
Abstract
During pregnancy, the maternal immune system is challenged by the semiallogeneic fetus, which must be tolerated without compromising fetal or maternal health. This review updates the systemic and local immune changes taking place during human pregnancy, including some examples in rodents. Systemic changes are induced by contact of maternal blood with placental factors and include enhanced innate immunity with increased activation of granulocytes and nonclassical monocytes. Although a bias toward T helper (Th2) and regulatory T cell (Treg) immunity has been associated with healthy pregnancy, the relationship between different circulating Th cell subsets is not straightforward. Instead, these adaptations appear most evidently at the fetal–maternal interface, where for instance Tregs are enriched and promote fetal tolerance. Also innate immune cells, that is, natural killer cells and macrophages, are enriched, constituting the majority of decidual leukocytes. These cells not only contribute to immune regulation but also aid in establishing the placenta by promoting trophoblast recruitment and angiogenesis. Thus, proper interaction between leukocytes and placental trophoblasts is necessary for normal placentation and immune adaptation. Consequently, spontaneous maladaptation or interference of the immune system with toxic substances may be important contributing factors for the development of pregnancy complications such as preeclampsia, preterm labor, and recurrent miscarriages.
Introduction
The maternal immune system during pregnancy is changed in order to tolerate the semiallogeneic fetus. This is especially important in species with a hemochorial placenta (for instance, humans, monkeys, and rodents), since this is the most invasive type of placenta, in which there is an intimate contact between the maternal immune system and fetal tissue (Meeusen, Bischof, and Lee 2001). The immune adaptations include changes in local immune responses, that is, in the uterine mucosa (decidua; Trundley and Moffett 2004), and systemic changes (Chaouat 2003). Immediately after implantation, the endometrium is infiltrated by fetal trophoblast cells and will develop into the decidua and ensure anchorage of the placenta and therefore proper fetal nutrition. However, this trophoblast invasion needs proper regulation in order to protect the integrity of the uterus. Therefore, local immune cells, such as uterine natural killer (NK) cells, macrophages, γ/δ, and regulatory T cells (Tregs), are present in the endometrium already before implantation and increase in numbers and adapt immediately following implantation (Trundley and Moffett 2004; Figure 1). These immune cells are important regulators of the balance between tolerance of fetal trophoblast cells and limitation of their invasion (Lash, Robson, and Bulmer 2010; Nagamatsu and Schust 2010). After establishment of the placental circulation, that is, between 8 to12 weeks of gestation, the peripheral blood also comes into close contact with the fetal cells in the intervillous space, that is, the villous trophoblasts, which may affect the systemic immune response (Figure 1). The most evident changes seen in the systemic circulation are a shift from a T helper (Th1) toward a Th2 immune response and increased activation of innate immune cells (described in more detail in later sections). The present review will focus on cellular and molecular adaptations that take place in the circulation (systemically) and in the decidua and the chorioallantoic placenta (locally) during pregnancy. The recent contributions in the field will be highlighted, providing a background for understanding the effects of maladaptation as well as interference of the immune response in pregnancy. In general, the review refers to studies in humans although information referring to studies in rodents is included as well.

The fetal–maternal interface. Fetally derived trophoblast cells are in contact with maternal immune cells at two sites: (1) villous trophoblasts (VTs) are surrounded by maternal immune cells in the intervillous space and (2) invading extravillous trophoblasts (EVTs) interact with decidual leukocytes. The VTs produce various cytokines and chemokines, and generate microparticles that may affect maternal blood leukocytes. The major decidual leukocyte populations are natural killer (NK) cells, macrophages (MΦ), and T cells (cytotoxic [Tc], helper [Th], and γδ T cells). A small population of dendritic cells (DCs) is also present. The main functions of NK cells, macrophages, and regulatory T cells (Tregs) are summarized schematically. NK cells support spiral artery remodeling and EVT invasion via the production of angiogenic and chemotactic factors. The interaction between NK cell receptors and ligands on EVTs are important for adequate decidual NK cell function. The macrophages have immune regulatory properties that may support tissue homeostasis, for example, by clearing infections and limiting T cell activation. They also support spiral artery remodeling via angiogenic factors and the clearance of apoptotic debris. Tregs are important immune suppressors and might contribute to fetal tolerance by inhibiting T cell activation and interacting with antigen presenting cells. CTLA-4, cytotoxic T lymphocyte antigen 4; Foxp3, forkhead box p3; GM-CSF, granulocyte-macrophage colony-stimulating factor; IDO, indoleamine 2,3-dioxygenase; KIR, killer immunoglobulin-like receptor; M-CSF, macrophage colony-stimulating factor; MMP9, matrix metalloproteinase 9; PlGF, placental growth factor; VEGF, vascular endothelial growth factor.
