
Abstract
Differentiating salient histopathologic changes from normal anatomic features or tissue artifacts can be decidedly challenging, especially for the novice fish pathologist. As a consequence, findings of questionable accuracy may be reported inadvertently, and the potential negative impacts of publishing inaccurate histopathologic interpretations are not always fully appreciated. The objectives of this article are to illustrate a number of specific morphologic findings in commonly examined fish tissues (e.g., gills, liver, kidney, and gonads) that are frequently either misdiagnosed or underdiagnosed, and to address related issues involving the interpretation of histopathologic data. To enhance the utility of this article as a guide, photomicrographs of normal and abnormal specimens are presented. General recommendations for generating and publishing results from histopathology studies are additionally provided. It is hoped that the furnished information will be a useful resource for manuscript generation, by helping authors, reviewers, and readers to critically assess fish histopathologic data.
Introduction
One of the most valuable skills for the novice anatomic pathologist to acquire is the ability to distinguish morphologic evidence of disease from common background findings, artifacts of slide preparation, and species-specific idiosyncrasies. Generally, that degree of expertise cannot be taught in a matter of weeks to months, nor can it be learned simply by rote through dedicated self-study. Instead, the path to proficiency traditionally requires initial broad-based training in comparative (multispecies) pathology, which is then followed by a steady and diverse caseload, continuing education, and most notably, many years spent laboring at the microscope. However, for the vast majority of scientists who find it necessary to incorporate histopathologic data as a minor (albeit integral) component of their research, an intense level of commitment to the study of histopathology is rarely practical. As a result, histopathologic findings of dubious accuracy may be published by well-intentioned investigators. Ironically, such authors may support their findings with evidence gleaned from prior reports that are equally questionable. In the worst-case scenario, this iterative circular process can create a body of unsound literature that ultimately becomes scientific dogma.
Fish histopathology studies seem inherently prone to this unfortunate sequence of events. One possible reason is that investigators in this field often have widely diverse scientific backgrounds and technical skill sets. For example, professionals who publish fish histopathology findings include aquatic animal biologists, research physicians, ecotoxicologists, anatomic pathologists, veterinary diagnosticians, and fisheries management specialists, among others. A second reason is that grant funding for fish research often favors projects that are geared toward practical application in preference to basic scientific investigation; consequently, there is little impetus to study fish histopathology as a discipline unto itself. A third contributing factor is the paucity (relative to the study of mammalian pathology) of authoritative reference materials and educational opportunities that are dedicated to fish histopathology. Finally, the profound anatomic diversity of fishes, when combined with a myriad of known fish diseases and potential toxicologic exposures, all contribute to the challenge of distinguishing true pathologic changes from the background “noise” of specimen processing artifacts and postmortem autolysis.
The purpose of this article is to pool the expertise of a number of highly trained, veteran fish pathologists in order to provide guidance in the diagnosis (and perhaps more importantly, the nondiagnosis) of commonly encountered morphologic changes in the gills, kidney, liver, and other less frequently examined fish tissues. Some of the information presented in this article will not be supported by citations because applicable references based on unbiased empirical research or historical control data are simply not available. Accordingly, the guidance provided in this document will rely heavily on the consensus opinions of the authors, many of whom have decades of fish pathology experience. More important than the length of their tenure, however, is the fact that these individuals have had the opportunity to examine thousands of disease-free fish over the course of their careers. These include fish devoid of known microbiological infections or toxic exposures (such as those sacrificed during routine animal health screens), fish collected in pristine habitats, and negative control subjects from experimental bioassays and infectious disease trials. It is through the often-tedious work of evaluating untold numbers of “normal” animals that pathologists attain the ability to reliably differentiate between diseased and unaffected tissues. The subjective nature of this reliance on consensus opinion as the gold standard for histopathological accuracy is fully acknowledged; it is no secret that even the most seasoned pathologists are occasionally humbled by errors of diagnostic judgment. However, the authors believe that this limitation is outweighed by the value to be gained from an open discussion of diagnostic and interpretive issues, and the promotion of a judicious approach to the reporting of histopathologic findings.
The Two Most Common Misses
Before we address specific findings, it may be helpful to briefly describe basic terminology that will be used to discuss various diagnostic issues. Over the years, informal reviews by the authors of posters, oral presentations, manuscripts, and published articles have revealed that the two most common “misses” in fish histopathologic reports are misdiagnosis and misinterpretation.
Misdiagnosis occurs when a morphologic observation is considered incorrectly to be abnormal, or when inaccurate or imprecise terminology is used to describe a particular morphologic observation. In such cases, the “correct” diagnosis for a particular lesion should be derived via a weight-of-evidence approach, in which the greatest weight is assigned to compelling results from careful scientific experimentation and valid published observations. Admittedly, assessing the validity of findings in prior articles is often challenging. Alternatively, when legitimate-appearing data are not available, the correct diagnosis defaults to the prevailing consensus opinion of skilled fish pathologists. Experience dictates that the three most frequently encountered causes of misdiagnosis are inadequate tissue fixation, poor histologic slide quality, and a lack of histopathologic expertise.
In contrast to misdiagnosis, misinterpretation involves the derivation of inappropriate conclusions from morphologic findings that have been identified and described correctly. Misinterpretation of histopathologic data can take multiple forms. One form occurs when the results from one experimental group of fish are reported to be different from those of another group, when, in actuality, no valid significant differences exist. This can occur, for example, when experimental control animals are presumed to be normal, and thus receive a lesser degree of diagnostic scrutiny as compared to compound or disease challenged animals. Of course, the reverse situation is also possible; that is, there can be a failure to detect groupwise differences that are actually present. Weaknesses in the experimental design or surveillance sampling protocol can predispose to this form of error (e.g., too few animals per comparison group or collection site, too few replicates, and/or failure to control certain independent variables), or errors may arise because sufficient care was not taken to minimize bias during sample collection or during the slide evaluation process. A second type of misinterpretation involves the premature assumption of a cause-and-effect relationship without adequate corroborating evidence in the experimental data or scientific literature. This often stems from a failure to consider alternative explanations for the induction of a particular histopathologic effect. A third form of misinterpretation occurs when a treatment-related finding is afforded an exaggerated degree of biological, pharmacodynamic, toxicological, or ecological importance that is not supported by existing scientific evidence. Recommendations for avoiding misdiagnosis and misinterpretation will be addressed later in this article.
Findings That Are Frequently Overdiagnosed
The fish histopathology literature features a number of morphologic diagnoses that seem to appear with inordinate frequency. Table 1 presents examples of commonly reported diagnoses from a diverse array of histopathologic investigations. Often, such diagnoses are particularly overrepresented in studies that involve intentional (experimental) or suspected toxicological exposure, as opposed to those that deal primarily with the characterization of infectious disease, for example. One obvious and quite reasonable explanation for this phenomenon is that fish tissues are capable of reacting to adverse stimuli in only a finite number of ways. Among fish and other vertebrates, morphologic manifestations of inflammation, degeneration, necrosis, atrophy, and regenerative proliferation are hallmark responses of living tissues to cytotoxic damage and/or physiological stress. Consequently, histopathologic changes that result from exposure to biological pathogens or noxious chemical agents are rarely novel or cause-specific. Furthermore, it is important to note that essentially all of the diagnoses presented in Table 1 can be legitimate pathological findings in many instances (including at least some of those particular studies). However, there also appear to be numerous reports in which the incorporated photomicrographs, morphologic descriptions, and lesion occurrence data suggest that the histopathologic findings were misdiagnosed and/or misinterpreted in the context of the publications in which they appeared.
Suites of commonly reported histopathologic findings from toxicological studies.
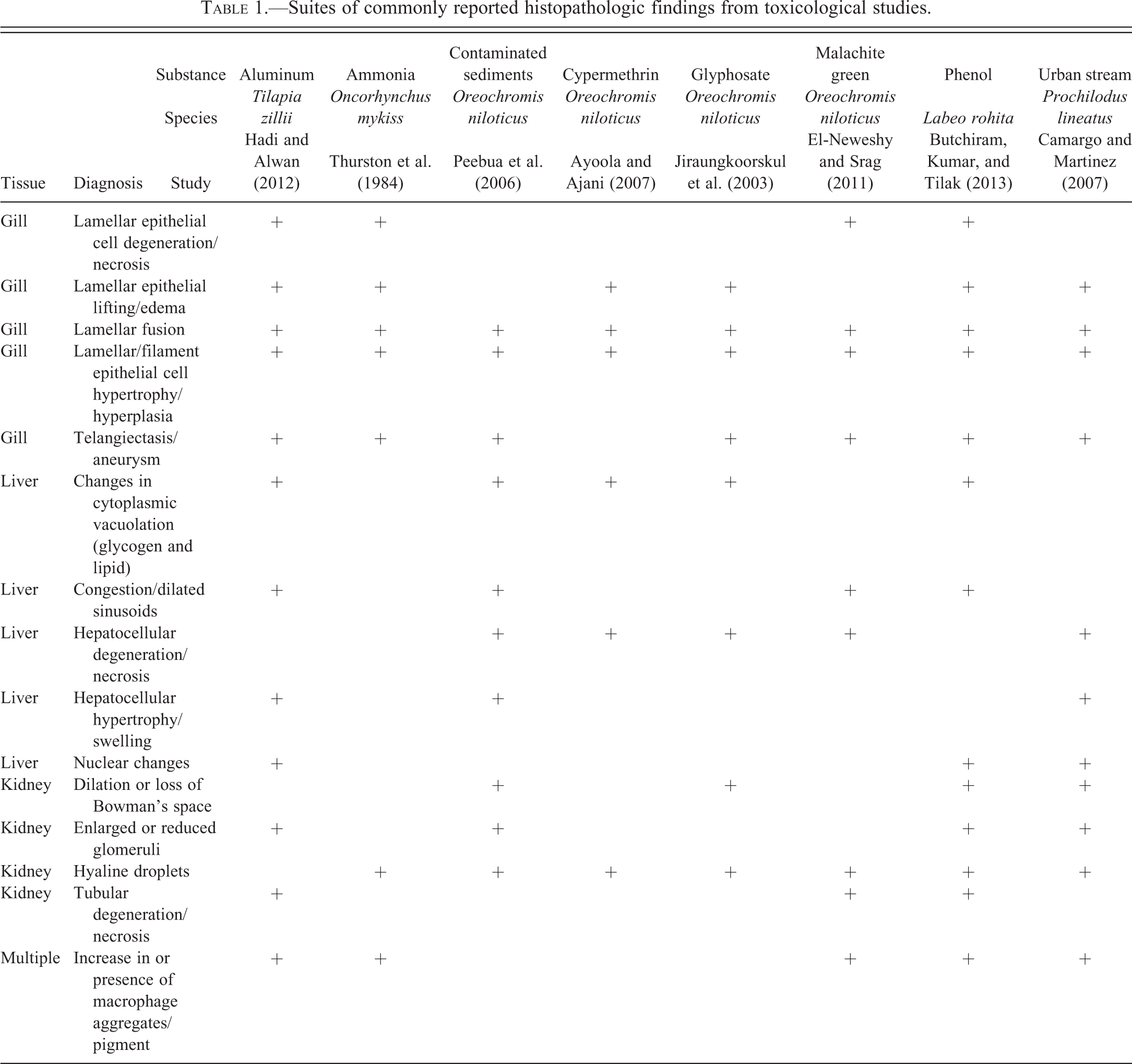
Table 2 presents an expanded catalogue of findings that tend to be over- and underdiagnosed, based on experience of the authors of this article. The following discussion will concentrate on these general types of pathologic responses, as opposed to specific concerns such as the identification of infectious agents or neoplastic lesions. Regarding the latter, standardized diagnostic criteria have been established for certain categories of fish neoplasms (e.g., Fournie and Vogelbein 1994; Boorman et al. 1997; Fournie and Hawkins 2002; Fournie et al. 2005; Blazer et al. 2006), although further guidance is also needed in that area.
Examples of over- and under-diagnosed findings.

Note. IP = intraperitoneal.
Gills
It is easy to understand why the gills, liver, and kidney are the most frequently examined tissues in fish histopathological evaluations. All three organs have high metabolic activity and together they bear heavy responsibility for the biotransformation and elimination of noxious substances of both endogenous and exogenous origin. However, because the gills are directly exposed to the aquatic environment, they are exceptionally vulnerable to an assortment of traumatic, toxicological, and infectious insults. Fortunately for fish, the regenerative capabilities of gill tissue are nothing short of astonishing, as even severely injured gills are capable of complete recovery within a matter of days to weeks, once the cause of the problem has been alleviated. This recuperative ability is seldom acknowledged when authors discuss the implications of experimentally induced gill findings in published reports; such failure to account for recoverability may cause investigators to overestimate the long term impacts of certain gill lesions on the health of individual fish and fish populations.
Of the three aforementioned tissues, the gills are the most technically challenging to prepare histologically. As luck would have it, the gills are also the tissue for which optimal histologic preparation is the most important. Experience demonstrates that poor preparation is a major cause of both misdiagnoses and missed diagnoses. The generation of diagnostic quality gill sections requires the use of suitable histologic methods for specimen collection, preservation, and orientation for sectioning.
