
Abstract
Alternate transgenic mouse models are accepted as replacements for the standard carcinogenicity mouse bioassay by regulatory agencies with a companion 2-year rat bioassay. The slower rate of industry acceptance of these shorter transgenic mouse cancer bioassays has been due to lack of historical data and diagnostic criteria, and the use of nonstandardized terminologies in published data. To address these issues, especially that of generating a large historical database, a retrospective analysis of the spontaneous tumor incidences in rasH2 mice from internally sponsored 6-month carcinogenicity studies was compared to the published literature. Incidences of common spontaneous tumors (incidences > 1%) observed in these studies were lung bronchiolo-alveolar adenomas (mean 3.9–9.9%; range 0–18%), lung bronchiolo-alveolar adenocarcinomas (mean 1.4–2.4%; range 0–5%), splenic hemangiosarcomas (mean 3.0–3.9%; range 0–17%), cutaneous squamous cell papillomas (mean 1.1–1.2%; range 0–4%), Harderian gland adenoma (mean 0.8–1.2%; range 0–4%), and hepatocellular adenomas (mean 1.8%; 0–9% in males only). The remarkable similarity in the tumor incidences in multiple rasH2 studies over a decade and the observed stability of the inserted human gene are important indicators of the minimal drift in this model. Overall, the historical control data for spontaneous neoplasms should assist in the interpretation of future rasH2 mouse studies.
Introduction
For cancer hazard identification of pharmaceuticals, alternate short- or medium-term mouse models including the p53+/– mouse and rasH2 mouse are generally accepted as possible replacements of the standard lifetime carcinogenicity mouse bioassay by the International Conference on Harmonization (ICH) and the U.S. Food and Drug Administration (FDA) (Anonymous 1998). The use of these alternate mouse models can provide significant advantages with similar predictivity for human carcinogens, fewer irrelevant positive results, more flexible resourcing, decreased animal use, shorter testing duration and reduced costs (Alden, Smith, and Morton 2002).
Among the alternate shorter-term mouse models, p53+/– and rasH2 models appear to be the most commonly used (Long et al. 2010). p53+/– mice are generally accepted only for genotoxic pharmaceuticals, while rasH2 mice are recommended for both non-genotoxic and genotoxic pharmaceuticals (Long et al. 2010; MacDonald et al. 2004). The transgenic rasH2 mouse model was initially developed in Japan to evaluate the role of ras over expression in tumorigenesis (Saitoh et al. 1990). The rasH2 mouse, officially known as CByB6F1-Tg(HRAS)2Jic, is a hemizygous transgenic mouse carrying the human c-Ha-ras gene with its own promoter/enhancer elements and harboring a single point mutation in the 3′-end (in the last intron), which induces high enhancer activity (Morton et al. 2002; Suemizu et al. 2002; Tamaoki 2001). This prototypic human transgene, which encodes for the p21ras protein, was noted to be inserted tandemly (3–6 copies) in the murine chromosome 15E3 with approximately 2- to 3-fold the levels found in wild-type mice. The hemizygous transgenic mice are produced by breeding transgenic male C57BL/6 J mice with wild type BALB/CByJ female mice.
Major gaps identified in the use of these alternate mouse models (as gauged by a recently published survey of the Society of Toxicologic Pathology membership) include issues with study interpretation primarily due to lack of historical background data and issues with tumor identification and characterization of pre-neoplastic lesions (Long et al. 2010). Although there are several reports of control data from rasH2 mice, they have typically been conducted in multiple laboratories, are several years old, have generally lacked details on study-design parameters such as age of animals, study duration, dosing procedures, laboratory practices, and pathology diagnostic criteria, and often have interpathologist variability.
The major objectives of this study were to generate a control database of spontaneous tumors in rasH2 mice using consistent diagnostic criteria/nomenclature and to compare it with previous rasH2 literature. In this report compiled from eleven carcinogenicity studies, retrospective analysis of the background tumor prevalence in untreated rasH2 hemizygous transgenic mice was performed by an individual veterinary pathologist. Furthermore, the background tumor incidences in these rasH2 mice were compared to the incidences published in the literature. Comparison between Pfizer studies and published studies also included MNU-treated tumor incidences. Thus, this is a first collation of data from a large number of rasH2 mice (N = 333–363) from studies conducted with pharmaceuticals or studies designed to generate historical control data. Some of the key highlights of this retrospective analysis are adherence to the best practice guidelines for historical control data such as detailed information of study design parameters, relatively shorter time interval for data collection (4–5 years), and peer review of the data by an individual pathologist to ensure consistent application of diagnostic criteria and nomenclature (Keenan et al. 2009).