Changes in the Systemic Immune Response during Pregnancy
After the establishment of the placental circulation, maternal blood is in close contact with the fetal villous syncytiotrophoblasts (Burton, Jauniaux, and Charnock-Jones 2010). Such contact with fetal cells may influence the maternal immune response. It has indeed been shown that the passage of maternal blood through the placenta activates inflammatory cells such as granulocytes and monocytes (Mellembakken et al. 2002). This may be due to contact of these immune cells with fetal trophoblast cells during placental passage. However, villous trophoblasts have been shown to produce and shed various factors, like cytokines (Naruse et al. 2010; Sacks et al. 2001) syncytiotrophoblast microfragments (Redman et al. 2012), fetal cells (Bianchi et al. 1996), and pregnancy hormones, for example, progesterone, estrogen, or human chorionic gonadotropin, into the maternal circulation (Figure 1). All these factors may affect the immune system (Dong et al. 2008; Han 1975; Piccinni et al. 1995; Southcombe et al. 2011). Recently, studies have indeed shown the presence of factors in the plasma of pregnant women which activate maternal immune cells such as monocytes in vitro (Faas et al. 2008; Faas, van Pampus, et al. 2010).
Pregnancy-induced Changes in the Systemic Innate Immune Response
It has long been established that innate immune responses change during pregnancy. This has most often been shown by higher numbers of circulating monocytes and granulocytes, resulting in increased numbers of total leukocytes during pregnancy (Kuhnert et al. 1998; Siegel and Gleicher 1981; Veenstra van Nieuwenhoven et al. 2002). However, there is substantial evidence that not only the numbers of these innate immune cells are increased, but they also show phenotypic and functional activation. Various studies have for instance shown upregulation of activation markers, such as CD11b or CD64, on monocytes and granulocytes (Davis, Kaufmann, and Moticka 1998; Luppi, Haluszczak, Trucco et al. 2002; Naccasha et al. 2001; Sacks et al. 1998). In accordance, it has recently been shown that changes take place in monocyte subsets during pregnancy. Increased numbers of nonclassical monocytes (CD14+CD16+ or CD14++CD16+) and decreased numbers of classical monocytes (CD14++CD16−) have been found in the third trimester of human pregnancy (Melgert et al. 2012). The nonclassical monocytes can be considered more proinflammatory than the classical subset, since they have been shown to produce increased amounts of proinflammatory cytokines, such as tumor necrosis factor (TNF) and interleukin (IL)-1β (Ancuta et al. 2009; Cros et al. 2010; Ziegler-Heitbrock et al. 2010). Their proinflammatory association was also illustrated by the fact that their numbers increased during inflammatory conditions, such as during sepsis (Fingerle et al. 1993). Interestingly, these phenotypic changes observed in monocytes during pregnancy appear to be similar to changes seen in septic patients (Fingerle et al. 1993; Sacks et al. 1998). Various other studies have shown that monocytes also show functional changes during pregnancy, for instance, cytokine as well as oxygen radical production change following activation in vitro (Luppi, Haluszczak, Betters et al. 2002; Sacks et al. 1998; Veenstra van Nieuwenhoven et al. 2003). Also, granulocyte function is changed during pregnancy: granulocytes show increased production of oxygen radicals (Sacks et al. 1998) and increased phagocytic activity (Barriga, Rodriquez, and Ortega 1994). The number of dendritic cells (DCs), which are antigen presenting cells, on the other hand, were found to be decreased during pregnancy (Cordeau et al. 2012; Darmochwal-Kolarz et al. 2003; Shin et al. 2009; Ueda et al. 2003). However, it has been shown that these cells show increased tolerogenic properties (Darmochwal-Kolarz et al. 2012): upon stimulation, interferon (IFN)-α production and costimulatory molecules such as CD54 and CD86 are decreased (Cordeau et al. 2012), while the expression of tolerance-associated molecules, for example, CD200 and CD200R, is increased during pregnancy (Darmochwal-Kolarz et al. 2012). Finally, the number of circulating NK cells is decreased during pregnancy (Kuhnert et al. 1998; Veenstra van Nieuwenhoven et al. 2002; Veenstra van Nieuwenhoven, Heineman, and Faas 2003; Watanabe et al. 1997). Moreover, their production of IFNγ is decreased in pregnant as compared with nonpregnant women (Veenstra van Nieuwenhoven et al. 2002) and it has been shown that, similar to the shift toward Th2 cells (described in the next section), there was also a shift toward NK2 cells (NK cells producing Type 2 cytokines) during pregnancy (Borzychowski et al. 2005). It is now generally accepted that the innate immune response is activated during pregnancy.