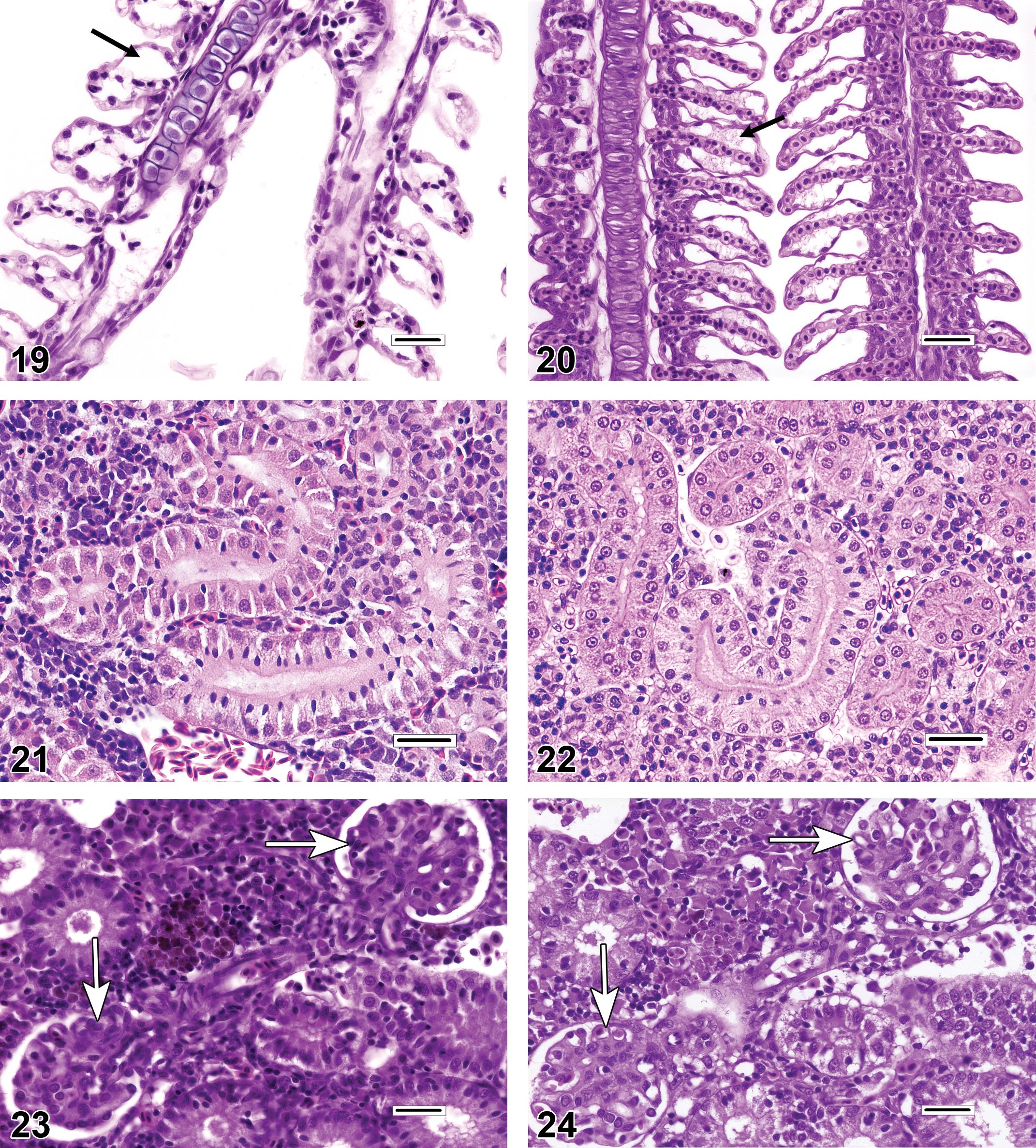
The choice of initial fixative solution for the gills is key. While 10% neutral buffered formalin (NBF) is an excellent general histologic fixative that is relatively inexpensive and readily available, it is arguably not the best option for obtaining optimal results from certain fish tissues, such as the gills, kidney, and testis. Problems with NBF-fixed gills can include excessive condensation of nuclei and other structures, and separation artifacts (see epithelial lifting) that create artifactual spaces in the tissue (Speare and Ferguson 1989). Superior alternatives for initial fixation include modified Davidson’s solution, Dietrich’s solution, and Bouin’s solution (Speare and Ferguson 1989; Fournie, Krol, and Hawkins 2000). Because they contain acetic acid or picric acid, respectively, tissues fixed in modified Davidson’s or Bouin’s solutions should be rinsed thoroughly in 70% ethanol after 24 to 48 hr, following which they can be stored in NBF. One benefit of these acid-containing fixatives is that they may partially or completely soften bony and cartilaginous structures, depending on the specimen size. For further information on fixatives and fixation, readers are encouraged to consult Fournie, Krol, and Hawkins (2000). Gills from large fish may require additional decalcification prior to routine histologic processing for paraffin embedding, and commercially prepared formic acid/ethylenediaminetetraacetic acid (EDTA) combinations seem to work well for this application. It is also imperative that the gills be oriented in the embedding medium (e.g., paraffin), in a manner that allows for optimal examination of the specimens. Ideally, gill specimens should be positioned so that microtome cuts can be made as parallel as possible to the long axis of both the filaments (primary lamellae) and the lamellae (secondary lamellae). To accomplish this in large fish, it may be necessary to transect the gill arches into smaller, more manageable portions. Postmortem autolysis occurs rapidly in the gills and other fish tissues. If appropriate fixation is delayed, such as when collected specimens are placed in water or on ice for more than 5 or 10 min following fish sacrifice, then the quality of the resulting histologic preparation is likely to suffer, and thereby induce preservation artifacts that may confound the microscopic evaluation (Figure 1). For fish preserved whole, it is recommended that the gill cavity be flushed with fixative prior to immersion of the carcass in fixative, in order to enhance preservation.
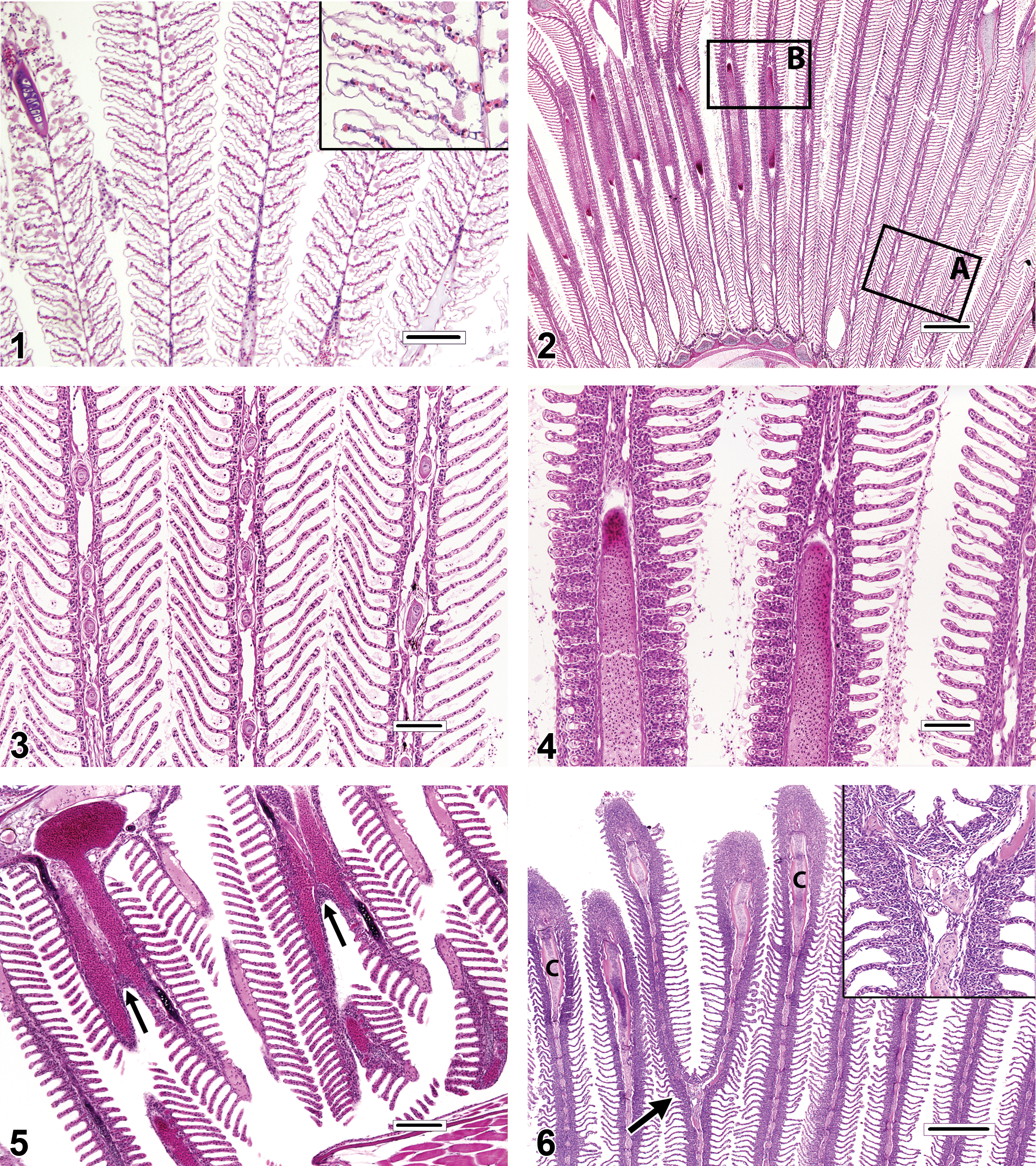
Gills destined for histopathologic evaluation can be either excised from the carcass as whole gill arches or individual filaments, or in the case of small fishes, examined in whole-body sections. Excised gill specimens are best obtained as intact gill arches, and sampling of the first gill arch is often avoided because it is thought that those filaments may bear the brunt of particulate matter exposure. Fish gills have a complicated 3-dimensional architecture that can be difficult to appreciate in 2-dimensional histologic sections. When evaluating histologic sections of gill tissue, diagnostic determinations should be based, as much as possible, on filaments in which the lamellae are of equal length and maximum extension. Filaments meeting those criteria are thus oriented in a manner that allows for the accurate and consistent appraisal of pathological changes. Conversely, the evaluation of filaments with uniformly truncated lamellae, or those in which the lamellae are much longer on one side of the filament than the other, should be avoided. This is because incorrectly positioned filaments will inevitably appear more proliferative than filaments that are oriented properly (Figures 2–4). Evidence from published photomicrographs indicates that the failure to follow this approach can lead to inappropriate diagnoses of lamellar fusion, lamellar epithelial hyperplasia (LEH), lamellar loss or atrophy, chloride cell hyperplasia, mucous (goblet) cell hyperplasia, and gill inflammation (branchitis).
Unless fish are very small, in which case gill excision might cause excessive iatrogenic damage, excised specimens are usually preferred for the accurate assessment of gill changes. However, for juvenile fish or diminutive fish species, it may be necessary or expedient to examine the gills in situ in whole-body sections. Evaluation of the gills using this approach can be challenging, because they are invariably microtomed along a plane that is tangential to the long axis of the filaments, and this can create 3-dimensional distortions that may be misdiagnosed as pathologic findings. For example, evidence from published photomicrographs suggests that some reported occurrences of filament fusion and/or branching are actually optical illusions caused by the superimposition of neighboring filaments (Figure 5). By contrast, true filament branching (Figure 6) is a relatively rare finding, potential causes of which include congenital gill defects, myxozoan infections of the gill cartilage, and environmental contamination. Gills sectioned obliquely in whole-body sections also tend to appear artificially thickened and hypercellular (Figures 7–8) when compared to gills that have been excised.
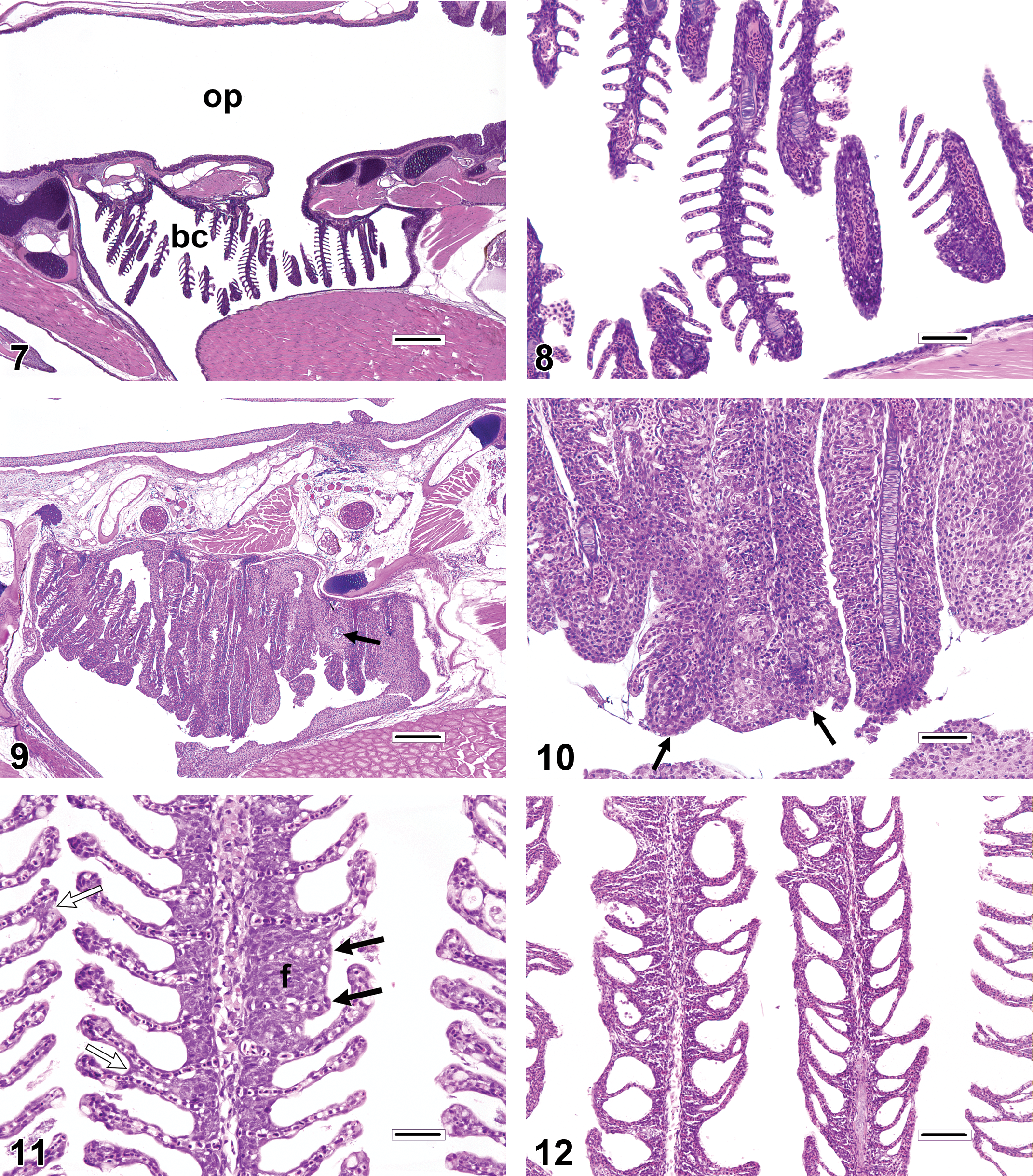
The gills have a limited repertoire of pathologic responses to various chemical or physical injuries. A common, nonspecific response to subacute to chronic gill damage is proliferation of the squamous epithelial cells that line the gill surface (i.e., pavement cells). This change is typically termed LEH (Figures 9–12). In addition to multiple layers of pavement cells which are often enlarged, gills with LEH may feature increased numbers of mucous cells, chloride cells, and/or inflammatory leukocytes as components of the thickened respiratory epithelium. LEH may serve to safeguard the fragile lamellar capillaries from further external damage or microbial penetration; however, it also increases the diffusion distance between the capillaries and the environment and thus may hamper respiratory, excretory, and osmoregulatory functions. A frequent consequence of minimal to advanced LEH is focal or diffuse lamellar fusion (Figures 9–11). Lamellar fusion is a term that is used when one or more interlamellar sulci become partially to completely filled by proliferating pavement cells, with or without increased mucous cells, chloride cells, and/or inflammatory leukocytes. Filament fusion (Figure 10), which involves the partial or complete attachment of adjacent gill filaments, is a much less common finding than lamellar fusion. Actual filament fusion usually only occurs secondary to severe LEH. It can be useful to differentiate lamellar fusion from lamellar adhesions(synechia; Figures 11–12), in which neighboring lamellae become focally attached at one or more mutual points along their lengths (often at the tips), because the latter finding can be a more specific indicator of pavement cell necrosis. Unlike lamellar fusion, the formation of adhesions does not require epithelial cell proliferation. True lamellar loss or atrophy (Figure 11) is an uncommon finding that most often affects only scattered individual lamellae, as opposed to those on the entire side of a filament, for example. In many fish species, the basilar areas of gill filaments, gill arches, and pharyngeal mucosae may contain dense populations of resident lymphocytes and/or eosinophilic granular leukocytes (Figure 13), and it is tempting to diagnose such constituent infiltrates as gill inflammation (branchitis). However, it should be recognized that actual branchitis tends to involve the distal two-third of the filaments in addition to the more proximal regions and that other features of inflammation (e.g., necrosis of pavement or endothelial cells, LEH, and/or vascular congestion) often accompany the leukocytic infiltrates in such cases. Chloride cells are specialized osmoregulatory cells that tend to be more numerous in marine or euryhaline fishes. These plump, slightly hypereosinophilic cells are typically located within interlamellar sulci, and chloride cell hyperplasia (Figure 14) involves proliferation of these cells along the lengths of lamellae in a basal to apical direction. Mucous (goblet) cells can be mistaken for chloride cells. Most teleost gills contain only low numbers of mucous cells, especially along lamellae, and the morphology of these cells tends to vary by species. Mucous cell hyperplasia (Figure 15) generally occurs secondary to persistent gill irritation.
