). This CRISPR technology is based on a bacterial Cas9 nuclease from Streptococcus pyogenes and is part of the bacteria’s adaptive immune response. This immune defense mechanism utilizes short RNA to direct the degradation of foreign DNA. The CRISPR locus consists of four genes, including the Cas9 nuclease as well as two noncoding CRISPR RNAs (crRNAs; Cong et al. 2013). Each crRNA hybridizes with trans-activating RNAs (called guide RNAs) and this pair of RNAs complex with the Cas9 nuclease (Sander and Joung 2014). The protospacer-encoded portion of the crRNA directs Cas9 to cleave complementary target DNA sequences if they lie near a protospacer adjacent motif sequence (Sander and Joung 2014).
The CRISPR/Cas9 system was used successfully to produce CD163 and CD1D knockout pigs both by modifying somatic cells followed by SCNT and by injection of in vitro-derived zygotes (Whitworth et al. 2014). Whitworth and coworkers had 100% efficiency creating genetic modifications in injected zygotes and more importantly, they found embryos with homozygous or bialleleic modifications illustrating the high efficiency of this technology in pigs. We have injected zygotes and produced piglets attempting to edit 4 different genes (CD1D and CD163 are included). Twenty-three piglets have been born and 22 had biallelic mutations (Table 1 and Figure 1). Not only is the technique very efficient at producing gene-edited pigs, but we have seen none of the problems associated with cloning, that is, large tongues, contracted tendons, and so on (Carter et al. 2002).
Generation of Biallelic Edited Pigs by Using the CRISPR/Cas9 System into In Vitro–Produced Zygotes (Unpublished).


CD163 gene edited piglets produced by using the clustered, regularly interspaced short palindromic repeat (CRISPR)/CRISPR-Cas9 system.
Since the CRISPR/Cas9 system was introduced, a number of gene-edited pigs have been produced. Hai and coworkers (2014) found that by injecting CRISPR/Cas9 against the von Willebrand factor (vWF) gene into zygotes, they were able to create piglets with edits in the vWF gene at about 63% efficiency. Similarly, Chinese Bama miniature pig zygotes were injected with NPC1-like single guide RNA/Cas9 and again 100% genetic modification was detected (Wang et al. 2015). Another group used the CRISPR/Cas9 system to modify porcine fetal fibroblasts followed by SCNT for tyrosinase or parkin RBR E3 ubiquitin protein ligase and PTEN-induced putative kinase 1 and the knockout pigs were successfully produced (Zhou et al. 2015). While reports show that CRISPRs have the same specificity as ZFNS and TALENs, it is very easy to quickly assemble them at a minimal cost (Petersen and Niemann 2015). Prior to the introduction of meganucleases, the time line for creating the knockout somatic cells was months to years prior to even performing SCNT to generate the pigs. In contrast, the time line for generation of a pig with a gene knocked out by using the CRISPR/Cas9 technology is now on the order of 5 months.
Useful Models of Human Diseases
SCID
Animals possessing a compromised immune system can be used as a recipient for human-induced pluripotent stem cells (iPSCs) for the potential to regenerate human organs/tissues or for xenotransplantation (Roberts et al. 2015). SCID mice have been developed and are routinely used for tumor growth and treatment (Schmidt-Wolf et al. 1991; Bastide et al. 2002; Lunardi et al. 2014; Clohessy and Pandolfi 2015). Recently, two reports of pigs with SCID have been generated by mutating recombination-activating gene 1 or RAG2 (Lee et al. 2014; J. Huang, Guo, et al. 2014). TALENs were used for transfection of porcine Minnesota minipig fetal fibroblast cells and colonies with modifications were identified. The cells with genetic RAG2 modifications were used for SCNT and edited Minnesota minipigs were generated (Figure 2). The pigs with a mono-allelic modification were predicted to have a nonfunctioning RAG2 gene as a stop codon that was generated in one allele. The RAG2 edited pigs with biallelic modifications exhibited an SCID phenotype and lack T and B cells. Three of the four biallelic modified pigs did not have a thymus and the thymus of the fourth was very small compared to age-matched control pigs. To determine if these pigs are able to accept human grafts, human iPSCs were injected. Not only did the cells survive but by day 12 postinjection tumors were palpable, and by day 28 the tumors were solid. These teratomas contained all three germ cell layers and human mitochondria DNA. This is a great step in identifying a model for testing the safety and efficacy of stem cell therapies. In addition, such pigs would be very useful for whole organ transplants and others studying immunodeficiencies.