These changes in the innate immune response appear to be important for normal pregnancy, since pregnancy complications, like preeclampsia or intrauterine growth retardation, are associated with deviations from pregnancy-induced adaptations in the innate immune response. During preeclampsia, both neutrophils and monocytes are even further activated, phenotypically and functionally, as compared with normal pregnancy (Gervasi et al. 2001; Mellembakken et al. 2002; Redman, Sacks, and Sargent 1999; Sacks, Redman, and Sargent 2003; Sacks et al. 1998; Sakai et al. 2002; Veenstra van Nieuwenhoven et al. 2008). These data show that aberrant activation of the innate immune response during pregnancy results in pregnancy complications, suggesting that the adaptation of the innate immune response is tightly regulated.
Pregnancy-induced Changes in the Systemic Adaptive Immune Response
Not only the innate immune response is changed during pregnancy but also the adaptive immune response shows changes. The concept proposed by Wegmann et al. (1993) that a healthy pregnancy is accompanied by a decreased Th1/Th2 ratio has been confirmed by many others (Naccasha et al. 2001; Sacks et al. 1998; Saito et al. 1999; Veenstra van Nieuwenhoven et al. 2002). However, although it is generally recognized that a shift away from the Th1 response is important in certain stages of pregnancy, it has now also been shown that Type 1 cytokines, such as IFNγ or TNF, are not only detrimental to pregnancy but also play an important role in placental development. It has for instance been shown that they are needed in triggering pregnancy-induced spiral artery remodeling (Ashkar, Di Santo, and Croy 2000; Zhang et al. 2003). Moreover, the immunological paradox of pregnancy appears to be much more complicated than previously thought. Th17 cells and Treg cells have now also been implicated in the complex immune regulation during pregnancy (Ernerudh, Berg, and Mjosberg 2011; Saito et al. 2010). However, the exact role of circulating Treg cells during pregnancy remains unknown, since inconclusive results have been found. Several studies have demonstrated that Treg cells are essential for promoting immune tolerance, and initially circulating Treg cell numbers were reported to increase during pregnancy (Aluvihare, Kallikourdis, and Betz 2004; Sasaki et al. 2004; Somerset et al. 2004). However, more recent studies have shown that peripheral blood Treg cells are not altered or even decreased during pregnancy (Mjosberg et al. 2009; Tilburgs et al. 2008), indicating that they may predominately exert their tolerogenic functions locally.
Only a few studies have been performed on Th17 cells in pregnancy and also these results are inconclusive: some studies have shown that circulating Th17 cell numbers are not different in pregnant as compared with nonpregnant women (Nakashima et al. 2010; Toldi et al. 2011), while others have reported them to be increased during pregnancy (Liu et al. 2011). The contradicting results may be due to differences in methods or patient selection, which may have a great impact given the limited number of studies performed to date. Therefore, future studies should focus not only on the role of Th1 and Th2 cells but also on the role of Treg and Th17 cells during pregnancy. This is more important since findings of the numbers of Treg cells during preeclampsia are conflicting. Some authors show that in preeclampsia the numbers of circulating Treg cells are decreased compared with healthy pregnant women (Darmochwal-Kolarz et al. 2012; Prins et al. 2009) while others did not find different numbers of Treg cells between preeclamptic patients and normal pregnant women (Hu et al. 2008; Paeschke et al. 2005). Some of the inconsistencies regarding Treg cells may refer to how they were defined (Mjosberg et al. 2009). Although there is disagreement on the role of Treg cells in preeclampsia, there is agreement that preeclampsia is associated with a shift of the Th1/Th2 balance toward Th1 immune responses in severe preeclampsia (Boij et al. 2012; Borzychowski et al. 2005; Darmochwal-Kolarz et al. 1999, 2002; Makhseed et al. 1999). In mild preeclampsia, however, it was shown that the maternal immune response is shifted toward a Type 2 immune response (Veenstra van Nieuwenhoven et al. 2008). Therefore, as for innate immune responses, it also appears important for normal pregnancy to tightly regulate the Th1/Th2 balance.