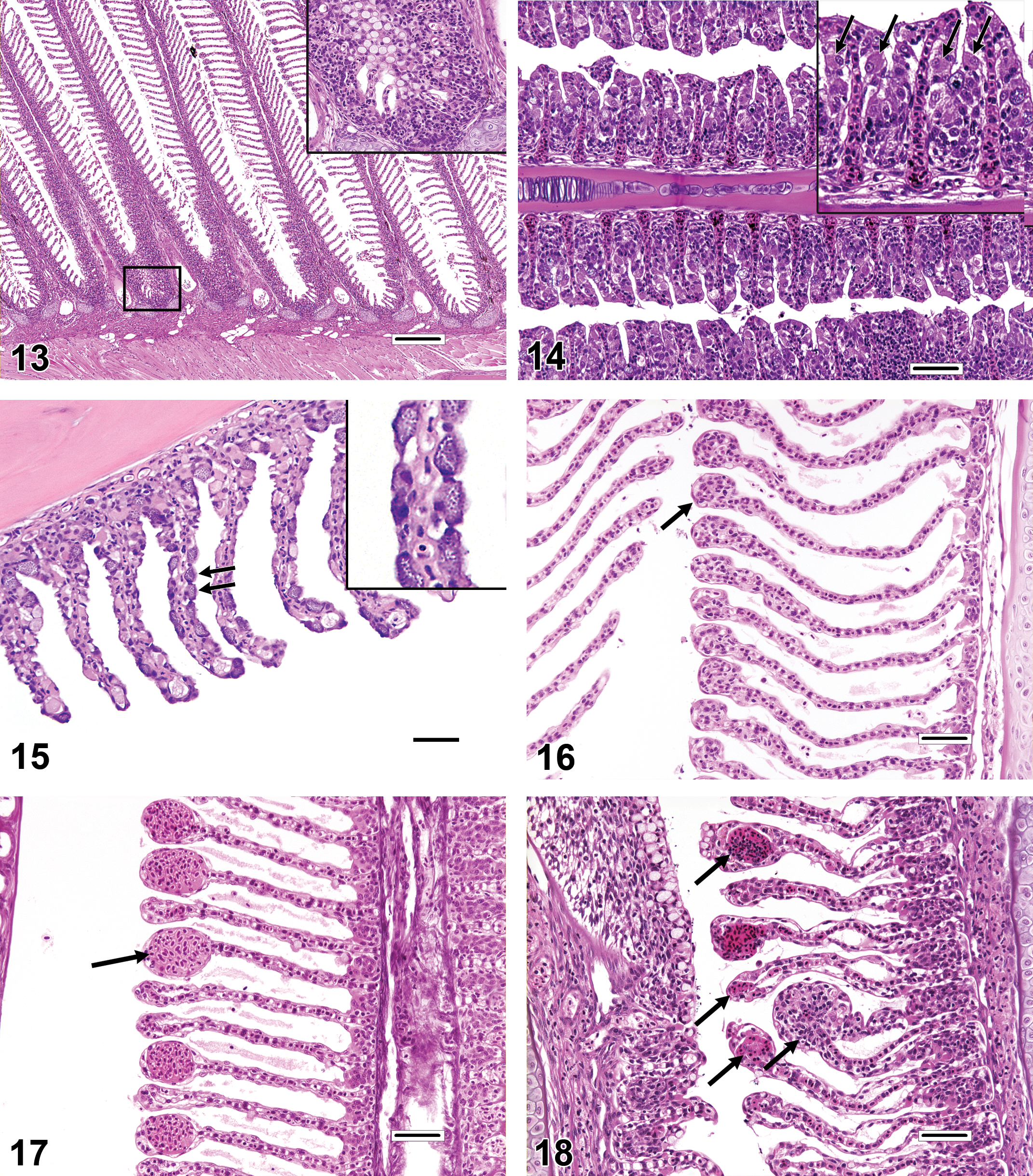
A diagnosis that can be simulated by the malposition of gill filaments is lamellar clubbing. Lamellar clubbing (Figure 16) refers to nonspecific, bulbous enlargements of the distal ends of the lamellae, and causes of this can include focal epithelial (pavement) cell proliferation, focal lamellar inflammation, and resolving lamellar thrombosis. Conversely, artifacts resembling mild lamellar clubbing are frequently evident in sections that are not oriented parallel to the long axis of the filaments; such artifacts should not be reported as pathologic findings. Lamellar clubbing should not be confused with filament clubbing, which is characterized by lamellar epithelial cell proliferation and fusion that is most prominent near the apical tips of filaments (Figure 6).
Two commonly reported gill diagnoses that are often credited with far more significance than they deserve are lamellar telangiectasis and lamellar epithelial lifting/edema. Although both telangiectasis and gill edema can be bona fide antemortem findings, they seem to occur most commonly as artifacts induced by procedures that take place at or around the time of death. Typically, telangiectasis (telangiectasia, angiectasis, and lamellar aneurysm; Figure 17) is readily identified as blood-filled, ovoid expansions of individual lamellar capillaries. Anecdotal causes of telangiectasis include head concussion and ice water bath sedation, among other euthanasia-related procedures. Because large telangiectatic lesions generally resolve by lamellar thrombosis (Figure 18), findings of thrombosis can provide evidence that telangiectatic lesions were actually present in the gills prior to the necropsy period. Conversely, a lack of thrombotic changes suggests that the duration of telangiectasis was peracute; in such cases, telangiectasis may not represent a true pathologic finding. Epithelial lifting (Figure 19) refers to lamellae in which a noticeable gap has developed between pavement cells and lamellar capillaries. Such gaps may appear completely clear, or they may contain flocculent, pale pink, proteinaceous material, consistent with interstitial lamellar edema (Figure 20). Potential causes of epithelial lifting and lamellar edema, respectively, can include suboptimal fixation procedures and poor water quality (e.g., fish placed in small containers for extended periods of time while awaiting sacrifice; Fournie, Krol, and Hawkins 2000).
Kidney
Similar to the gills, the histologic preparation of fish kidneys can be problematic. The piscine kidney is fragile and easily damaged during postmortem collection. In certain fishes (e.g., centrarchids), the kidney exists as a narrow, flat band of tissue located immediately subjacent to the spine; in those species, the acquisition of undamaged samples may require excision of the spine in conjunction with the kidney. Also like the gills, formalin fixation (Figure 21) often generates inferior results (artifactual spaces between tubular epithelial cells and pyknotic-looking nuclei) when compared to kidneys fixed in Davidson’s or Bouin’s solutions (Figure 22). Histologic sections that are excessively thick (e.g., greater than 4–6 microns) can appear falsely hypercellular with dense-looking glomerular tufts (Figures 23–24), and the plasma within renal vessels may have an artificially proteinaceous (dark pink) appearance.
The piscine kidney typically contains components of several physiological systems, including tissues with hematopoietic, immunologic, endocrine, and urinary functions. The most commonly examined components for evidence of toxicological disturbances or inflammatory disease are the urinary glomerulotubular structures (= mesonephros = opisthonephros) and the renal interstitium. In most teleosts, the anterior portion of the kidney (i.e., the cranial or head kidney) is exclusively hematopoietic, while the remaining posterior portion (caudal or trunk kidney) primarily consists of urinary tract tissues. Other fishes (e.g., guppies Poecilia reticulata and zebrafish Danio rerio) have urinary tissue distributed throughout the entire length of the organ. Physiologically, the primary role of the urinary kidney is osmoregulation, as the gills can be responsible for greater than 80% of nitrogenous waste excretion (Evans, Piermarini, and Choe 2005). Additionally, the piscine kidney typically contains (or is intimately associated with) endocrine tissues such as interrenal and chromaffin cells (Figure 25), which are analogues of the adrenal cortex and medulla, respectively, and the bilateral corpuscles of Stannius (Figure 26), which secrete the calcium-regulating hormone stanniocalcin.
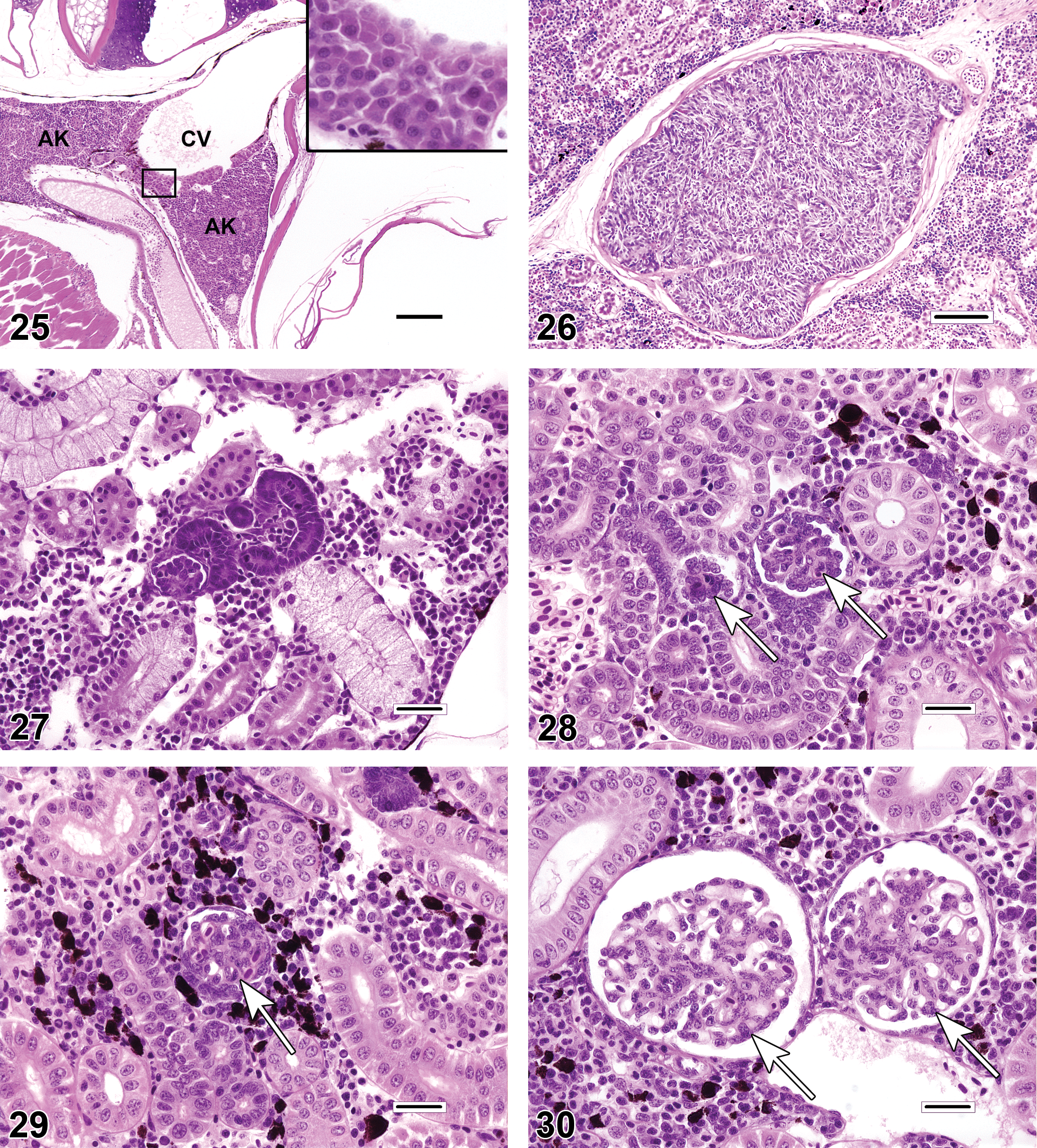
There are fundamental anatomic and physiologic differences between the teleost mesonephros and the metanephric kidney of higher vertebrates. Anatomic features of the teleost mesonephric kidney include left and right kidneys that are predominantly conjoined, undefined cortical and medullary regions, fewer nephrons than the metanephric kidney, species-specific reductions in certain nephron segments (e.g., the kidneys of seahorses Hippocampus spp. and toadfish Opsanus spp. lack glomeruli), and the ability to regenerate nephrons (nephron neogenesis) in both juvenile and adult teleosts. Morphologically, nephron neogenesis (Figure 27) is characterized by the presence of one or more immature, dark basophilic nephron segments (tubules and/or glomeruli) that are commonly observed in overtly healthy kidneys as one or more scattered individual structures. Because of the basophilic, embryonic appearance of these regenerating tissues, they could potentially be mistaken for tubular mineralization or incipient neoplasia. There is evidence that numbers of regenerating nephrons may increase in response to renal injury (Reimschuessel 2001).