Materials and Methods
Study Design
Data reported in this article are from control/vehicle groups from 10 carcinogenicity studies for males and 11 studies for females of 6-month dosing duration conducted between March 2004 and July 2009 under good laboratory practice (GLP) conditions. All studies and animal procedures were conducted in accordance with animal use protocols approved by the animal care and use committee in compliance with The Guide for Care and Use of Laboratory Animals and all applicable regulations. CB6F1/Jic-Tg (rasH2-Tg) male and female mice were purchased from Taconic (Germantown, NY) at 6 weeks of age and housed individually in suspended polycarbonate or stainless steel wire cages in an environmentally controlled room (temperature 18–26°C, relative humidity 30%–70%, controlled 12-hour light cycle) and provided ad libitum access to water and a certified rodent diet. Each mouse was genotyped before leaving the vendor’s facility. The mice were typically allowed to acclimate for 2 weeks before study start, and weighed between 17.6 to 25.4 g at study initiation. The total number of control males and control females evaluated in these carcinogenicity studies were 333 and 363, respectively. The number of control animals in each study ranged from 25 to 90/sex. These control mice received either certified rodent diet (n = 4 diet admix studies) alone or a vehicle such as 0.5% methylcellulose ± Tween or sterile water via oral gavage (n = 7 studies). The alkylating carcinogen, N-methyl-N-nitrosourea (MNU), was used as a positive control in 8 out of 11 studies. A single dose of MNU pH 4.0 in citrate buffer was given at 75 mg/kg IP (n = 7 studies) and at 58 mg/kg IP (n = 1 study). Only 6 of these 8 studies were used for MNU-related incidence reporting, as only these 6 included evaluation of a full tissue list. The numbers of rasH2 mice that were evaluated from the MNU-treated groups were 87/sex.