Maternal Consequences of the Adapted Systemic Immune Response
Although the maternal immune response adapts to pregnancy, most pregnant women experience a healthy pregnancy and do not show an increased susceptibility to most infections. This suggests that the immunological changes do not dramatically affect the integrity of the mother. However, it has been shown that pregnant women are more sensitive to certain infections that depend on Th1-mediated responses, such as the risk of developing clinical disease after infection with poliovirus or hepatitis A virus or an increased infectivity of cytomegalovirus (Weinberg 1984), herpes (Fink et al. 1993), or malaria infection (Lagerberg 2008). Also, influenza virus infections, including the recent pandemic H1/N1 influenza, lead to a higher morbidity and mortality in pregnant women (Louie et al. 2010). Notably, pregnant women seem to respond properly to influenza vaccination (Fisher et al. 2012; Steinhoff et al. 2010), and vaccination against influenza, as well as to other infectious agents, is widely and strongly recommended in order to protect the mother, the fetus, and the neonate (Munoz and Ferrieri 2012). In addition to the effects of pregnancy on infection susceptibility, it has long been known that changes in the expression of autoimmune diseases are obvious during pregnancy: rheumatoid arthritis and multiple sclerosis, which are roughly dominated by a Th1/Th17-type response, often ameliorate during pregnancy followed by a “rebound” worsening postpartum (Confavreux et al. 1998; Ostensen, Aune, and Husby 1983). These effects of pregnancy on autoimmune diseases likely result from the shift of the immune response from a Th1-type response toward a Th2-type immune response.
In accordance with the increased sensitivity of pregnant women to certain infections, pregnant individuals are also much more sensitive to proinflammatory stimuli than nonpregnant individuals (Bakker et al. 1989; Faas et al. 1992, 1995; Theiss and Beller 1972; Visscher et al. 1993). It has for instance been shown that pregnant animals are extremely sensitive to lipopolysaccharide (LPS) or ATP, potent stimulators of innate immune responses: a very low dose of LPS or ATP infused into pregnant rats induced hypertension and/or proteinuria as well as a persistent inflammatory response, whereas infusion of similar amounts of these substances into nonpregnant animals did not induce these effects (Faas et al. 1995, 2000; Faas, van der Schaaf, et al. 2010). The exact mechanism of this increased sensitivity to proinflammatory stimuli during pregnancy remains unknown. However, it seems to be related to the changes in the innate immune response during pregnancy, since infusion of proinflammatory stimuli in the pregnant rat induced a much more intense and persistent activation of the inflammatory response (Faas et al. 2004, 1995, 2000; Faas, van der Schaaf, et al. 2010). Moreover, vascular tissue of pregnant rats appeared more sensitive to the products of activated inflammatory cells, such as oxygen free radicals (Faas et al. 1999). Adverse effects on pregnancy in experimental animals have also been described after infusion of other proinflammatory substances, also in low doses, such as poly I:C (Arad et al. 2005) and TNF (Alexander et al. 2002), as well as injection of Type 1 cytokines (Athanassakis et al. 1996). Interestingly, the adverse effects of proinflammatory changes during pregnancy are counteracted by anti-inflammatory responses, in particular involving IL-10. In IL-10(−/−) mice, LPS (Murphy et al. 2005), or CpG-induced toll-like receptor 9 stimulation (Thaxton, Romero, and Sharma 2009) caused adverse pregnancy outcomes in lower amounts than in wild-type mice, which could be rescued by administration of IL-10. Thus, not only hyperinflammatory responses but also failures of mounting an anti-inflammatory response may be key elements in the development of pregnancy complications.
Local Adaptation of the Immune System during Pregnancy
The decidua, the site where maternal cells come into direct contact with fetally derived trophoblast cells, is a specialized tissue with a unique composition of immune cells (Figure 1). The majority of the decidual leukocytes are innate immune cells, with NK cells and macrophages encompassing ∼90% of all leukocytes, whereas only ∼10% are T cells in early human pregnancy (King et al. 1991; Starkey, Sargent, and Redman 1988). A few DCs are also present while granulocytes and B cells are practically absent (Bulmer and Johnson 1984; Gardner and Moffett 2003). This is in contrast to the situation in blood, where the dominating populations are granulocytes and T cells.
Cells of the Innate Immune System
Decidual NK cells
NK cells are the dominating population in the first trimester decidua (∼70% of all leukocytes; Bulmer et al. 1991; King et al. 1991), highlighting their central role at the time when the placenta is being established. The common view is that decidual NK (dNK) cell numbers decrease after the first trimester to become nearly absent at term (Moffett-King 2002). However, more recent data indicate that they are present during all stages of pregnancy (Rieger et al. 2009; Sanchez-Rodriguez et al. 2011; Williams, Searle, et al. 2009), implying that they also contribute to the regulation of pregnancy at later stages. The origin of dNK cells or the reason for their dramatic increase is not well established. However, several mechanisms (that may operate in parallel) have been suggested including: (1) the recruitment of peripheral blood NK cells that differentiate locally into dNK cells (Carlino et al. 2008; Keskin et al. 2007; Male et al. 2010), (2) the maturation from endometrial NK cells in response to pregnancy-associated factors (such as IL-15; Manaster et al. 2008), and (3) the differentiation from hematopoietic precursors present in the decidua in response to decidual stromal factors (Vacca, Vitale, et al. 2011).