Compared to mammals, fish glomeruli tend to be somewhat pleomorphic; an otherwise normal kidney can exhibit substantial variation in the size and cellularity of glomerular tufts, diameter of capillary loops, thickness of the mesangium, and in the volume of Bowman’s space (Figures 28–30). Some of this variability can be attributed to differences in nephron maturity as a result of nephron neogenesis. Additionally, glomerular features, and renal morphology in general, may differ according to the region of kidney sampled. Care must be taken to account for pleomorphism during histopathologic evaluations, so that normal variation will not be misdiagnosed as a pathologic finding. Frequently, descriptions of published reports do not contain sufficient detail for the reader to determine that the kidneys were sampled consistently according to standardized procedures (e.g., “1 cm long specimens of kidney were excised ventral to the nth vertebra … ”). Ultrastructural evaluations may aid in the characterization of glomerular or tubular changes, but because only minute amounts of tissue are evaluated, electron microscopy is not the best tool for investigating comparative differences among individual fish or groups.
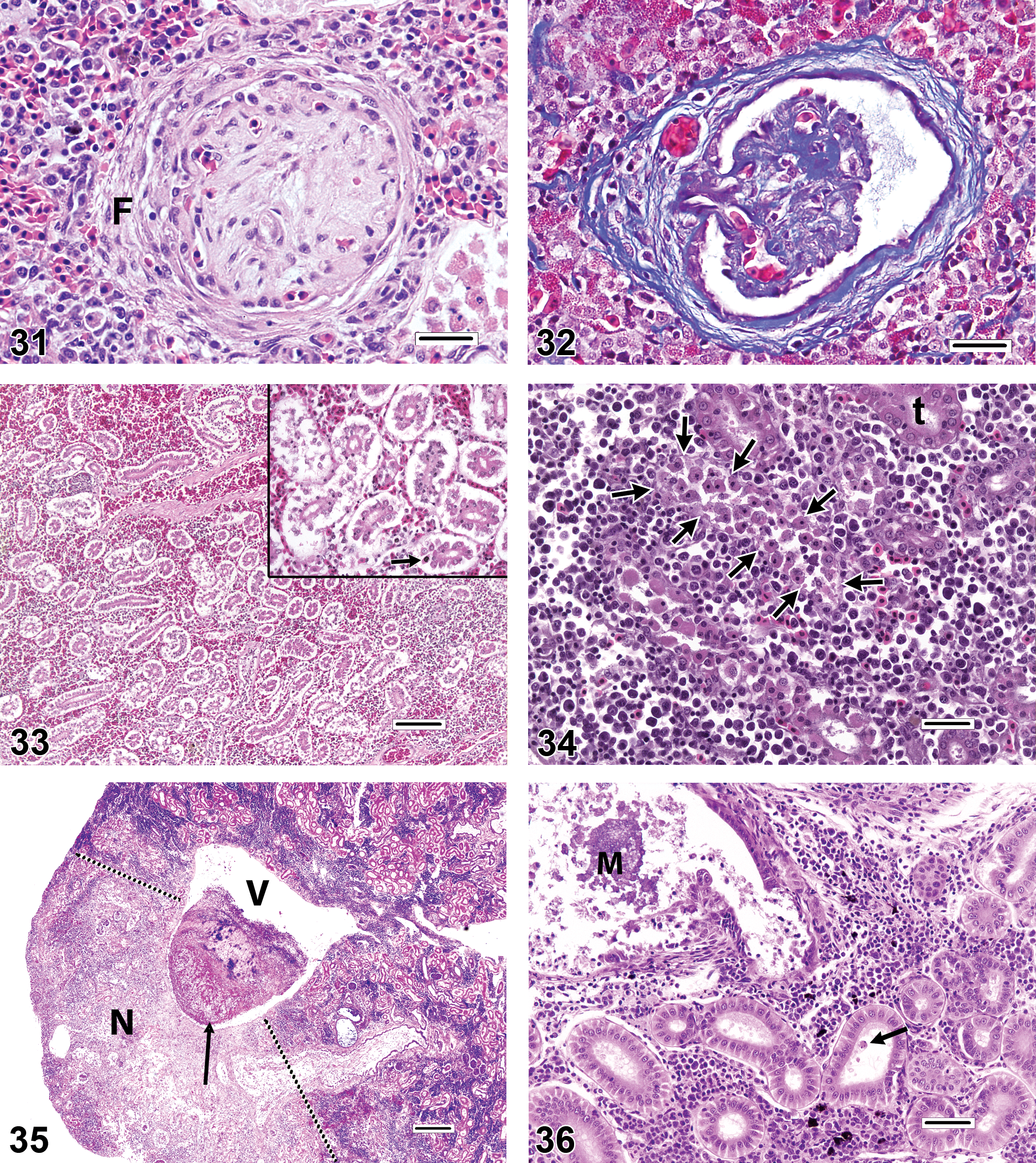
As discussed previously, care must be taken to account for pleomorphism during histopathologic evaluations, so that normal variation will not be misdiagnosed as a pathologic finding. On the other hand, two genuine glomerular changes that are seen with some regularity in certain fishes are periglomerular fibrosis and glomerulosclerosis (Figures 31–32). The authenticity of these findings can be verified through the use of special stains that highlight collagenous tissue (e.g., trichrome stain).
In certain fish species, and more often in males, the proximal tubular epithelial cells may have unique cytoplasmic characteristics, such as hypereosinophilia, granular texture, the presence of mucoid material (e.g., zebrafish), or increased cytoplasmic volume (e.g., reproductively active male three-spined sticklebacks Gasterosteus aculeatus). Pathologists who do not become familiar with such species-specific characteristics risk reporting them incorrectly as pathologic diagnoses. Sex-related differences in the size and coloration of proximal tubular epithelial cells can be a confounding factor for studies in which the fish gender was not determined or reported.
Next to the gills, the kidney is perhaps the most overdiagnosed organ in fish histopathology. The most commonly misdiagnosed findings in the kidney are tubular degeneration and tubular necrosis. Based on published photomicrographic figures, it seems that at least some incorrect findings of degeneration and/or necrosis can be attributed to autolytic changes, whereas in other instances, tubular loss may be diagnosed erroneously in specimens collected from kidney regions where the tubules are normally sparse (i.e., at or near the anterior kidney). In autolyzed kidney sections (Figure 33), renal tubular epithelial cells may have ragged-looking edges and condensed nuclei, and there may be some degree of intercellular dissociation; however, the general architecture of urinary structures is generally maintained, there are few if any karyorrhectic tubular cells, and downstream tubules typically do not contain large numbers of exfoliated cells or casts. The appearance of actual tubular necrosis can vary considerably, but three of the most common presentations in fish kidneys include individual cell necrosis, necrosis involving whole tubules, and extensive necrosis. Individual cell necrosis (apoptotic necrosis) is characterized by one or more scattered cells that have condensed into heterochromatic spheres within the tubular epithelium. Each cell is typically surrounded by a thin halo of clear space. It is not unusual to find occasional individual necrotic cells within the tubules of an otherwise healthy kidney. In whole-tubule necrosis (Figure 34), which is often of the coagulative type, the tubular epithelial cell cytoplasm is slightly hypereosinophilic and refractile, with irregular margins, and the nucleus is pyknotic or karyorrhectic. Coagulated cells often exfoliate into the tubular lumen, although by itself, the observation of occasional sloughed cells in tubular lumina does not automatically warrant a diagnosis of necrosis. Extensive necrosis, as observed in renal infarcts caused by systemic fungal infections, for example, is characterized by distinctly outlined regions of pale, moth-eaten, glomerulotubular tissue that may have a narrow zone of inflammation at the border between affected and less affected tissue (Figure 35). Renal mineralization (Figure 36) seems to be a fairly common cause of tubular necrosis that may be underrecognized. In undecalcified H&E-stained sections, mineralization typically presents as purple spherical or irregular concretions within renal tubules (especially the distal tubules or collecting ducts) or the interstitium, or as granular purple discoloration and fragmentation of the tubular epithelium itself. Whether intraepithelial or intraluminal, the physical presence of mineralized material within tubules is often sufficient to cause localized damage, which may be accompanied by tubular dilation and epithelial attenuation as a result of obstruction. Often, the mineralized material is not present in every histologic section; in such instances, it is possible to misinterpret the cause of the tubular necrosis as something other than mineralization. It is reasonable to presume that the cause of mineralization in captive fish is probably husbandry related (e.g., dietary or water quality issues) in most cases; however, renal mineralization is also observed occasionally in wild fishes.
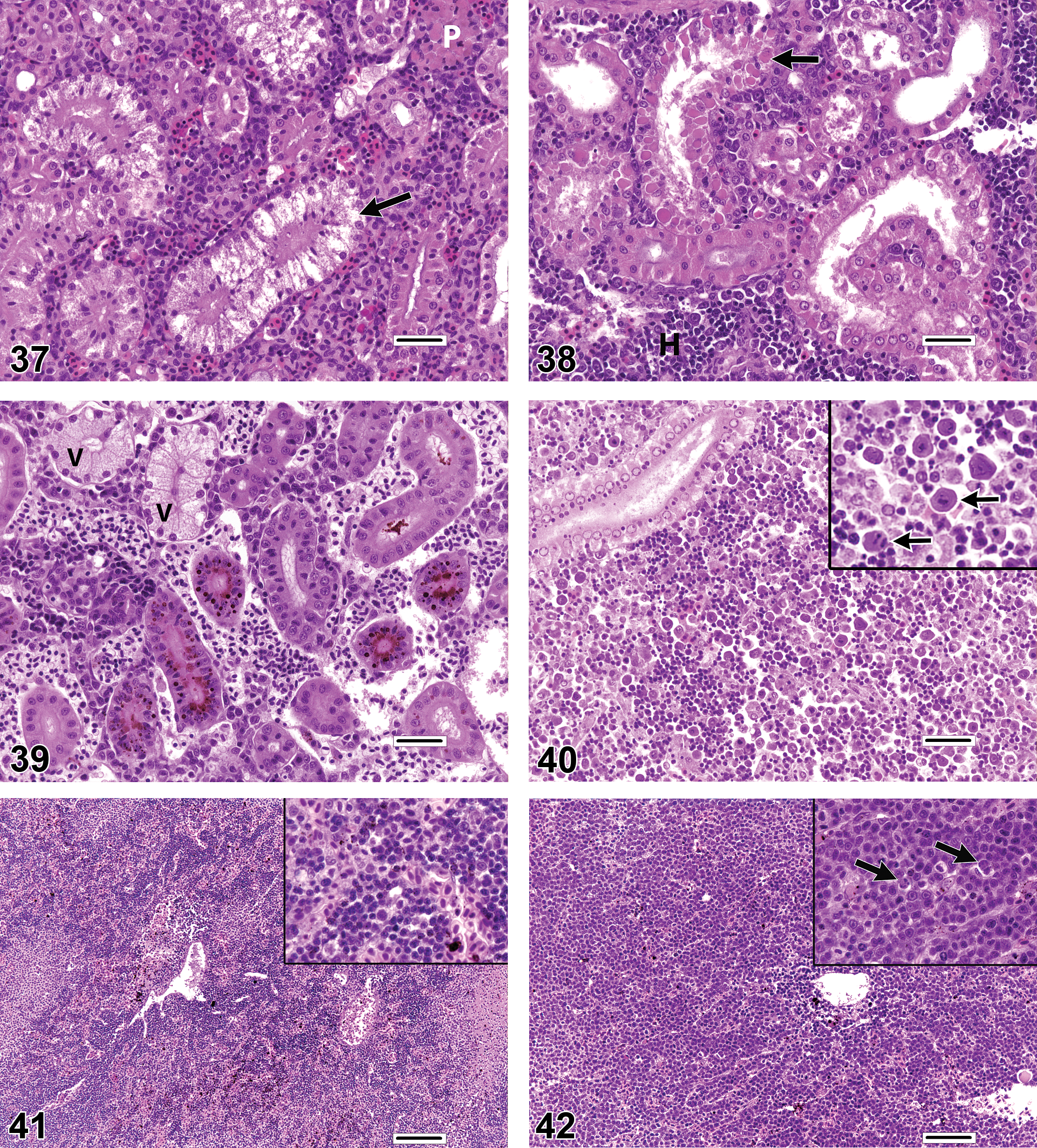
The appearance of renal tubular degeneration can also be quite variable, but common general categories include vacuolar degeneration, hyaline degeneration, and pigment accumulation. Vacuolar degeneration (Figure 37) of tubular epithelial cells can present as single or multiple clear areas within the cytoplasm. The edges of vacuoles may be sharp (lipid-like) or indistinct, and the spaces may be entirely clear or contain faint flocculent material (e.g., cloudy swelling). Hyaline degeneration (Figure 38) is characterized by the intracytoplasmic presence of refractile spherical eosinophilic droplets or irregularly outlined coagula. In the latter case, hyaline degeneration can be differentiated from coagulative necrosis by the relative lack of adverse nuclear changes in degenerating cells. Pigment accumulation (Figure 39) can manifest as fine or globular, brown or olive green granules within the renal tubular cytoplasm. Because the appearance of pigments in H&E-stained sections of kidney and other tissues can be affected by various factors related to tissue preservation and slide preparation, investigators should avoid making conclusive statements about the chemical nature of the pigment based on the color alone, and instead are advised to use special histologic stains for iron, bile, melanin, lipofuscin, ceroid, and so on, to help determine the precise underlying substance (for further reference, see Bancroft and Gamble 2008).