Pathology Evaluation
All surviving animals were humanely euthanized at the end of each 6-month study. A complete necropsy was performed with fixation of most tissues in 10% neutral buffered formalin followed by embedding in paraffin, sectioning at 3–5 μm and staining with hematoxylin and eosin (H&E) using standard procedures. Tissues such as eyes and testes/epididymides were fixed in 3% glutaraldehyde or Davidson’s solution, respectively. The H&E stained tissue sections from the control animals were retrospectively evaluated by a board-certified veterinary pathologist using published nomenclature and criteria (Mohr 2001). The standard tissues collected and evaluated in these studies are included in Table 1. Individual control mice that were euthanized moribund or found dead prior to scheduled necropsy were also included in the data.
General list of tissues examined.
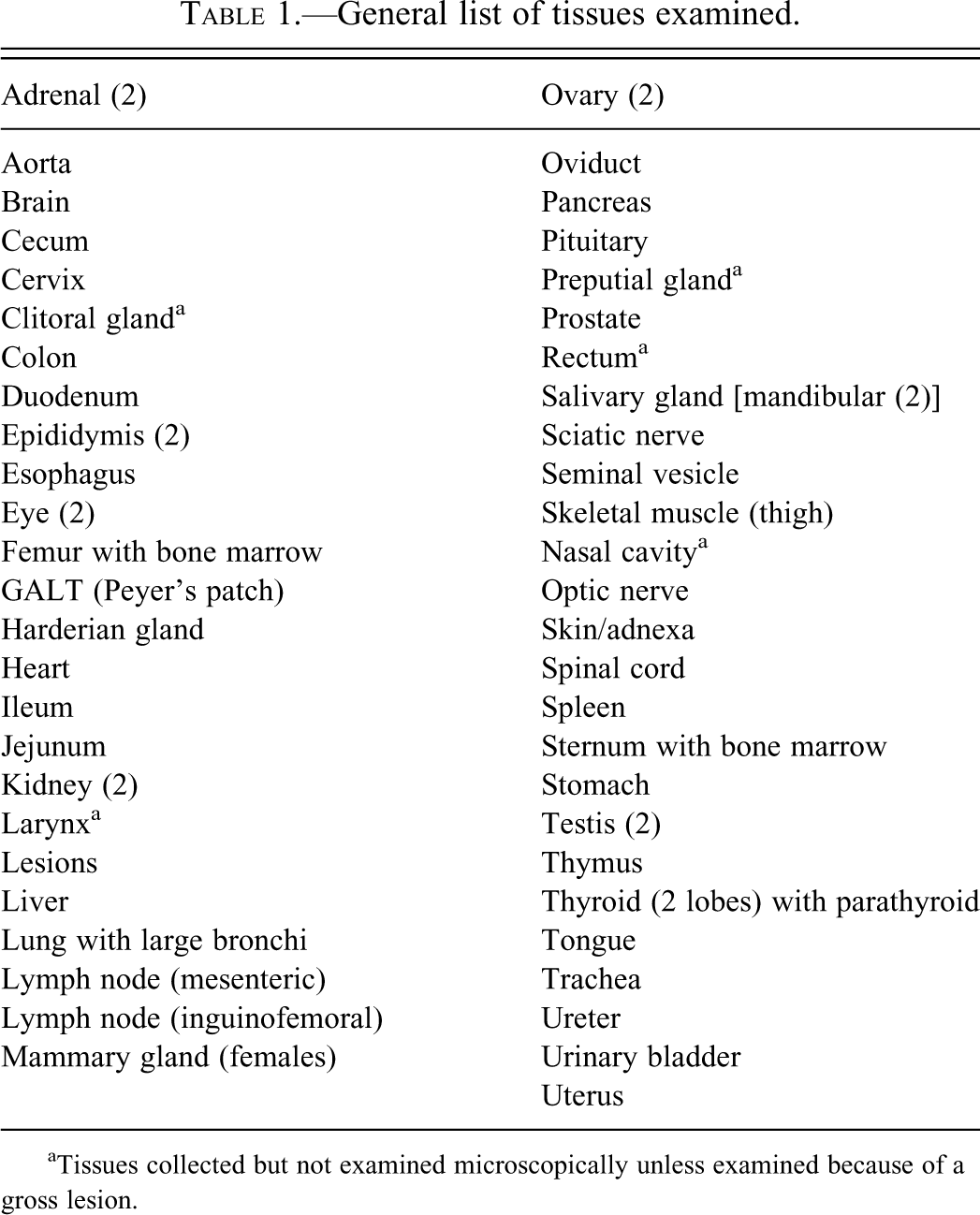
aTissues collected but not examined microscopically unless examined because of a gross lesion.
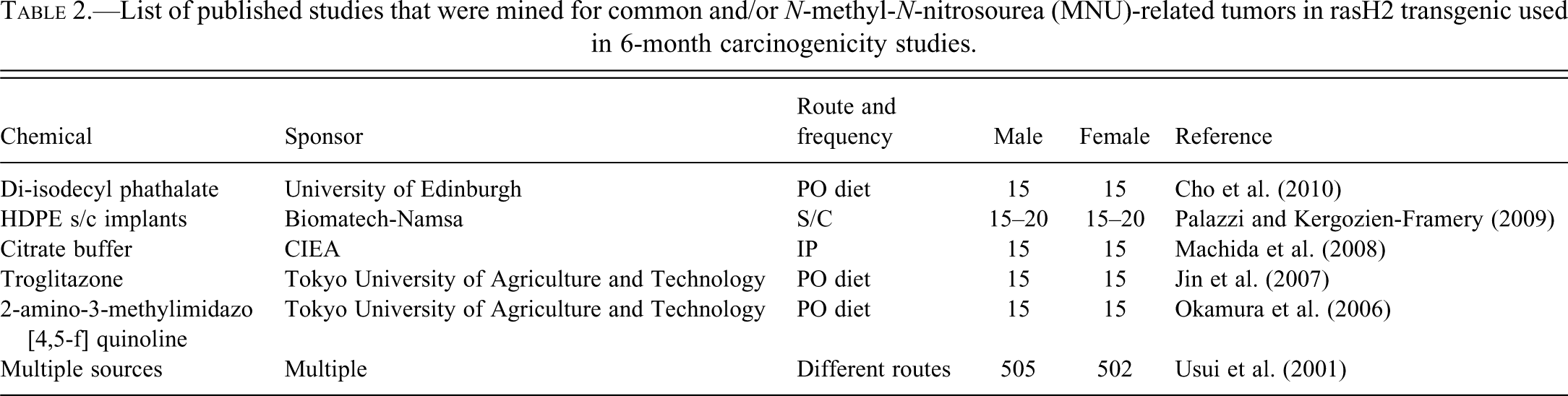
To compare the internally generated tumor incidence data with previous reports in rasH2 mice, tumor incidence from control and MNU-treated rasH2 mice from 6-month carcinogenicity studies were also collated from literature published from 2001–2011 (Table 2) (Cho et al. 2010; Jin et al. 2007; Machida et al. 2008; Okamura et al. 2006, 2007; Palazzi and Kergozien-Framery 2009; Usui et al. 2001). It is important to note that MNU-related tumor incidences in the Pfizer studies were obtained from study reports and not by retrospective analysis of slides.