In peripheral blood, NK cells can be divided into a major subset of CD56dimCD16bright (∼90%) and a smaller population of CD56brightCD16−/dim cells (∼10%; Nagler et al. 1989). These populations differ in several aspects, including cytotoxic potential, cytokine production, and expression of cell surface markers (Cooper et al. 2001). The CD56brightCD16−/dim population is sometimes referred to as regulatory because of its reduced cytotoxic capacity and dominating potential to produce a range of cytokines influencing immune responses (Cooper, Fehniger, and Caligiuri 2001; Nagler et al. 1989). The vast majority of the dNK cells are CD56brightCD16− (King et al. 1991; Starkey, Sargent, and Redman 1988), although they seem to be a unique subset, different from both CD56dim and CD56bright blood NK cells, as judged by their gene expression profile (Koopman et al. 2003). dNK cells share the ability of blood CD56bright NK cells to produce cytokines (Engert et al. 2007; Lidstrom et al. 2003; Saito et al. 1993) and they express immunosuppressive molecules (e.g., CD9, galectin 1, and glycodelin A; Koopman et al. 2003). However, they also express NK cell activating receptors (e.g., 2B4, NKp30, NKp44, NKp46, NKG2D; Hanna et al. 2006; Kopcow et al. 2005) and cytolytic granules (King et al. 1991; Koopman et al. 2003) making them potentially cytotoxic. Nevertheless, dNK cells have significantly reduced cytotoxic ability (Kopcow et al. 2005), which favors their close association with invading extravillous trophoblasts (EVTs; Figure 1). The inefficient cytotoxic ability is believed to be due to the interaction of inhibitory receptors on dNK cells (e.g., ILT2, KIR2DL4, and CD94/NKG2A) and MHC class I molecules (HLA-C, -E, and -G) on trophoblasts (Manaster and Mandelboim 2010; Vacca, Moretta, et al. 2011; Figure 1).
Two major functions attributed to dNK cells are the ability to promote trophoblast invasion and angiogenesis (Figure 1). The recruitment of EVTs has been shown to be mediated via secretion of CXCL8 (IL-8) and CXCL10, for which EVTs express the receptors CXCR1 and CXCR3, and by hepatocyte growth factor (De Oliveira et al. 2010; Fraser et al. 2012; Hanna et al. 2006). dNK cells, found in close association with remodeling vessels (Hazan et al. 2010; Smith et al. 2009), may also contribute to spiral artery remodeling through production of angiogenic factors such as vascular endothelial growth factor (VEGF), placental growth factor (PlGF), and angiopoietins (Engert et al. 2007; Hanna et al. 2006; Kalkunte et al. 2009; Lash et al. 2006). The interactions between dNK cell receptors and HLA class I molecules on EVTs seem to play an important role in these processes. In particular, the ligation of activating dNK cell receptors by HLA-C molecules is favorable for the secretion of both chemotactic and angiogenic factors (Hanna et al. 2006). On the other hand, the presence of inhibitory receptors (e.g., KIRs) has been associated with an increased risk of complications, including preeclampsia (Hiby et al. 2004; Sanchez-Rodriguez et al. 2011), which is characterized by shallow trophoblast invasion and defective vascular remodeling (Figure 2). These data suggest that proper dNK cell activation is a prerequisite for adequate trophoblast invasion and spiral artery remodeling, and thus a normal placentation.