It should be stressed that minor degenerative changes may be present to a limited degree in glomeruli or tubules of seemingly healthy fish that have had no known exposure to toxic substances or pathogens. On the other hand, such renal findings may attain pathologic importance if they are observed in excess of what would be considered typical for a particular demographic group of fish (e.g., as defined by species, sex, and age) or when the prevalence and/or severity of these findings are increased as compared to conspecific controls or reference site fish.
Interstitial areas of the piscine kidney normally contain erythrocytic, thrombocytic, and leukocytic precursors in various developmental stages, plus mature leukocytes. Additionally, pigmented macrophage aggregates (PMA, melanomacrophage centers) exist constitutively in most teleost kidneys.
Typically, hematopoietic cell necrosis (Figure 40) is usually of the individual cell (apoptotic) or lytic variety, with the latter occurring as a feature of some viral infections, whereas extensive necrosis may be caused by infarcts. A change in the renal interstitium that may be underdiagnosed (based on the scarcity of published reports) is proliferation (hyperplasia) of the hematopoietic elements. Hematopoietic tissue hyperplasia (Figures 41–42) can be a compensatory response to the increased demand created by localized or systemic inflammation, a regenerative response to anemia, or a sequela of direct hematopoietic cell toxicity. In any event, this change is characterized by expansion of the interstitium by a population of leukocytic and/or erythrocytic cells in which the proportion of immature cell types (e.g., blast cells) appears increased relative to the kidneys of control fish, or compared to expectation of normality if controls are not available. Blast cells are typically larger than mature leukocytes and erythrocytes, and they generally have larger, rounder nuclei with more distinct nuclear membranes. Mitotic figures may be observed with increasing frequency as the severity of hyperplasia progresses. Care must be taken not to mistake this proliferative reaction for renal inflammation, which is most commonly either granulomatous (i.e., comprised primarily of epithelioid macrophages) or granulocytic (i.e., consisting primarily of mature neutrophils and/or eosinophilic granular leukocytes). Hematopoietic tissue hyperplasia must also be distinguished from relatively rare neoplastic disorders such as lymphoma. Proliferating hematopoietic tissue may cause renal tubules to spread apart from each other, and it is a common cause of renomegaly. However, because tubule density tends to vary along the axial length of the kidneys, determinations of hematopoietic tissue hyperplasia are most reliable when the specimens are collected in a standardized manner and a series of kidney sections can be examined.
Because of the high degree of resident cellularity in the normal renal interstitium, the identification and characterization of renal inflammation (interstitial nephritis; Figures 43–44) can be challenging. The most easily appreciated form of interstitial inflammation is the granulomatous type. Granulomatous inflammation can present as discrete spherical granulomas (with or without necrotic centers or peripheral sheaths of flattened macrophages) or as diffuse nodular lesions that consist of pale epithelioid macrophages (i.e., cells with large, eccentric, open-faced nuclei). Granulomatous lesions may be found in random interstitial locations or they may expand the marginal zones of PMA. Lymphoplasmacytic and granulocytic inflammation may also occur in the renal interstitium but are less common and can be more difficult to differentiate from hematopoiesis.
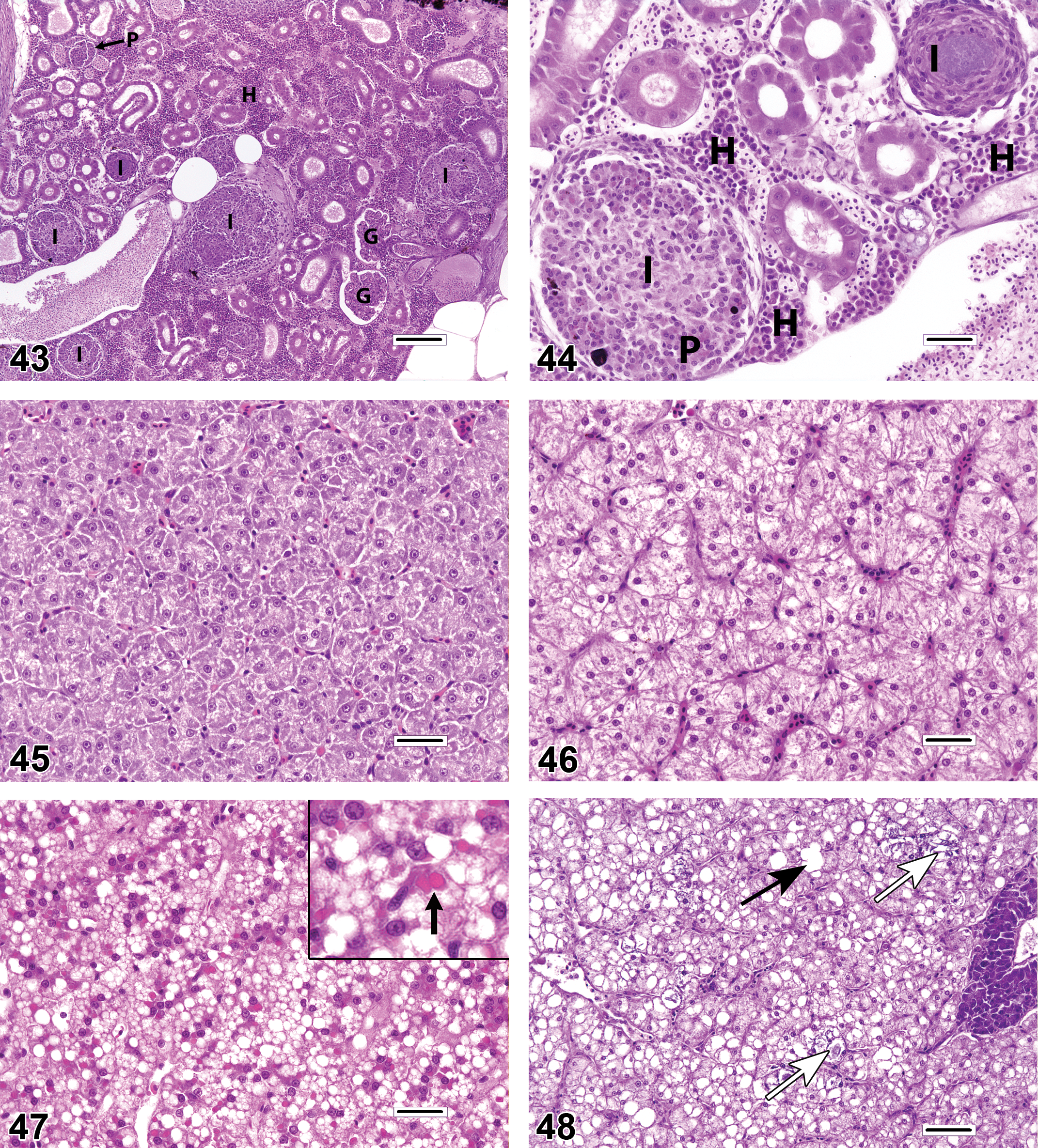
Liver
Compared to the gills and kidney, the histologic preparation of the fish liver is less demanding. A variety of fixatives will generate suitable results, and proper orientation in the paraffin block is usually not critical. That being said, structures such as PMA and bile ducts are not distributed uniformly throughout the liver, and studies should be designed to standardize specimen collection as much as practicable. Additionally, adequate assessment of the liver in large fish (e.g., >1 kg) often necessitates the acquisition of multiple samples from different areas of the same liver specimen.
The coloration, texture, and relative amount of hepatocyte cytoplasm in an individual fish may be influenced by a variety of factors such as species, age, gender, reproductive phase or season, nutritional status, and effects of toxic or inflammatory disease. In reproductively active adult females of oviparous species, upregulated synthesis of the egg yolk protein vitellogenin often causes the hepatocyte cytoplasm to have a mottled, basophilic appearance (Figure 45). When energy intake exceeds the demands of metabolism, growth, exertion, reproduction, and other physiologic requirements, the piscine liver tends to store a portion of the excess energy as glycogen and/or lipid in the cytoplasm of hepatocytes. This is especially common in captive-reared fish, which are typically fed calorically dense foods and do not have to expend much energy foraging. Glycogen and lipid storage manifests visually as hepatocyte cytoplasmic vacuolation in histologic sections. Vacuoles that contain slightly flocculent material and have indistinct margins are considered consistent with glycogen (Figure 46), whereas lipid generally appears as single or multiple clear round vacuoles with sharp edges (Figure 47). The predilection to accumulate glycogen versus fat appears to be somewhat species dependent and may also be influenced by diet. Arguably, the most overdiagnosed finding in the fish liver is hepatic lipidosis. In human and veterinary medicine, the term lipidosis is frequently defined as a congenital or acquired disorder of lipid metabolism. However, this term has been used somewhat arbitrarily to indicate fish that merely exhibit abundant liver lipid (e.g., in commercially reared fish or aquarium specimens), whether or not a pathologic condition truly exists. For example, excessive-appearing amounts of lipid are considered “normal” for the livers of some fishes, including many elasmobranchs and teleosts such as Atlantic cod Gadus morhua. Because the point at which hepatic lipid accumulation becomes deleterious to fish is currently indeterminate and probably quite variable, it is prudent to avoid the use of the term lipidosis, unless there is visible (or functional) evidence of adverse histopathological effects in the liver, such as cell membrane rupture or saponification (Figure 48). A more appropriate descriptive diagnosis for fish with fat laden livers is “increased lipid-like vacuolation.” Alternatively, some diagnosticians may elect to score the degree of glycogen or lipid vacuolation in all fish according to a semiquantitative scale (e.g., minimal, mild, moderate, and severe). This approach may be advantageous because sick, stressed, or malnourished fish frequently exhibit a visible reduction in hepatic energy stores, and may thus have decreased hepatocellular vacuolation. The major problem associated with the indiscriminate use of the term lipidosis is that various disease effects (i.e., morbidity and mortality) may be attributed erroneously to an observation (i.e., ample hepatic lipid) that is essentially nonpathologic. Hepatocellular hypertrophy (Figure 49) is observed in fish livers on occasion and this change is characterized by enlargement of the hepatocyte cytoplasm, with or without a concomitant increase in nuclear size. Diffuse hypertrophy is most often associated with toxic exposures, although megalocytic hypertrophy can also occur as a feature of regenerative hyperplasia following substantial liver damage (Wolf and Wolfe 2005). In such cases, hypertrophy may be accompanied by increased numbers of multinucleated hepatocytes, plus proliferations of biliary cells and pluripotential oval cells. Diffuse hepatocellular hypertrophy may be difficult to appreciate in single fish because there is little basis for comparison. When evaluating multiple fish it is important to recognize that relative differences in hepatocyte size can only be diagnosed accurately in studies where sampling of the liver was conducted in a rigorously standardized and consistent manner.
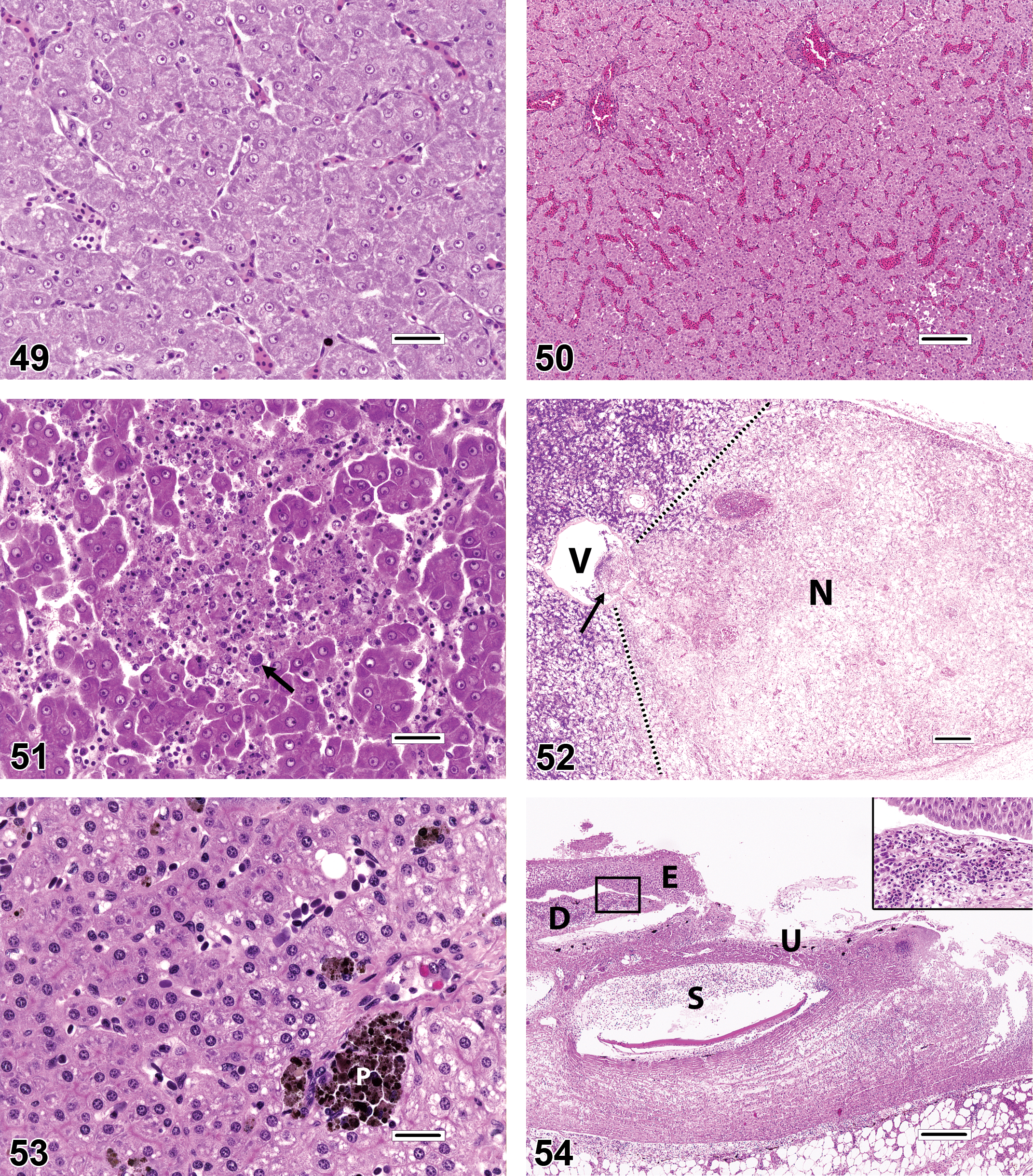
Other commonly reported liver diagnoses that frequently appear questionable when viewed in published images include congestion, dilated sinusoids, hepatocellular degeneration, and hepatocellular necrosis. In many instances, it appears that these are actually artifacts of tissue collection, preservation, or processing. Assessments of liver congestion and dilated sinusoids (Figure 50) are heavily contingent on the degree to which the fish was exsanguinated at sacrifice, and the amount of care taken not to manually squeeze the liver sample at necropsy. Livers that are partially autolyzed or focally traumatized during collection may result in misdiagnoses of hepatocellular degeneration and/or hepatocellular necrosis. Actual hepatocellular necrosis can be of the individual cell (apoptotic), focal (lytic or coagulative; Figure 51), or extensive (Figure 52) types, depending upon the cause. Note that except for the individual cell type, it is unusual for necrosis to be present without some degree of spatially associated inflammation. It is not uncommon to observe occasional spherical eosinophilic or amphophilic cytoplasmic inclusions in the livers of presumed healthy fish (Figure 47). These inclusions may represent a variety of different cellular and extracellular substances, including protein droplets, lipofuscin, ceroid, phagocytized erythrocytes, iron pigments, and/or apoptotic hepatocytes. When present in low numbers, such inclusions do not necessarily indicate a pathological degree of liver degeneration, although it is thought that an abundance of lipofuscin/ceroid material could signal a dietary imbalance in antioxidants such as vitamin E and selenium (Moccia et al. 1984).