List of published studies that were mined for common and/or N-methyl-N-nitrosourea (MNU)-related tumors in rasH2 transgenic used in 6-month carcinogenicity studies.
Results
Body Weight and Survival Rate
The terminal body weight of male and female rasH2 mice in the Pfizer studies ranged from 28.4–34.5 and 22.9–25.1 g, respectively (Table 3). Over a period of 6 months, the body weight gain, as expected, was slightly higher in the males (4.6–9.0 g) than females (4.1–6.2 g). The survival rate in these mice at the end of 6 months was high (>95% average) with a range between 92–100%. Cause of death or early euthanasia was due to hemangiosarcomas, pulmonary bronchiolo-alveolar tumors, squamous cell carcinoma, urinary tract obstruction, preputial abscess, and so forth (Table 3).
Body weight and survival for untreated/vehicle rasH2 mice at end of 6-month study (Pfizer Studies).
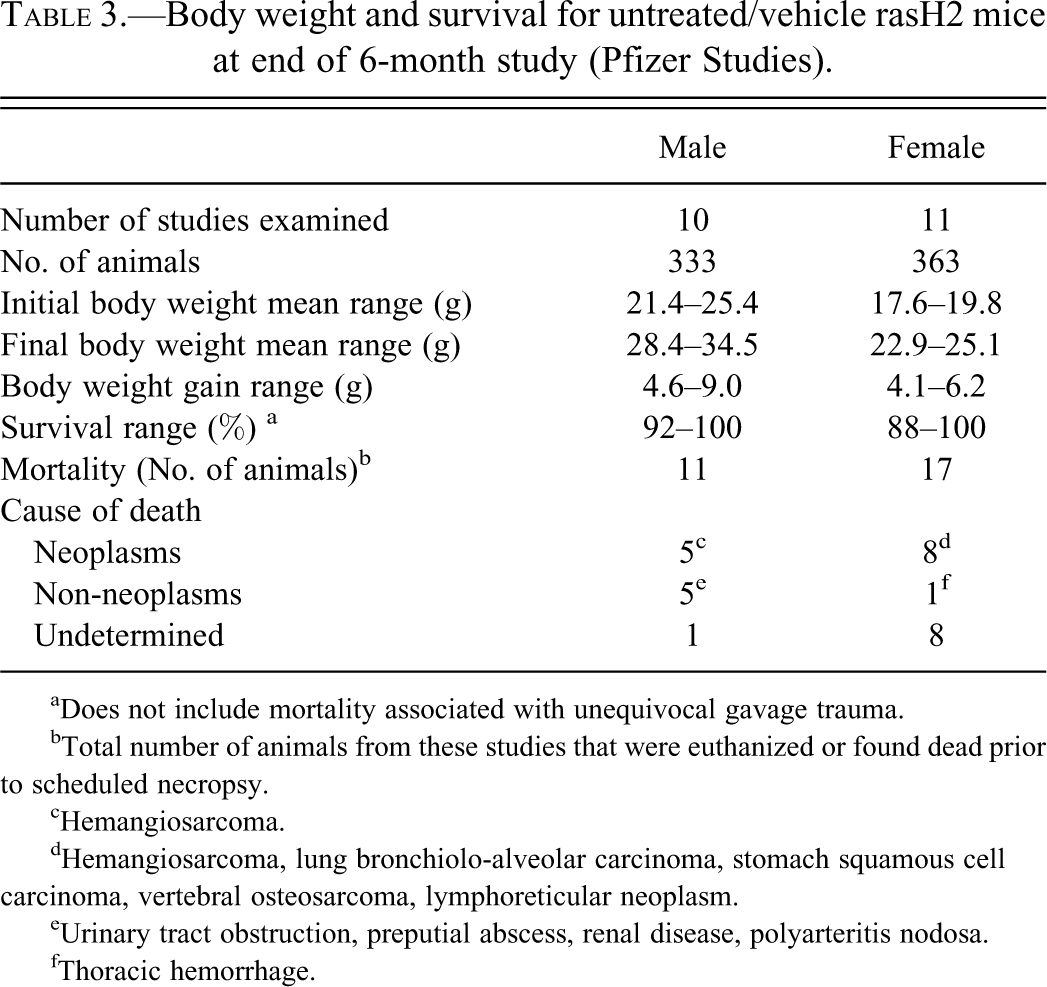
aDoes not include mortality associated with unequivocal gavage trauma.