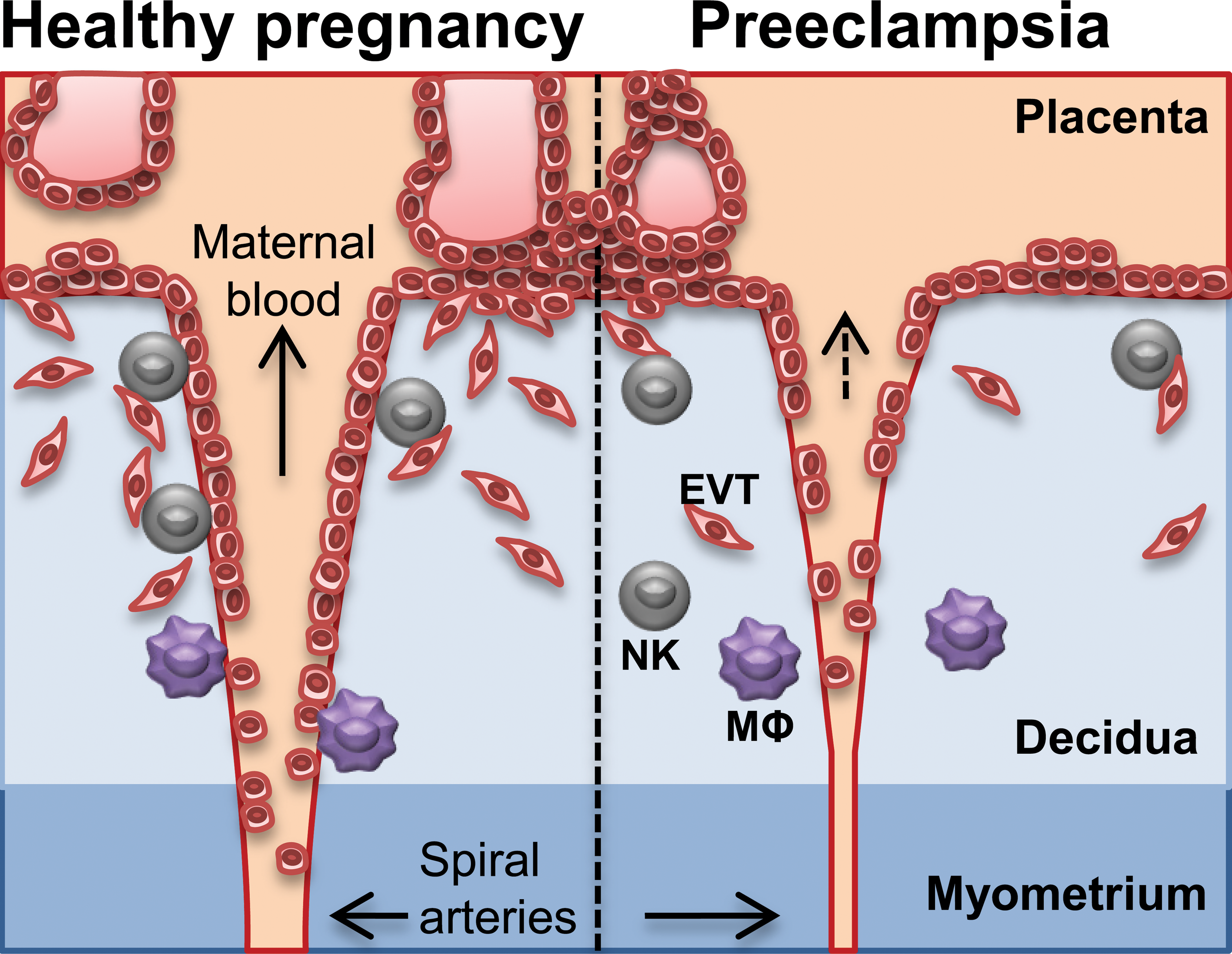
Spiral arteries and trophoblast invasion in healthy and preeclamptic pregnancy. Preeclampsia is characterized by shallow trophoblast invasion and defective vascular remodeling leading to reduced maternal blood flow, which in turn may compromise fetal growth. Failure of decidual natural killer (NK) cells and macrophages (MΦ) to recruit trophoblasts and support angiogenesis may contribute to the development of the disease. EVT: extravillous trophoblast.
Numerous studies have also addressed the numbers of dNK cells in complications, although the results are inconsistent, for instance, showing reduced, increased, or unchanged dNK cell numbers in preeclampsia and fetal growth restriction (Bachmayer et al. 2006; Eide et al. 2006; Rieger et al. 2009; Sanchez-Rodriguez et al. 2011; Wilczynski et al. 2003; Williams, Bulmer et al. 2009).
Decidual Macrophages
Macrophages account for ∼20% of decidual leukocytes and represent the most frequent antigen presenting cells throughout pregnancy (Bulmer and Johnson 1984; Starkey, Sargent, and Redman 1988; Williams, Searle et al. 2009). Tissue macrophages are commonly divided into proinflammatory M1, and immune regulatory M2 macrophages (Gordon 2003; Mantovani et al. 2004). The latter encompass a range of different macrophages with diverse functions including immune suppression, scavenging of apoptotic cells, and tissue remodeling. Based on gene expression analysis, decidual macrophages can be classified as M2 macrophages, with functions particularly related to immune modulation and tissue remodeling (Gustafsson et al. 2008).