PMAs (Figure 53) are constitutively present in organs such as the kidney, spleen, and liver. The general abundance, anatomic distribution, and coloration of these structures are often species dependent. PMAs are comprised chiefly of histiocytic macrophages that contain hemosiderin, melanin, lipofuscin, and/or ceroid pigments, and some reported functions of PMA include erythrophagocytosis, iron storage, and antigen presentation. Although it has been demonstrated that comparative changes in PMA size, number, and pigment content may be influenced nonspecifically by toxic exposures or environmental degradation (Fournie et al. 2001), some authors have erroneously reported the mere presence of PMA as an indication of environmental toxicosis. Additionally, because PMA mass tends to increase over time (Agius and Roberts 1981; Blazer et al. 1987; Brown and George 1985; Schwindt et al. 2006; Wolke 1992), the relative ages of individual fish must be considered as a confounding factor for any quantitative analysis of PMA.
Other Tissue Types
Tissues that are less frequently examined than the gills, kidney, and liver are also susceptible to errant diagnoses. For example, excessive handling of skin specimens can cause the epidermal layer to be inadvertently abraded off of the external surfaces of scales which can mimic dermal erosions or ulcers. It is important to recognize that true dermal ulcers (Figure 54) are often accompanied by acute or chronic inflammation that is patently evident subjacent and/or adjacent to the lesions.
There have been instances in which mineralized foci, cross-sections of nerves, or gonadal immaturity have been mistaken for gonadal intersex (Figures 55–58; Wolf 2011). Furthermore, there is evidence that false diagnoses of gonadal intersex may also be caused by inadvertent cross-contamination of samples during tissue collection or processing. Cross-contamination was demonstrated experimentally during a study in which various salmonid fishes (salmon, trout, and whitefish) were collected from various U.S. National Parks in the western United States as part of a survey for potential reproductive endocrine disruption effects (Schreck and Kent 2012). Results of this and a previous survey by Schwindt et al. (2009) had revealed a relatively low prevalence of testicular oocytes in male fish obtained from certain geographic locations, a finding that was supported by similar observations in historically archived specimens. However, in a subsequent survey, Schreck and Kent (2012) additionally detected a remarkably high prevalence (often >50%) of female fish that had testicular tissue situated deep within their ovaries. To investigate the authenticity of the latter finding, an experiment was designed in which plastic tissue cassettes containing ovaries from newly collected fish were placed in the same fixative container with testis specimens, and the resulting histologic preparations were compared to those of contralateral (i.e., left or right) ovaries that were not exposed to testicular tissue at any time (Schreck and Kent 2012). All instruments were wiped clean between animals to ensure that cross-contamination did not occur during the tissue-trimming phase. Ultimately, the results indicated that the presence of errant testicular tissue was limited to histologic sections in which the ovaries had been exposed to male gonads within the fixative container, thereby implicating cross-contamination as the cause of the false intersex lesions (Figure 57). It should be noted, however, that genuine ovarian spermatogenesis has been observed rarely, either as an apparently spontaneous or chemically induced finding (Figure 58).
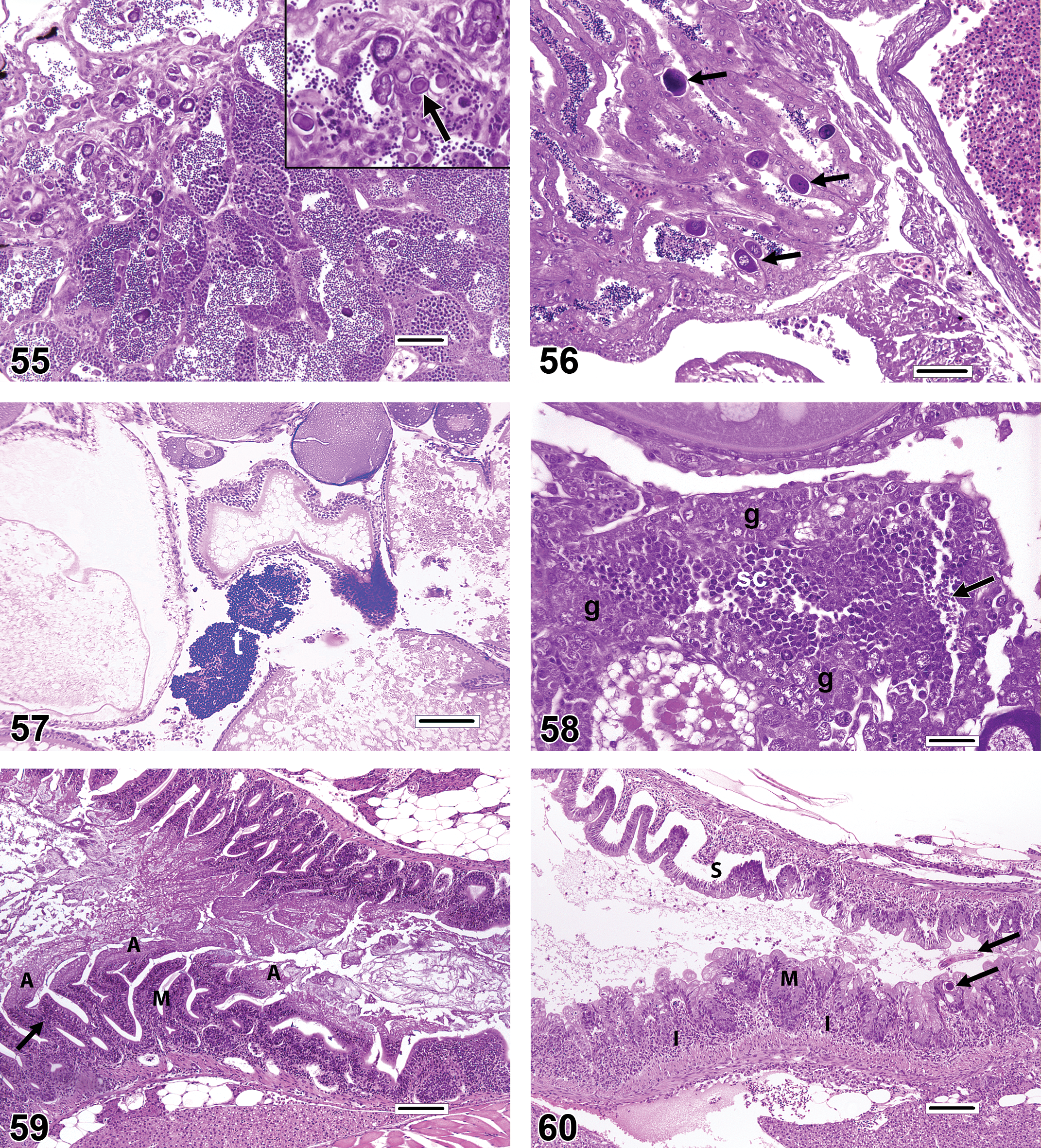
The intestinal epithelium is prone to autolysis if sampling is not performed appropriately and with rapid fixation (Figure 59); such autolytic changes can be mistaken for pathologic findings such as mucosal fold necrosis and/or atrophy (Figure 60). Additionally, while it is true that persistent toxic damage to the gut mucosa or chronic inflammation can produce morphologic changes in intestinal folds (villi) such as atrophy (shortening, blunting), enterocyte hyperplasia, and fusion of adjacent folds, the illusory appearance of these lesions in transverse sections can also be caused by plane-of-section artifacts in which individual folds are cut obliquely or tangentially, or where mucosal folds are superimposed (Figure 61). Because the size and complexity of folds tends to vary progressively along the length of the intestine, caution should be exercised when attempting to analyze specimens that were not collected in a uniform fashion (e.g., “transverse sections obtained precisely 1 cm distal to the pyloric ceca”). McKnight cells are individual apoptotic cells found within and among epithelial cells of the intestinal mucosa (Figure 62). Low numbers are often present in healthy fish, but they may increase dramatically as a sequel to enteritis, as in the pyloric ceca of trout clinically affected with infectious pancreatic necrosis virus (McKnight and Roberts 1976). It is possible to mistake these defunct cells for intracellular intestinal parasites such as coccidia.
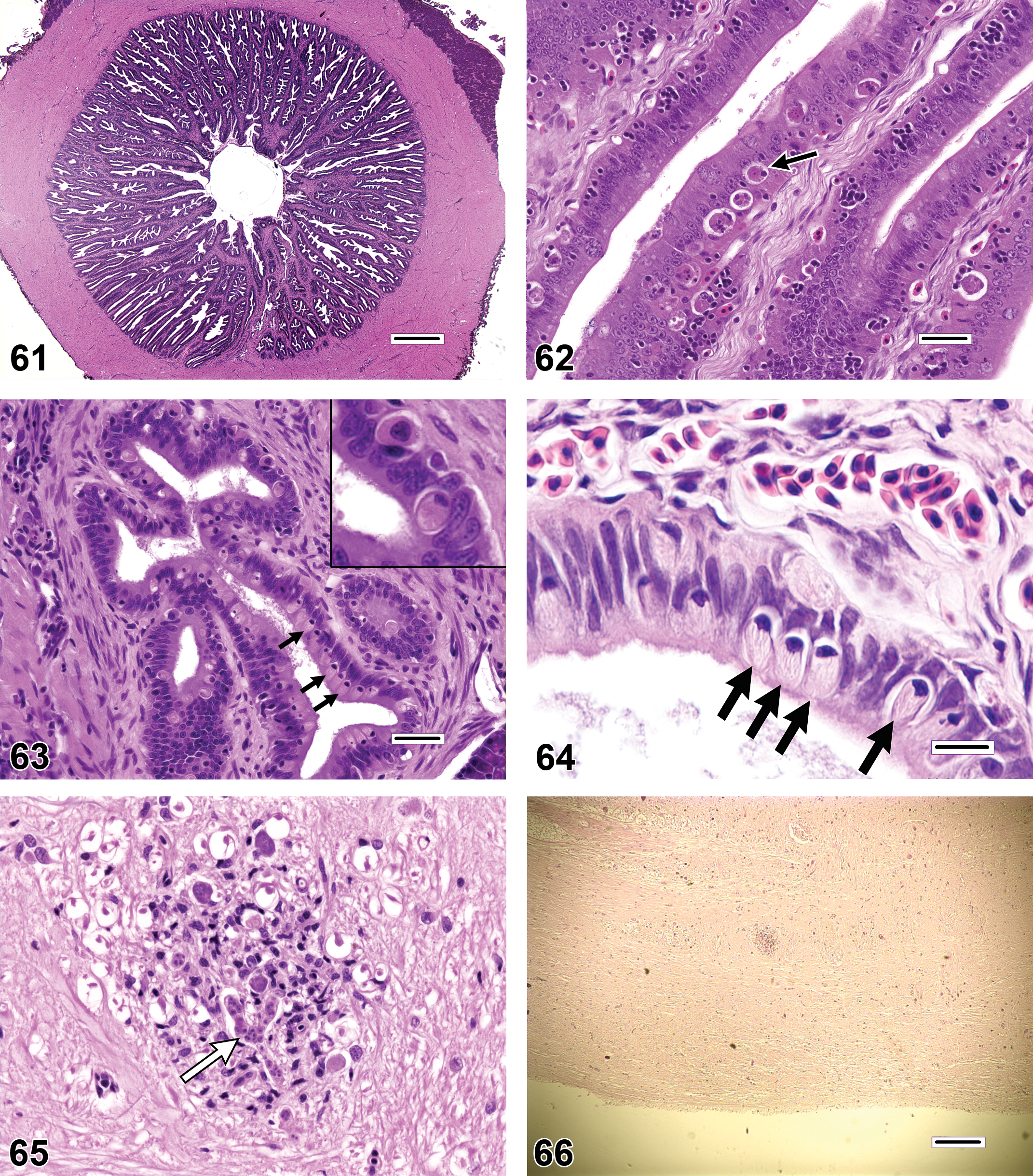
Rodlet cells (Figures 63–64) are found to varying degrees in a variety of anatomic locations in many species of teleost fishes. Once speculated to be parasites, they are now considered by most authorities to be of endogenous origin, but despite years of study the precise function of these cells remains unknown (Manera and Dezfuli 2004). Reportedly, rodlet cells have been observed at sites of tissue damage or parasitization; however, numerous rodlet cells can also be found in many overtly healthy tissues. Because consistent relationships between rodlet cell numbers and disease have not yet been established, care should be taken when ascribing pathological significance to the presence or abundance of these cells. Rodlet cells have also been mistaken for protozoan pathogens such as cryptosporidia (Saraiva et al. 2009).