bTotal number of animals from these studies that were euthanized or found dead prior to scheduled necropsy.
cHemangiosarcoma.
dHemangiosarcoma, lung bronchiolo-alveolar carcinoma, stomach squamous cell carcinoma, vertebral osteosarcoma, lymphoreticular neoplasm.
eUrinary tract obstruction, preputial abscess, renal disease, polyarteritis nodosa.
fThoracic hemorrhage.
Spontaneous Tumor Incidence in rasH2 Mice
Spontaneous tumor incidences in control rasH2 mice were evaluated from the Pfizer 6-month carcinogenicity studies. The common tumors, defined as tumors with incidence ≥ 1% over all studies, are listed in Table 4. A comprehensive list of the tumor incidences in these mice is provided in the supplementary table (S1). The mean incidence of common tumors observed in these studies were lung bronchiolo-alveolar adenomas / adenocarcinomas (1.4–9.9%), splenic hemangiosarcomas (3.0–3.9%), cutaneous squamous cell papillomas (1.1–1.2%), Harderian gland adenoma (0.8–1.2%), and hepatocellular adenomas (1.8% in males only). Isolated occurrences of hemangiosarcomas in individual organs (such as testes, skin, kidney, bone marrow, etc.) were observed in individual animals across studies. When combined (as they were mutually exclusive), the incidences of hemangiosarcomas in all organs in both males and females were 5.1% and 6.1%, respectively. The tumor incidences were similar between males and females except for pulmonary bronchiolo-alveolar adenoma incidence, which had a higher incidence in males (9.9%) compared to females (3.9%), and liver adenomas which had a higher incidence in males (1.8%) compared to females (0%). The histologic phenotype of these tumors was based on previously reported criteria and was similar to those described in rasH2 mice previously (Mitsumori et al. 1998; Mohr 2001).
