
Abstract
Incidental findings in the rat eye are not uncommon in acute and long-term toxicological studies. These findings can be associated with a number of causes unrelated to treatment with the test article, including congenital malformation, trauma, infection, metabolic disease, genetic predisposition, and age-related changes. The occurrence of pigment deposition in the retina of Wistar Hannover (Crl:WI (Han)) rats in a 4-week toxicity study is reported in this communication. The microscopic examination of the eyes in the 4-week toxicity study revealed focal yellow–brown pigment deposits in the retina, mainly located in the ganglion cell layer. The retinal pigment deposits were randomly distributed in the control and treated groups and were considered incidental. The deposits were clearly positive for ferric iron in the Perls’ stain but not for lipofuscin by the Schmorl's and Long Ziehl-Neelsen methods. The iron-containing pigment is likely to represent hemosiderin accumulation after retinal micro-hemorrhage or could be indicative of the normal intraretinal iron transport and turnover.
Incidental findings in the rat eye are not uncommon in toxicological studies, irrespective of study duration. Congenital malformations, such as coloboma and retinal dysplasia, are occasionally seen. Age-related changes such as corneal opacity, cataract, and retinal degeneration are common in older rats. Opacity of the lens may result from hereditary, traumatic, metabolic, nutritional, environmental, or age-related causes (Greaves 2007; Gruebbel and Hoenerhoff 2010; McInnes 2012). Accumulation of lipofuscin in the retinal pigment epithelium is a feature of aging found in both rats and humans (Kennedy, Rakoczy, and Constable 1995; Weisse 1995).
This report describes the incidental occurrence of pigment deposition in the ganglion cell layer of the retina of young male and female Wistar Hannover (Crl:WI (Han)) rats in a 4-week toxicity study. Retinal pigmentation or pigment deposits have not been recorded in control rats (421 animals) at Charles River Laboratories (A. Bradley, Charles River Laboratories, personal communication, May 13, 2013), and no other descriptions of similar incidental findings have previously been published in the literature.
Eighty Wistar Hannover (Crl:WI (Han)) rats were necropsied according to protocol after 4 weeks of oral gavage treatment at approximately 10 weeks of age. During the treatment period, a number of procedures were conducted, including ophthalmoscopy. All animal procedures were conducted according to Danish animal welfare legislation as well as to the animal welfare policy of H. Lundbeck A/S. At the end of the treatment period, the animals were subjected to necropsy during which selected organs were weighed and collected. Organs were trimmed and representative samples were taken for histological processing. The samples were embedded in paraffin, cut at a nominal thickness of approximately 5 µm, stained with hematoxylin and eosin (H&E), and examined under a light microscope.
Eyes from all animals showing retinal pigment deposits in the H&E-stained sections were stained for ferric iron/hemosiderin (Perls’ stain) and lipofuscin (Schmorl's, Long Ziehl-Neelsen, and Sudan Black B stain) (Churukian 2002; Luna 1973; Lyon 1985) for identification purposes.
There were no hemorrhages or other ocular findings in any of the animals at the ophthalmoscopic examination or any gross ocular findings at necropsy. At the microscopic examination of the H&E-stained eyes, subtle focal yellow–brown pigment deposits were observed in the retina (Figure 1) of 5 male and 6 female rats. The pigment deposits were mainly located in the ganglion cell layer, but also in the inner and outer segment of rods and cones and in 1 case also in the choroid. In the majority of the rats, the deposits were found in the peripheral retina, but in 1 rat, they were more centrally located.