Examples of Findings That May Be Underdiagnosed
The fish disease literature contains many photographic illustrations of histopathologic findings that appear to have been misdiagnosed; however, it is clearly more difficult to determine the degree to which certain morphologic changes may be largely underappreciated. Table 2 contains examples of findings that the authors tend to observe periodically in histologic sections but seldom encounter in published reports. For some of the items on this list, such as brain and ocular lesions (Figure 65), it is presumed that the lack of reported findings is due to the fact that these particular tissues are only collected intermittently for histopathologic evaluation. Additionally, the assessment of neurologic tissues in fish can be daunting, because the normal fish brain can appear hypercellular, and gray and white matter tracts may not be well delineated.
In the case of artifacts and background disease (e.g., incidental infections), it is likely that authors are hesitant to report such occurrences routinely; this is understandable, because the inclusion of ancillary findings tends to dilute the primary message that the authors would prefer to convey. However, it must also be acknowledged that the presence of some background diseases may influence diagnostic interpretations and affect study results, and to that extent it is both ethical and scientifically useful to describe the occurrence of potentially confounding background findings in published reports.
It is possible that other types of histopathologic changes are underreported because they have insufficient precedence in the literature. That is, investigators tend to search for types of lesions that have been described commonly in the past, whereas less frequently reported changes are not necessarily at the forefront of their awareness. Examples of diagnoses in this category are gill lamellar adhesions (vs. lamellar fusion), decreased hepatocellular vacuolation (often seen in sick, stressed, or malnourished fish), intestinal inflammation (focal or diffuse expansion of the submucosa by leukocytic infiltrates), renal hematopoietic tissue hyperplasia (which can be a response to profound localized or chronic systemic infection), and renal mineralization (a common finding that may be diet related in captive fish).
General Recommendations for Generating and Publishing Fish Histopathology Data
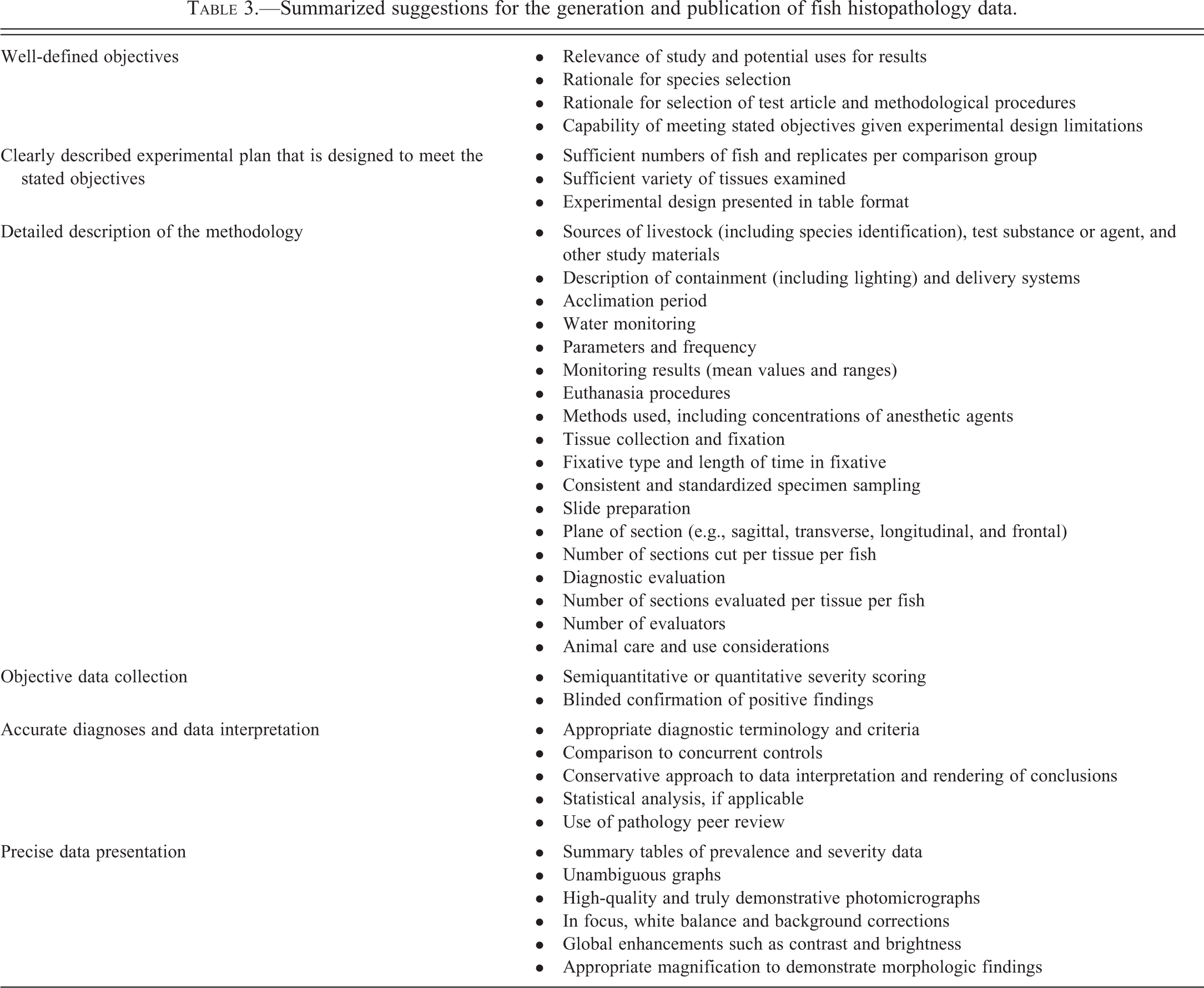
Few would argue that diagnostic accuracy should be a top priority for all scientific presentations and publications of fish histopathology data. However, the validity of a study is measured by more than diagnostic accuracy alone. Only when there is adequate evidence that the entire investigation was conducted in a scientifically appropriate manner with conclusions that are justified, can the audience or reader have confidence in the results. Table 3 outlines some suggestions for the generation and publication of histopathologic data for fish studies. Although a few of the listed items pertain specifically to fish or aquatic animals, many of these recommendations have broader application for pathology reporting in general. Major elements listed on Table 3 include well-defined objectives; a clearly described experimental plan that is designed to meet the stated objectives; a detailed description of the methodology; impartial data collection; accurate data interpretation; and precise data presentation.
Summarized suggestions for the generation and publication of fish histopathology data.
Objectives
A clear statement of purpose is the cornerstone of a well-written scientific manuscript, as it not only provides a preview of the research to follow, it often presents a narrative version of the null hypothesis (assuming the research is hypothesis based). Thus, by the end of the article, the reader should be able to determine readily whether the stated objectives have been achieved and whether the null hypothesis has been accepted or rejected. An important aspect of the objectives statement that often receives insufficient consideration is its scope, and there seems to be a tendency in fish health articles to overstate the goals in the text (and sometimes the title). For example, a statement such as “The objective of this article is to establish that chemical X negatively affects wild fish populations” may not take into account the limitations of an experimental design in which a single fish species was exposed to designated concentrations of a particular formulation under a defined set of controlled laboratory conditions; such a study would be unlikely to provide definitive evidence that chemical X affects all fishes within various environments at the population level. Additional elements related to the objectives that should be included routinely in the Introduction or Discussion sections are the overall academic, economic, and/or societal relevance of the study, potential utilization of the study results, and justifications for selection of the test species, the test compound or agent, and the methodological procedures.
Experimental Design
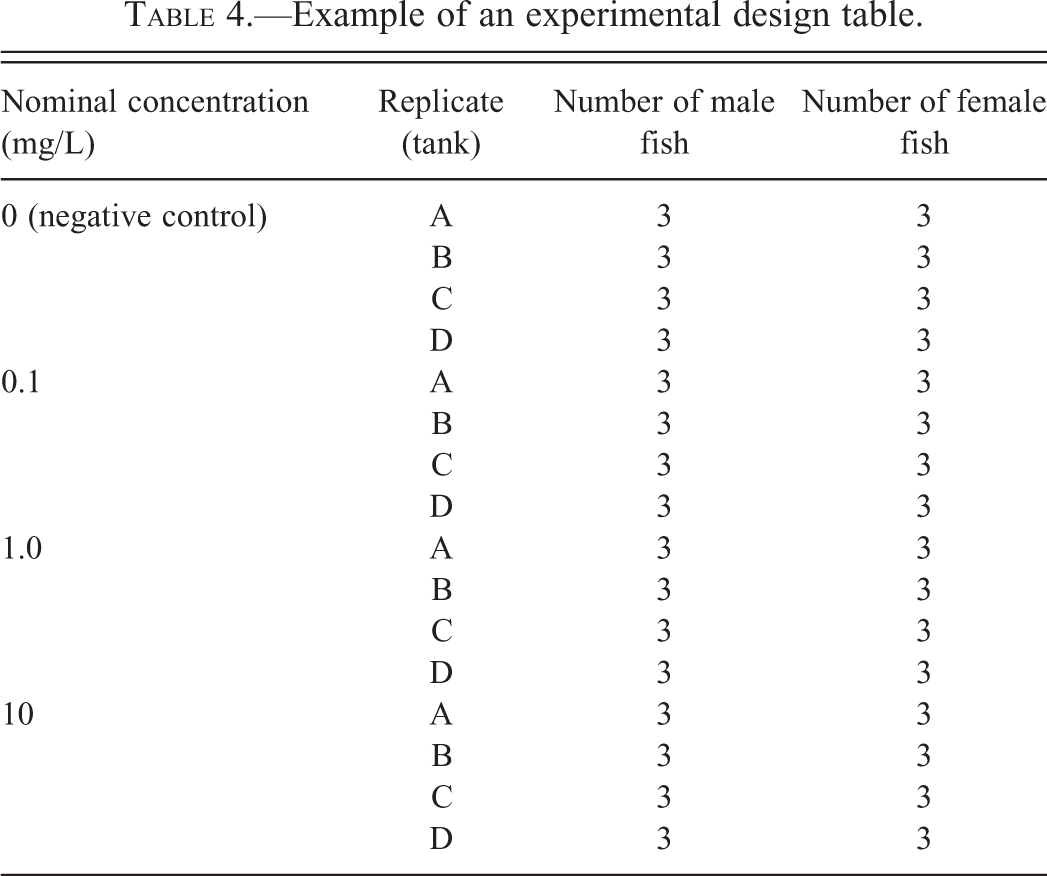
One of the most common issues that reviewers encounter when evaluating manuscripts from fish studies is difficulty in determining the number of animals that actually participated in various experimental phases. Frequently, numerical information presented in the text is confusing or ambiguous. This situation can be easily remedied by including in the methods section a well-designed table that outlines the experimental design. Table 4 is an example of a hypothetical design from a toxicological or pharmacological study performed in the laboratory. For a study that involves a survey of various waterways, the nominal or measured concentrations of the test substance can be replaced by collection site locations.
Example of an experimental design table.
There is no single correct formula for determining the number of animals per treatment group that should be used for studies that include fish histopathology. Consistent with welfare initiatives to “reduce, refine, and replace” animals used in biomedical research, the number of fish employed in any study should be limited as much as possible. Keeping that in mind, it is imperative that sufficient numbers of fish be used to answer the given research question; otherwise, all sacrificed animals will have been expended to no purpose. This optimum number of fish per group will vary according to the anticipated frequency of the expected histopathologic effect/effects in the treated population. Consequently, statistical power analysis, combined with a small pilot project to establish the appropriate dose range, may be of benefit in the long run. It is also important to recognize that unless it can be demonstrated that responses were not significantly different among replicates (tanks), then the tank, rather than the individual fish, should be considered the experimental unit for statistical comparisons.
Methods
Methodology should be described in enough detail to allow other investigators to reproduce the work if so desired. Common components that merit description include sources of fish livestock and key study materials; containment systems or collection sites; acclimation periods, water monitoring parameters and procedures; procedures used for euthanasia, tissue collection, and fixation; histologic slide preparation (including orientation of tissues during embedding, section thicknesses, and commercial sources for stains); antigen retrieval techniques, sources of primary and secondary antibodies, and specificity testing for immunohistochemical studies; diagnostic evaluation; and animal care and use considerations. Whenever possible, reports should also include descriptions of chemical analyses, biochemical testing, or other assays that were used to confirm levels of toxicant exposure or the patency of infection. Evidence should be presented to indicate that adequate sampling was performed, that sampling was consistent among animals, and that precautions were undertaken to minimize bias at every level (e.g., tank positioning, fish selection, and specimen evaluation). For example, the terminal sacrifice of experimental fish on a tank-by-tank basis, as opposed to the randomized sacrifice of individual fish, can be a potential source of bias because the groups may spend differing amounts of time in holding containers or on the bench awaiting necropsy dissection; consequently, artifactual changes such as autolysis may be mistaken for group-related findings. Regarding sampling bias, it is important to recognize that the term “random” suggests that mathematical tools (e.g., random number table or software generator) were used to facilitate the selection process; alternatively, when selection is performed without the use of mathematical tools, terms such as “indiscriminate,” “arbitrary,” or “impartial” are more appropriate.
Data Collection
As a discipline, histopathology has evolved beyond the point where it is acceptable to report experimental results merely as general observations (exceptions include non-hypothesis-based research, such as the morphologic characterization of a newly discovered pathogen). Rarely are biological responses entirely consistent throughout a randomly selected group of identically challenged animals. Therefore, for most studies, the prevalence and severity of histopathological findings should be recorded on an individual fish basis as opposed to group-wise, and this includes fish in negative control or reference site groups. When control or reference site fish are not scored because they are presumed to be normal, this creates a bias toward the generation of treatment or exposure effects. The use of semiquantitative grading systems (e.g., grade 1 = minimal, grade 2 = mild, grade 3 = moderate, grade 4 = marked, and grade 5 = severe) or quantitative morphologic measurements to assess the relative or absolute severity of nonneoplastic findings is strongly encouraged. As imperfect as these systems may be, they often greatly enhance the value of the histopathology data; this is especially true when lesion prevalence is comparable among the various comparison groups and exposure-related findings are only represented by differences in lesion severity. In addition, the use of a grading system allows the use of nonparametric statistical methodologies to compare treatment groups.
To improve the likelihood that subtle findings will be detected, it is advisable for the pathologist to be aware of the group identity of individual fish (i.e., remain “unblinded”) during the initial evaluation of the histologic sections (Crissman et al. 2004). Alternatively, if the study is to be read in a blinded manner, a second unblinded “reference” control group can be added to the experimental design to serve as a baseline for comparison; however, that reference control group should not be used for any statistical comparisons. Most importantly, once positive findings have been identified, it is highly recommended that the relevant slides be masked, coded, and reevaluated in a blinded fashion to confirm that significant findings are not false positive results. The value of incorporating this additional step for ensuring the accuracy of the study results cannot be overemphasized.