Background incidences of common tumors in rasH2 transgenic mice (from Pfizer Studies).
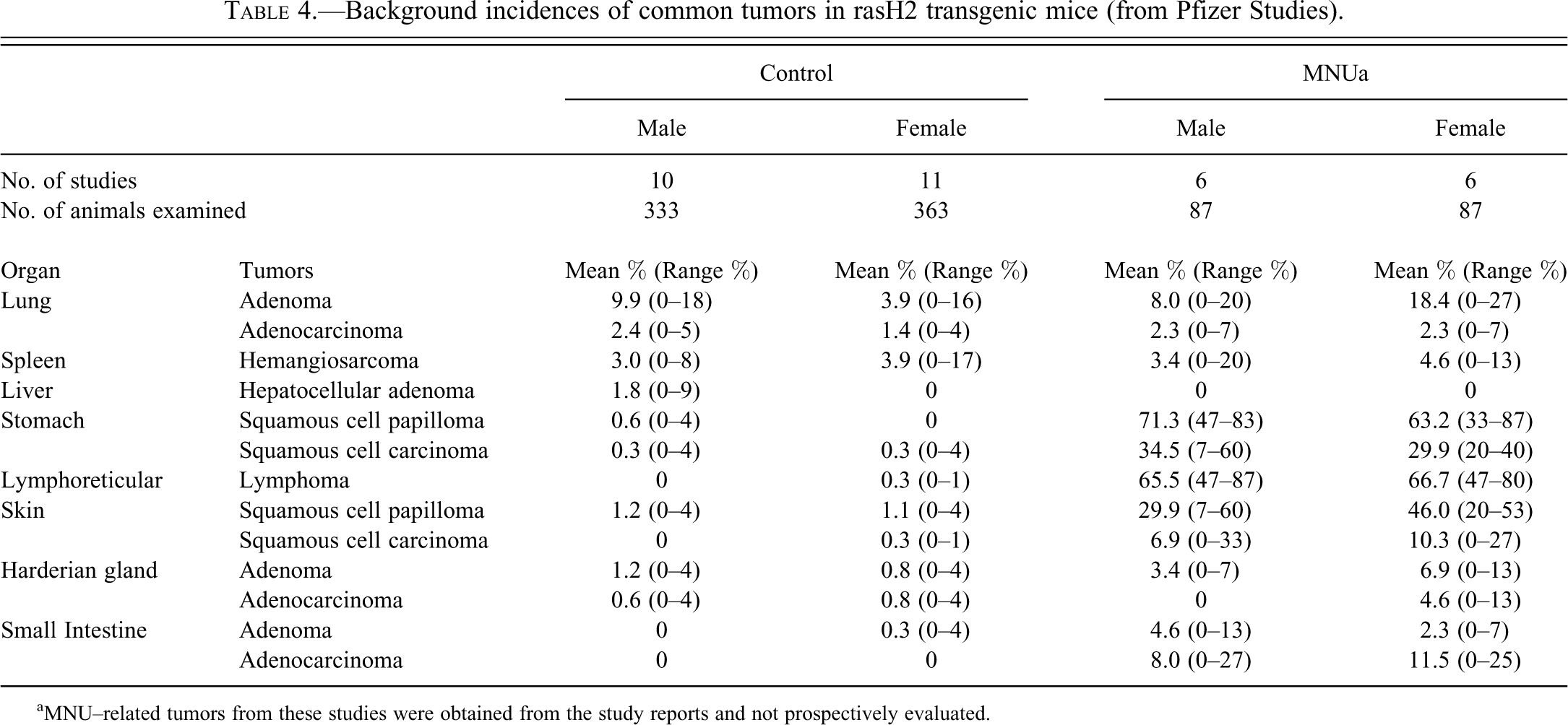
aMNU–related tumors from these studies were obtained from the study reports and not prospectively evaluated.
To compare the spontaneous tumor incidence in the Pfizer studies with those reported previously, we reviewed the literature for rasH2 6-month studies spanning from 2000 to 2011 (Table 2). Overall, the spontaneous tumor incidence in Pfizer studies was generally comparable to those reported previously (Table 5). For example, the mean incidences of the most common tumors for both sexes reported in the literature include pulmonary bronchiolo-alveolar adenomas (5.3–5.4%), splenic hemangiosarcomas (3.0–3.7%), cutaneous squamous cell papillomas (1.3–1.8%), lymphomas (0.3–2.4%), and gastric squamous cell papillomas (0.6–1.3%). The primary differences noted in the Pfizer data and the published literature include the incidence of lung bronchiolo-alveolar adenomas/adenocarcinomas in both sexes, which were slightly higher (5.3–12.3%) in the Pfizer studies compared to the literature (5.9–6.4%; Table 5). Furthermore, the incidence of lymphomas in females was relatively low (0.3%) in the Pfizer studies compared to previous reports (2.4%; Tables 4, 5).
Percentage incidences of the common and N–methyl–N–nitrosourea (MNU)–related tumors (> 1% incidence) in rasH2 transgenic mice used in 6–month carcinogenicity studies (from literature).