Retina of Wistar Hannover (Crl:WI (Han)) rat. (A) Pigment deposits in ganglion cell layer and small pigment granule in inner segment of rods and cones. H&E ×40. (B) Yellow–brown pigment deposits in ganglion cell layer. H&E ×63. (C) Pigment deposits in ganglion cell layer staining positive for ferric iron. Perls’ stain ×63. (D) Pigment deposits in ganglion cell layer staining negative with Schmorl’s stain ×63.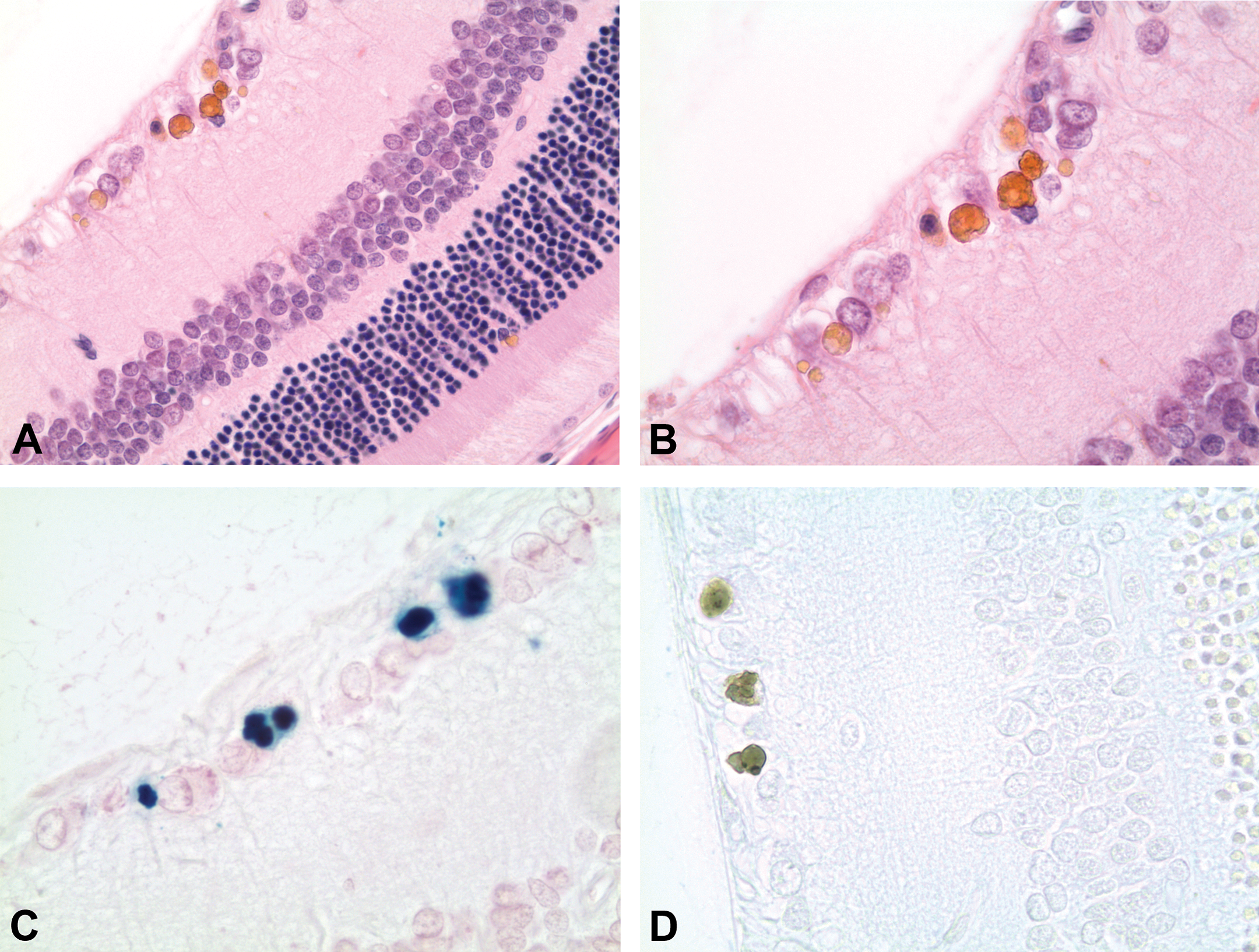
The findings were randomly distributed in the control and treated groups (2/20 control, 3/20 low-dose, 2/20 mid-dose, and 4/20 high-dose animals; Table 1) and therefore considered an incidental finding with no relation to treatment.
Incidence of retinal pigment deposits, H&E stain.
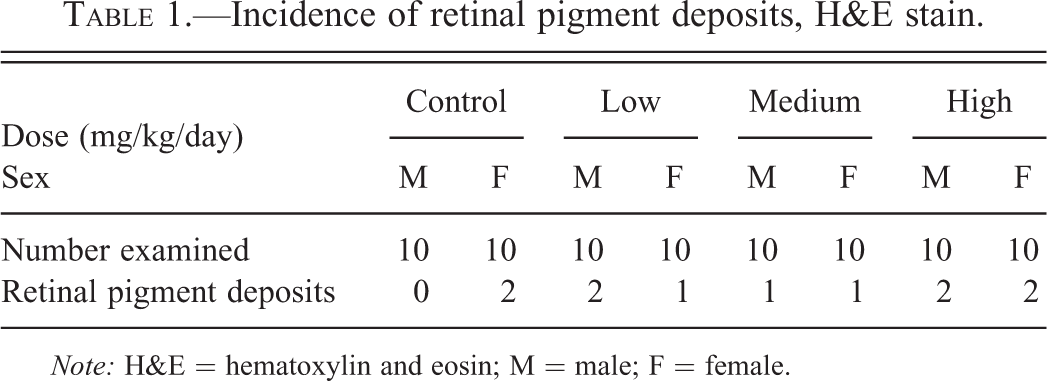
Note: H&E = hematoxylin and eosin; M = male; F = female.