Decidual macrophages (CD14+HLA-DR+) express typical M2-associated receptors, including CD163 (scavenger receptor), CD206 (mannose receptor), and CD209 (DC-SIGN; Kammerer et al. 2003; Laskarin et al. 2005; Repnik et al. 2008; Svensson et al. 2011), low levels of the costimulatory CD86 and M1-promoting transcription factor IRF5 (Heikkinen et al. 2003; Svensson et al. 2011), and produce IL-10 (Heikkinen et al. 2003; Lidstrom et al. 2003; Svensson et al. 2011; Figure 1). In particular, decidual macrophages seem to belong to a subset of M2 macrophages with homeostatic and tolerogenic properties, regulated by macrophage colony-stimulating factor (M-CSF) and IL-10, rather than macrophages polarized by Th2-associated cytokines (Svensson et al. 2011). This is supported by their expression of CD163, a homeostatic receptor expressed by tissue resident macrophages (Akila et al. 2012), and the production of CCL18 (Gustafsson et al. 2008), associated with IL-10 and Treg cells (Ambarus et al. 2012; Svensson et al. 2011; Tiemessen et al. 2007). Although few studies have addressed the functional properties of decidual macrophages, they have indeed been shown to be suppressive (Mizuno, Aoki, and Kimbara 1994) and were also recently demonstrated to induce Treg cells, in a dNK cell–dependent manner (Vacca et al. 2010). Nevertheless, the expression of pattern recognition receptors (CD163, CD206, and CD209) also suggests that decidual macrophages have the potential to clear infections, a function that was demonstrated by Singh et al. (2005). Interestingly, M1-associated genes (e.g., IL1B, IL12RB2) appear to be hypermethylated, while M2-associated genes (e.g., A2M, IL10) are hypomethylated, indicating that the anti-inflammatory machinery of macrophages at the fetal–maternal interface is kept more accessible by epigenetic regulation (Kim et al. 2012). Together, these characteristics may be essential for the ability of decidual macrophages to protect the fetus against invading pathogens while maintaining the homeostatic environment required for successful fetal development.
Decidual macrophages have also been attributed a role in spiral artery remodeling due to their production of molecules associated with tissue remodeling and angiogenesis (e.g., matrix metalloproteinase 9 [MMP9] and VEGF; Engert et al. 2007; Gustafsson et al. 2008). MMP9+ macrophages with phagocytic activity were found to infiltrate remodeling decidual vessels suggesting that they are involved in the degradation of the extracellular matrix as well as the clearance of apoptotic cells, which are critical processes during vascular remodeling (Hazan et al. 2010). The role of decidual macrophages in the removal of apoptotic debris (Abrahams et al. 2004; Mor and Abrahams 2003) is consistent with their homeostatic properties. During this process, molecules such as fibronectin-1 and the complement component C1q (Gustafsson et al. 2008) could facilitate the uptake of apoptotic cells that might otherwise create an unfavorable inflammatory environment in the uterus.
The contribution of decidual macrophages to pregnancy complications is not well characterized and the reported macrophage numbers associated with, for example, preeclampsia are inconsistent (Kim et al. 2007; Lockwood et al. 2006). However, the findings of elevated decidual granulocyte-macrophage colony-stimulating factor (GM-CSF levels; Huang et al. 2010), and reduced decidual and placental IL-10 in preeclamptic patients (Hennessy et al. 1999; Schonkeren et al. 2011) are consistent with decreased numbers of regulatory CD163+ macrophages in preeclamptic deciduas (Schonkeren et al. 2011). These data implicate a defect M2 macrophage polarization in the development of preeclampsia, and support a role for IL-10 in promoting, and for GM-CSF in counteracting, the induction of homeostatic decidual macrophages (Svensson et al. 2011). Supporting this, a recent study indicated that altered M2 macrophage polarization in early pregnancy may predispose to complications later in pregnancy (Prins et al. 2012).