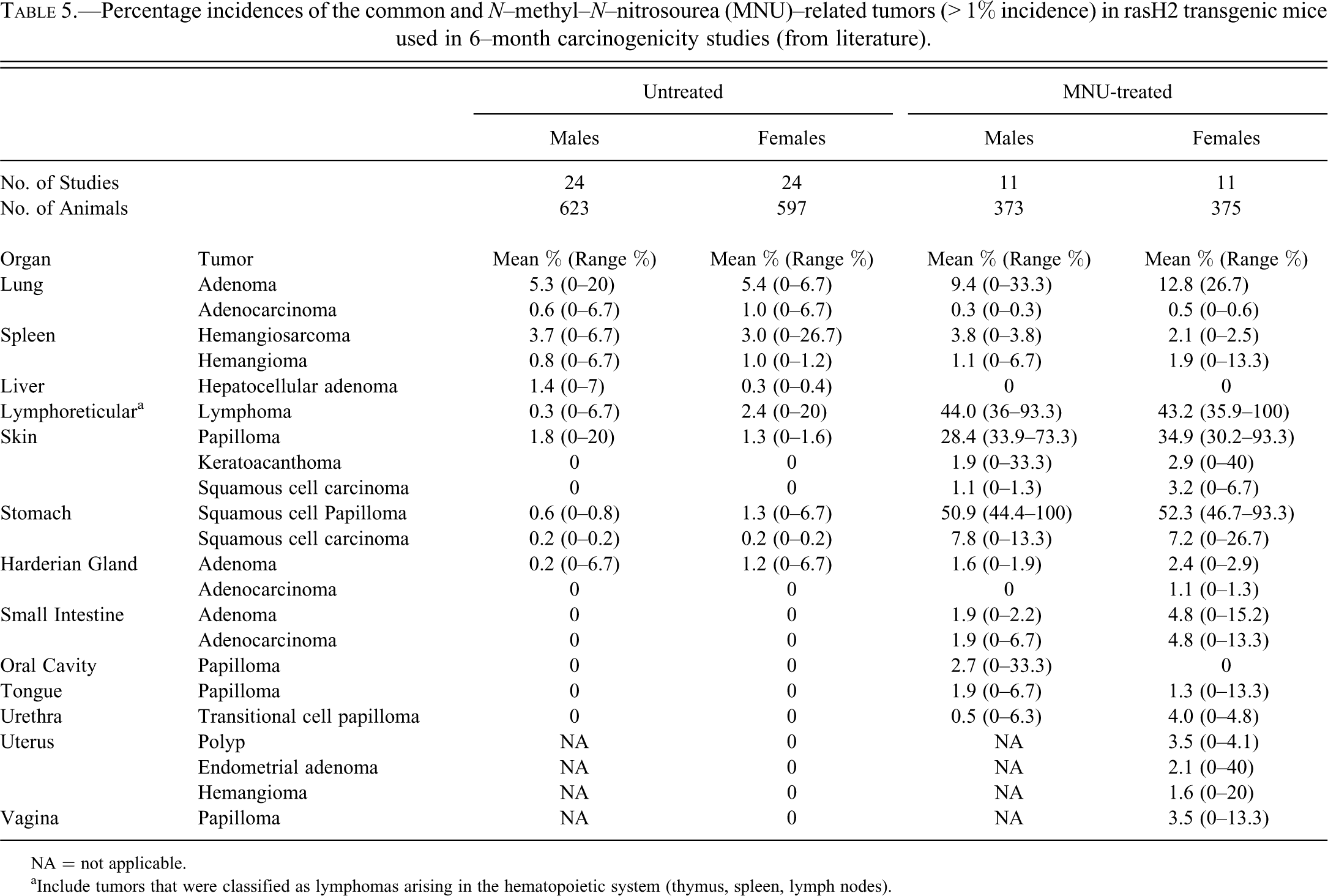
NA = not applicable.
aInclude tumors that were classified as lymphomas arising in the hematopoietic system (thymus, spleen, lymph nodes).
Histologic Phenotype
A brief discussion of the key diagnostic features of the common tumors in rasH2 mice based on previously published criteria (Mohr 2001; Renne et al. 2009) are presented below.
Pulmonary bronchiolo-alveolar adenomas were typically small in size (< 3–4 mm diameter), discrete without encapsulation, and often located on the peripheral border of the lung lobes. The tumor cells were arranged in glandular or papillary patterns with frequent compression of the adjacent normal alveoli. The tumor cells were uniformly cuboidal to columnar with basophilic cytoplasm and supported by a thin fibrovascular stroma. Mitotic figures were rare or absent (Figure 1).