Staining properties of retinal pigment deposits, special stains.

The most likely source of endogenous ferric iron deposits would be hemosiderin, and as capillaries are present throughout the retina, degraded erythrocytes and thereby hemosiderin may be present here as well. In support of this, the deposits stained weakly positive with Sudan Black B, and according to Lyon (1985) hemosiderin may, in some instances, exhibit weak sudanophilia.
The pigment stained negative in the Schmorl's and Long Ziehl-Neelsen stains, indicating that the pigment did not contain lipofuscin (Figure 1, Table 2).
Spontaneous hemorrhage in the retina has been reported in adult rats (Heywood 1973), and hemosiderin has been detected in the rat retina following induction of retinal hemorrhage (Gilliland, Folberg, and Hayreh 2005). In this study, leakage from retinal capillaries leading to retinal micro-hemorrhage may have been aggravated by the handling of animals during dosing, ophthalmoscopy, or blood sampling procedures. During these procedures, the rats were scruffed by holding a flap of skin tensely at the back of the neck thereby tightly stretching the skin of the head and the eye area. Retro-orbital blood sampling was not used in this study.
In growing fetuses, the temporary hyaloid vessels and tunica vasculosa lentis are the route of nutrient supply to the developing lens. Branches of the hyaloid vessels cover the surface of the retina (Cairns 1959) and hemosiderin deposits could originate from leakage of erythrocytes from the hyaloid vessels, as they involute after birth (A. Bradley, Charles River Laboratories, personal communication, May 13, 2013). Indeed, spontaneous hemorrhage was noted microscopically in several ocular tissues, including the retina and choroid, in a recent study of postnatal ocular development in Wistar Hannover, Sprague-Dawley, and Long–Evans rat pups (Inagaki et al. 2014). The retinal hemorrhage had the highest incidence in the Wistar Hannover strain and was located in the inner layer of the ganglionic optic nerve fibers, the nuclear layers, and the choroid (Inagaki et al. 2014). These locations are similar to those of the pigment deposits in the present study. Although the retinal hemorrhages were not observed in pups older than 16 days of age (Inagaki et al. 2014), hemosiderin may persist in the retina for several months (Gilliland, Folberg, and Hayreh 2005). It is therefore likely that the clearance of hemosiderin after postnatal retinal hemorrhage would not be complete in the 10-week-old rats of the present study resulting in the presence of retinal pigment deposits.
It has been shown that iron and the iron storage protein ferritin is widely distributed throughout the retina in rats and that iron is continuously taken up by the rat retina (Moos et al. 2011; Yefimova et al. 2000). The ganglion cell layer, the inner nuclear layer, and the inner segments of the photoreceptor cells have been described to contain the largest amount of iron (Goralska et al. 2009; Yefimova et al. 2000).
The presence of iron is required by a number of physiological processes in the retina, such as the renewal of photoreceptor discs and the phototransduction cascade, and retinal capillaries express transferrin receptors facilitating the uptake of iron-containing transferrin (Moos et al. 2011). The removal of excess iron from the eye is of the utmost importance due to the involvement of iron in reactive oxygen species formation. Data suggest that retinal iron is recycled through phagocytosis by retinal cells (Garcia-Castineiras 2010; Yefimova et al. 2002). Also, retinal ganglion cells have been shown to be able to take up iron and transport it along the axon towards the brain (Moos et al. 2011).
These data suggest that the presence of iron-containing pigment in the retina could also be indicative of normal intraretinal iron transport and turnover.
In conclusion, the focal retinal pigment deposits staining positive for ferric iron is likely to represent hemosiderin accumulation after retinal micro-hemorrhage or could be indicative of the normal intraretinal iron transport and turnover.
Footnotes
Acknowledgments
The authors would like to thank Sarah K. Mortensen and Steen Larsen for critically reading the article.
Author Contribution
Authors contributed to conception or design (AH, LO, LS, KW); data acquisition, analysis, or interpretation (AH, LO, LS, KW); drafting the manuscript (AH, LO); and critically revising the manuscript (LS, KW). All authors gave final approval and agreed to be accountable for all aspects of work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) received no financial support for the research, authorship, and/or publication of this article.