Diagnosis and Data Interpretation
Over the years, attempts have been made to standardize diagnostic criteria and terminology for describing pathologic changes in a variety of mammalian species. One of the most recent examples involves the ongoing joint efforts of the Society of Toxicologic Pathology, the European Society of Toxicologic Pathology, and several other groups to develop an authoritative systems-based guide: the International Harmonization of Nomenclature and Diagnostic Criteria for Lesions in Rats and Mice (INHAND; Society of Toxicologic Pathology 2013). Unfortunately, analogous guidance for fish histopathology has not traditionally been available. Although progress has been made regarding the standardization of diagnostic criteria and terminology for limited topics, such as proliferative thyroid lesions in fishes (Fournie et al. 2005), nomenclature encountered in the journal literature tends to be a mixed bag in terms of accuracy and consistency. Overall, two of the best resources to date for the fish pathologist are Systemic Pathology of Fish: A Text and Atlas of Normal Tissues in Teleosts and their Responses in Disease (Ferguson 2006), and Fish Pathology (Roberts 2012). Otherwise, when in doubt, a good general rule of thumb is to employ terminology that is more descriptive than interpretive. For example, the term “atrophy” implies that a normally sized tissue became smaller as part of a physiological or disease-induced process. While this is certainly true in some instances, in others it may also be possible that the tissue had never attained full size, in which case a more appropriate diagnosis would be “hypoplasia.” Consequently, the most straightforward approach would be to use less elegant terminology such as “decreased relative size,” if the pathogenesis of the finding cannot be determined with reasonable certainty.
However, the availability of appropriate diagnostic criteria and terminology does not guarantee that this information will be applied correctly by the pathologist. A paradigm that has become the standard for confirming the diagnostic accuracy of findings in mammalian toxicological pathology is pathology peer review. Pathology peer review entails the examination of a subset of slides from the study by a second (reviewing) pathologist. The slide subset often includes target tissues (i.e., those in which treatment-related findings have been identified), plus a predetermined small percentage of randomly selected control and treated fish for which all tissues are examined. Ideally, the reviewing pathologist has a greater level of study-relevant expertise than the original (study) pathologist. Following the second slide evaluation, the reviewing and study pathologists typically meet to examine slides together and resolve their diagnostic differences by mutual agreement. In special instances where biologically or toxicologically meaningful diagnostic differences persist, an ad hoc panel of experts called a Pathology Working Group (PWG) can be assembled to settle such differences. Although underutilized relative to mammalian research, peer review/PWG procedures have been employed successfully for some fish studies (Boorman et al. 1997; Wolf et al. 2014).
Statistical analysis of histopathology data is not always necessary (e.g., clear-cut cases in which the outcome is visually obvious by examination of tabulated data), but where applicable, lesion prevalence data may be evaluated using a Fisher’s Exact Test. Optimally, statistical treatments should simultaneously consider prevalence and severity scores of histopathologic findings; however, statistical tools for such combined analyses have not been widely available. A novel method for analyzing pathology data in this manner has been developed and is currently undergoing validation (Green et al. 2014). Whether statistical analysis is employed or not, caution should be exercised when grouping different types of histopathologic findings. For example, in an effort to facilitate data analysis, the pathologist may be tempted to consolidate all kidney findings into a single representative kidney lesion score. However, it is important to recognize that findings such as renal tubular necrosis and granulomatous renal inflammation often have dissimilar causes and consequences; thus, combining unrelated findings can potentially distort the interpretation of results by masking or exaggerating treatment effects. For similar reasons, the grouping of positive and negative classes of findings (e.g., increased and decreased liver vacuolation) into a single score is not advisable. Conversely, care should also be taken to avoid reporting types of diagnostic findings that are either redundant or entirely superfluous to the study outcome.
Data Presentation
It can be difficult to gauge the validity of an otherwise well-conducted scientific study if the data are not presented in a thorough, coherent, and unambiguous fashion. Table 5 provides an example of a summarized set of hypothetical histopathology data. Despite the complexity of the experimental design in this case, the relative prevalence and severity of findings among the various treatment groups and replicates can be readily appreciated. For example, from Table 5, it is evident that findings related to treatment with the test substance occurred primarily in the 1.0 and 10 mg/L dose groups, and included dose-related increases in the prevalence and severity of gill LEH, and a dose-related decrease in the severity of hepatocellular vacuolation in the liver. Not all tables have to be as complex as Table 5, which itself could be simplified by combining data from the 4 replicates. Again, it is important to note that although the data in this table are summarized, findings from individual fish are fully represented. Histopathology data can also be portrayed in graphical form, albeit somewhat less precisely and often less effectively. If data are presented graphically, it can be misleading to display the results simply as relative frequencies (i.e., percentages of animals affected), unless the number of fish per comparison group are also indicated in the chart.
Example of an experimental results table with hypothetical data.
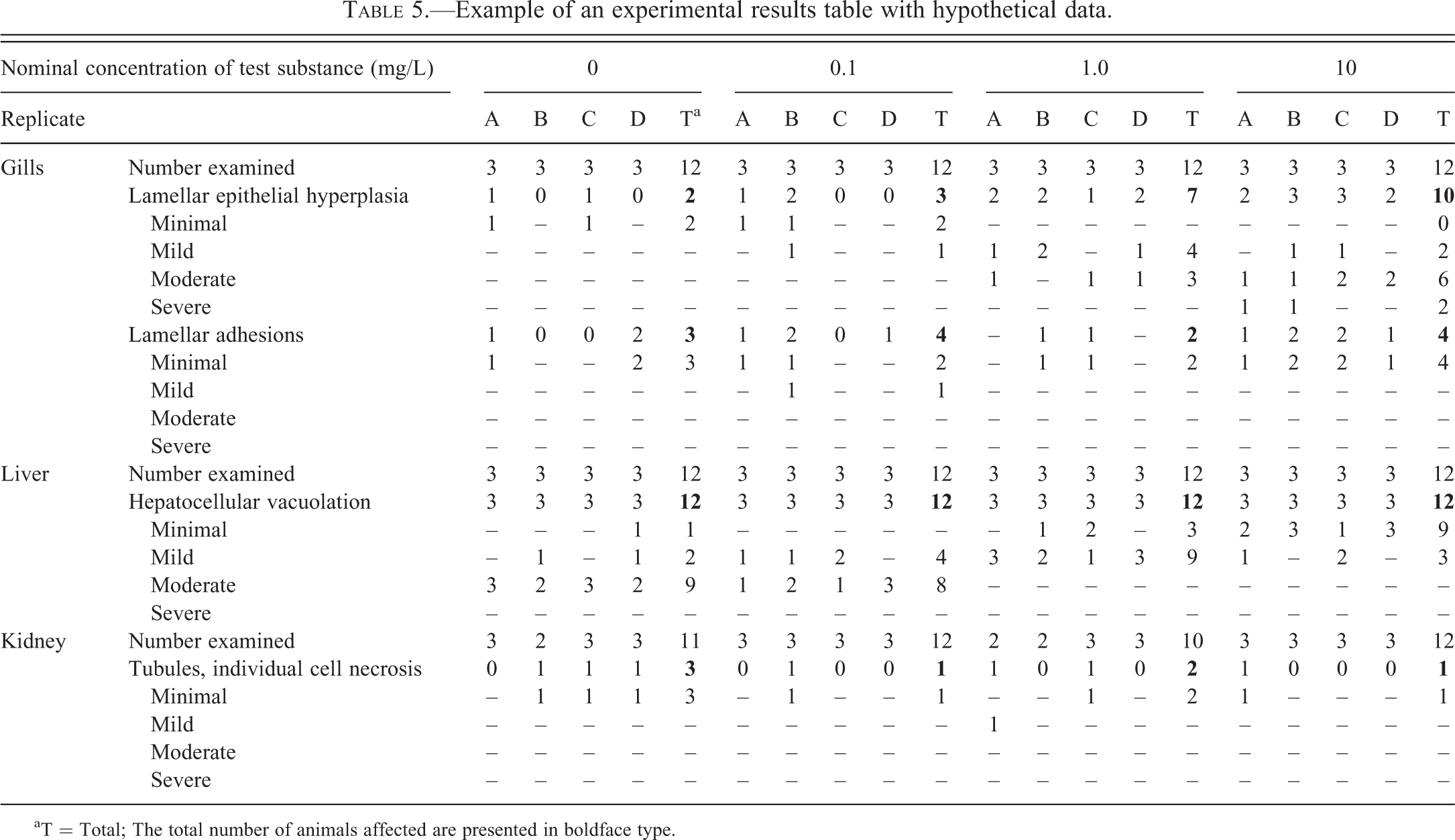
aT = Total; The total number of animals affected are presented in boldface type.
If positive histopathologic results are to be published, the manuscript should typically include representative photomicrographs of significant findings, because it is difficult to convey the subtleties of many morphologic changes through text descriptions alone. Admittedly, the overall legitimacy of microscopic diagnoses and interpretations cannot be confirmed merely by examining selected figure images; confirmation would require a review of the actual histologic sections. However, the inclusion of high-quality images can help knowledgeable readers and reviewers ascertain whether appropriate diagnostic criteria and terminology were applied, and if the quality of the histologic sections was sufficient for diagnostic purposes. The process of creating effective photomicrographic figures can be challenging. Not only do figures need to demonstrate features of abnormal versus normal tissues, they must illustrate the anatomic site, spatial extent, and distribution of lesions at magnifications that allow adequate detail to be observed. In order to accomplish these disparate objectives, it is frequently necessary to include multiple images of the same finding at different magnifications, or images with high-magnification insets. It should be acknowledged that most current camera systems are incapable of producing publication-ready images without some degree of post-acquisition processing. Accordingly, the creation of high-quality digital photomicrographs requires adjustments for Köhler (1894) illumination to optimize lighting of the histologic specimen, and corrections such as white balance (to correct for the color temperature of the light source), background subtraction (to compensate for uneven lighting across the image field), and global enhancement of contrast and color (Figure 66). However, care must be taken to ensure that all image processing is performed in an ethical manner; for further background and recommendations on this subject, readers are encouraged to consult Cromey (2010).
Conclusions and Final Recommendations
Despite a never-ending stream of technical advances in biological and ecotoxicological research, histopathology continues to be a valuable tool for investigating the morphologic features and extent of both naturally occurring and experimentally induced disease. Not only does it serve as a key link between apical (i.e., population relevant) and subcellular (e.g., molecular) endpoints, histopathology remains one of the most reliable, sensitive, and comprehensive assays for identifying and characterizing a vast array of physical disorders. In an experimental capacity, histopathologic results are used frequently to determine potential treatment effects, offer insights into mechanisms of action, and provide phenotypic mapping of genetically altered animals. Additionally, if sufficient sampling is performed, the presence of confounding subclinical disease/diseases may also be detected. However, histopathology is the proverbial double-edged sword. Analogous to erroneous conclusions in other scientific disciplines, histopathologic misdiagnoses and misinterpretations can have negative consequences that reverberate far beyond the scope of a single study. For example, based on incorrect histopathologic evidence, an article in a peer-reviewed journal may report that compound X causes renal tubular necrosis in a certain species of fish. That inaccurate report might prompt additional investigations into those and other effects of compound X, plus its metabolites and related chemicals, in that particular species and in other fishes, which would in turn require the allocation of further efforts and funding. Results of such unintentionally misguided research might then be used by regulatory agencies to formulate public policy decisions that ultimately have inappropriate financial, health, and/or environmental impacts. Furthermore, once they appear in the literature, inaccurate results tend to persist, and their existence is not necessarily expunged by letters to the editor or subsequent reports of contradictory findings.
Fortunately, there are practical measures that can be instituted to safeguard against the publication of patently incorrect histopathologic findings and data interpretations. First, investigators should reevaluate all experimentally significant findings in a blinded fashion to ensure that none represent false-positive results (i.e., type I errors). This is a rapid and effective initial approach for minimizing the effects of subconscious bias. Second, it is often advisable to have a second pathologist review at least a subset of the histologic slides to confirm the validity of key diagnoses (pathology peer review), and to verify that significant findings have not been missed (i.e., false negative, type II errors). This peer review process can be formal (e.g., paid consultation with documented results) or informal (e.g., second opinion performed as a professional courtesy), although for the sake of objectivity it may be preferable to select the reviewing pathologist from outside the original institution. The reviewing pathologist could also be asked to weigh in on interpretation of the histopathology data. A third mechanism would be to preview preliminary results as a poster or oral presentation to obtain feedback from knowledgeable professionals prior to manuscript submission. Fourth, journal editors should strive to solicit reviewers who have sufficient expertise to assess histopathologic results in manuscripts. This can be challenging, as the pool of qualified fish pathologists is to some extent limited, and there are no universally accessible registries for identifying such individuals. Although always a difficult decision, editors or associate editors of journals may opt to preemptively reject manuscripts in which it is clear that the histopathologic descriptions and photomicrographs do not meet minimum quality standards, rather than burden overtaxed reviewers with articles that have little chance of ultimate acceptance. Finally, if all else fails, authors should subscribe to the philosophy of “when in doubt, leave it out.” Thus, authors should avoid submitting manuscripts that contain questionable findings or conclusions, with the expectation that journal reviewers will automatically recognize and rectify any diagnostic or interpretive issues.
It is hoped that this article will prove to be a helpful guide for authors, reviewers, and readers. Ostensibly, Table 3 could serve as a useful checklist for determining a manuscript’s suitability for publication and for assessing the quality of articles that are already in print. The fundamental goal of this effort is to elevate the science and practice of fish histopathology, which has become an increasingly important discipline in fields that include basic biomedical research, aquaculture, environmental resource management, and ecotoxicology.
Footnotes
Acknowledgment
The authors would like to express their sincere appreciation to Beth Mahler for her assistance with the photomicrographic figures.
Authors’ Note
The views expressed herein are those of the authors and do not necessarily reflect the views or policies of the U.S. Geological Survey or U.S. Environmental Protection Agency. Eric D. Lombardini is a Lieutenant Colonel in the U.S. Army. The opinions or assertions herein are those of the authors and do not necessarily reflect the view of the Department of the Army or The Department of Defense. Use of trade names is for identification purposes only and does not imply endorsement by the U.S. government.
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Abbreviations
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.