Cells of the Adaptive Immune System
T cells in the Decidua
As mentioned, there is a low frequency of T cells in the decidua relative to blood. Furthermore, the composition of T cell subsets is completely different. In blood, most T cells (>90%) are conventional CD4+ or CD8+ T cells expressing the α/β T cell receptor, whereas in decidua there is enrichment of a population of CD4−/CD8− (double negative) T cells expressing the γ/δ T cell receptor (Fan et al. 2011; Mincheva-Nilsson et al. 1994). However, this very high frequency of γ/δ T cells (30–50% of the T cells) has been challenged (Williams, Searle et al. 2009). γ/δ T cells, which typically reside in mucosal sites, show a very restricted T cell repertoire and do not develop memory. Consequently, γ/δ T cells may be considered part of the innate immune system. In the decidua, the γ/δ T cells are activated and despite their morphology with cytoplasmic granulae, they do not normally perform cytolytic activity (Mincheva-Nilsson, Hammarstrom, and Hammarstrom 1992). Instead, they may contribute to fetal immune tolerance by secreting cytokines like IL-10 and transforming growth factor (TGF)-β (Fan et al. 2011; Nagaeva, Jonsson, and Mincheva-Nilsson 2002) and by regulating trophoblast invasion and proliferation, at least in vitro (Fan et al. 2011).
While γ/δ T cells respond to general danger signals, the conventional α/β T cells of the adaptive immune system respond to specific antigens. Among these adaptive T cells, regulatory forkhead box p3 (Foxp3)+CD4+ T cells, which are enriched in the decidua, have drawn much attention (Ernerudh, Berg, and Mjosberg 2011). Regarding the specificity of Treg cells there is growing evidence, in particular derived from mouse models, that they are induced by paternal antigens (reviewed in Guerin, Prins, and Robertson 2009; Leber, Teles, and Zenclussen 2010), with some supportive evidence from studies in humans regarding recognition of paternal antigens (Mjosberg et al. 2007) and enrichment of fetus-specific cells in decidua (Tilburgs et al. 2008). Interestingly, Treg cell induction is enhanced by exposure to semen, thus creating a specific immune tolerance that contributes to a successful subsequent pregnancy. These observations can explain that the risk of preeclampsia is lowered, the more exposed the woman is to paternal antigens (Kho et al. 2009).
The Treg cells are not only enriched in the decidua, they also show a more pronounced suppressive phenotype than in blood with regard to expression of, for example, Foxp3, cytotoxic T lymphocyte antigen 4 (CTLA-4), CD25, and TGF-β (Dimova et al. 2011; Mjosberg et al. 2010). The recruitment of decidual T cells is governed by human chorionic gonadotropin (Schumacher et al. 2009) as well as by the expression of chemokine receptors and the production of corresponding ligands in the decidua, including CCR5/CCL4 (Kallikourdis et al. 2007) and CCR4/CCL17 (Mjosberg et al. 2010). In addition, Treg cells may proliferate (Mjosberg et al. 2010) and mature (Dimova et al. 2011) locally. An intriguing mechanism for regulating T cell traffic was recently shown; stroma cells in the decidua downregulated, by epigenetic mechanisms, their expression of chemokines CXCL9-11, known to recruit Th1 cells (Nancy et al. 2012). Whether similar mechanisms exist for other populations, thereby creating the tolerogenic T cell setting, remains to be seen. In normal pregnancy, Treg and Th2 cells predominate over Th1 and Th17 cells, while a skewing of this balance seems to be involved in complications of pregnancy, as shown in part by mouse and in vitro studies (Ernerudh, Berg, and Mjosberg 2011; Saito et al. 2010). In addition to perturbations of Treg cells, other CD4+ cells as well as CD8+ cells, can contribute to pregnancy complications, for example, based on mismatches in HLA-C, which is expressed by trophoblast cells (for review, see Scherjon et al. 2011).
Conclusion
Human pregnancy is associated with immune adaptations that are essential for successful fetal and placental development. Although changes occur both systemically and locally (mainly in the placental bed), the immune changes observed in the placental bed are more marked than in the maternal circulation. The decrease of certain populations in blood, for example, Treg cells, NK cells, and classical monocytes, may in part be explained by their recruitment to the fetal–maternal interface where these populations are enriched. Consequently, the enhanced activation of innate immunity in the circulation (e.g., increase in nonclassical monocytes and granulocytes) might be a compensation mechanism necessary for maintaining a protective immunity against infections, and thus maternal health. Both systemic and local changes might be due to factors produced by the placenta and/or by direct contact of immune cells with placental trophoblast cells. Although there is still much to be settled, increasing evidence suggests that failure of the immune system to adapt adequately may contribute to pregnancy complications. Maladaptation may be spontaneous or caused by interference of the immune response by toxic substances, which could disturb the pregnancy-specific immune balance, placentation, and thus the outcome or course of pregnancy. It would be of great value to increase our understanding of the intricate interplay between maternal immune cells in the decidua and the role of placental trophoblast cells in their regulation.
Footnotes
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) received no financial support for the research, authorship, and/or publication of this article.