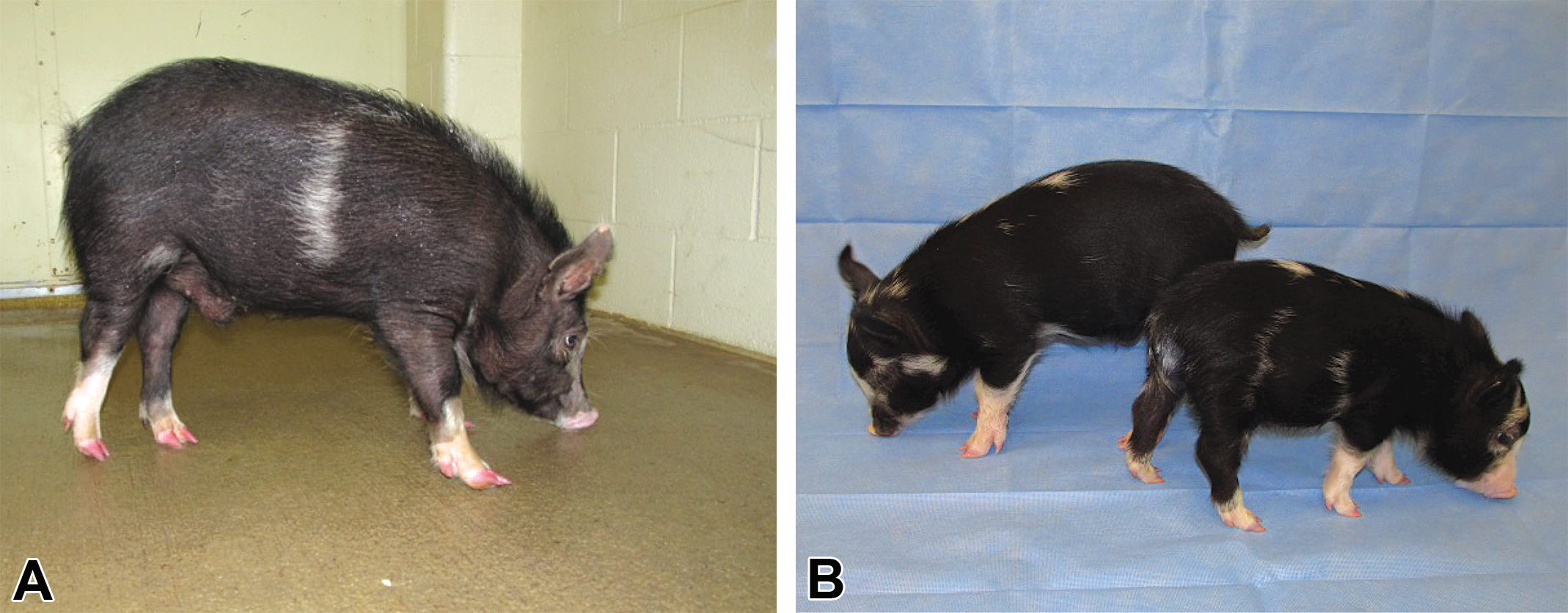
Recombination activating gene 2 (RAG2) monoallelic gene-edited (A) and RAG2 mono- and biallelic gene-edited pigs at 4 weeks of age (B).
Cancer
Cancer is one of the leading causes of death worldwide, which makes the need for cancer detection, diagnosis, and treatment crucial. Having an animal model that recapitulates human tumorigenesis and tumor progression is vital to develop new imaging approaches for noninvasive cancer detection and monitoring, understand drug metabolism studies, implementation of radiation oncology regimes, and for performing surgical training procedures (Sieren et al. 2014; Adam et al. 2007; Schook et al. 2015). Tumor protein 53 (TP53) is a tumor suppressor that is important for regulation of cell cycle, apoptosis, and senescence and is very frequently mutated in human cancers (Levine and Oren 2009). The majority of mutations in TP53 cause impairment in its binding to DNA, including the arginine to histidine mutation at codon 175 (R175H) mutations (Sieren et al. 2014). This same mutation is also found in Li–Fraumeni patients. Recently, a porcine TP53 model was created with the R175H mutation, which corresponds to arginine to histidine mutation at codon 167 (R167H) in pigs (Sieren et al. 2014). The R167 H mutation was introduced by gene targeting in the endogenous TP53 gene and delivered by using recombinant adeno-associated virus to Yucatan minipigs. The R167H cells were used for SCNT and pigs were generated. All TP53 R167H/R167H pigs that reached maturity developed some type of cancer. They primarily developed lymphomas and osteogenic tumors which are also seen in humans with the R175H mutation.
Another “oncopig” model was generated by creating Cre recombinase-inducible porcine transgenes encoding Kirsten rat sarcoma viral oncogene homolog (KRAS)G12D and TP53 R167H. KRAS is a commonly mutated oncogene in human cancers (Jancik et al. 2010). Fibroblasts collected from the transgenic oncopigs treated with adenoCre virus (AdCre) activated the transgene and led to the formation of an in vitro–transformed cell with increased proliferation and migration. These oncopig cells were also highly tumorigenic when explanted into immunocompromised mice. When the oncopigs were injected with AdCre, development of a mesenchymal tumor arose. This was highly reproducible within different injection sites on the same pig and also in different littermate oncopigs (Schook et al. 2015). Together, both of these models provide a platform to better understand and treat cancer in humans.
Conclusions
Pigs have profoundly improved the understanding of human diseases. They make a useful biomedical model to continue to study the pathogenesis, treatment of symptoms, and even identify a cure for the disease. These pigs share many similarities with humans that, along with the advancement of new genetic engineering technology, will result in treatments, therapies, and cures for human diseases.
Footnotes
Acknowledgment
The authors would like to acknowledge funding from the National Institutes of Health (U42OD011140-12).
Author Contribution
Authors contributed to conception or design (BR, RP); drafting the manuscript (BR, RP); and critically revising the manuscript (RP). All authors gave final approval and agreed to be accountable for all aspects of work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.