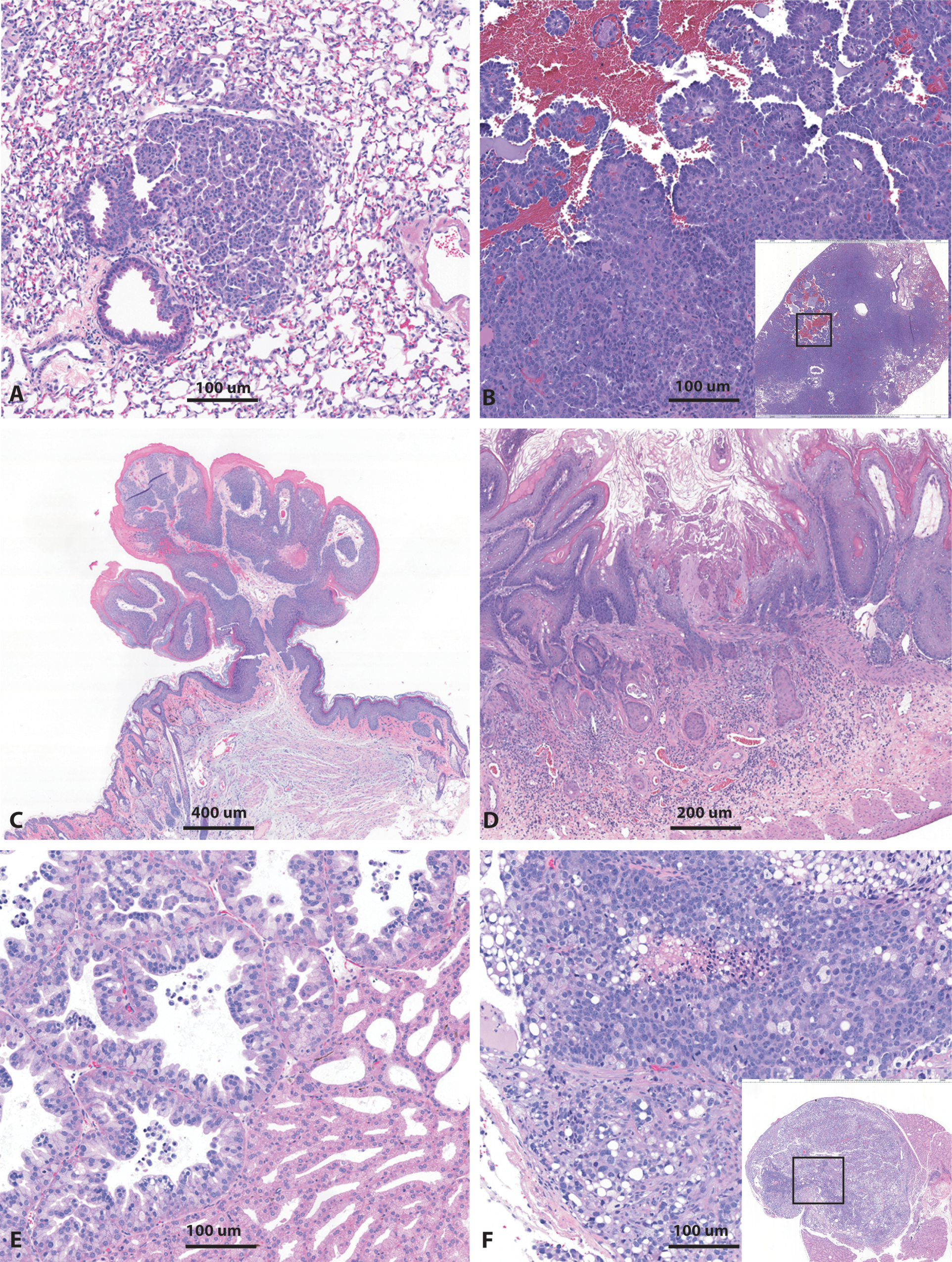
Representative photomicrographs of spontaneous neoplasms noted in rasH2 mice in 6-month carcinogenicity studies. These include pulmonary bronchiolo-alveolar adenoma (A) and adenocarcinomas (B), cutaneous squamous cell papilloma (C), gastric squamous cell carcinoma (D), Harderian gland adenoma (E), and adenocarcinoma (F). Insets represent low-magnification images of the neoplasm while the boxed area is the magnified portion of the region. H&E stain.
The pulmonary bronchiolo-alveolar carcinomas were mostly poorly demarcated neoplasms with diameter often > 3–4 mm and occupying an entire lobe with distortion of and invasion into underlying architecture. The tumor cells were cuboidal to columnar with anisocytosis and anisokaryosis and often arranged in solid glandular/tubular and papillary patterns. Occasional infiltration into bronchioles and lymphatics were also noted. Increased inflammatory cells (macrophages) and loosely attached neoplastic cells were noted within the alveoli adjacent to the tumors (Figure 1).
Splenic hemangiosarcomas were often expansile infiltrative masses with effacement of the underlying normal structures. These tumors were hypercellular and composed of elongate to plump atypical cells with large anisokaryotic nuclei forming vascular channels separated by varying amount of connective tissue as well as solid areas with little evidence of vascular channels and composed of atypical elongate to plump neoplastic cells arranged in a storiform pattern. Mitotic figures were frequent (Figure 2). Metastases of these tumors were uncommon.
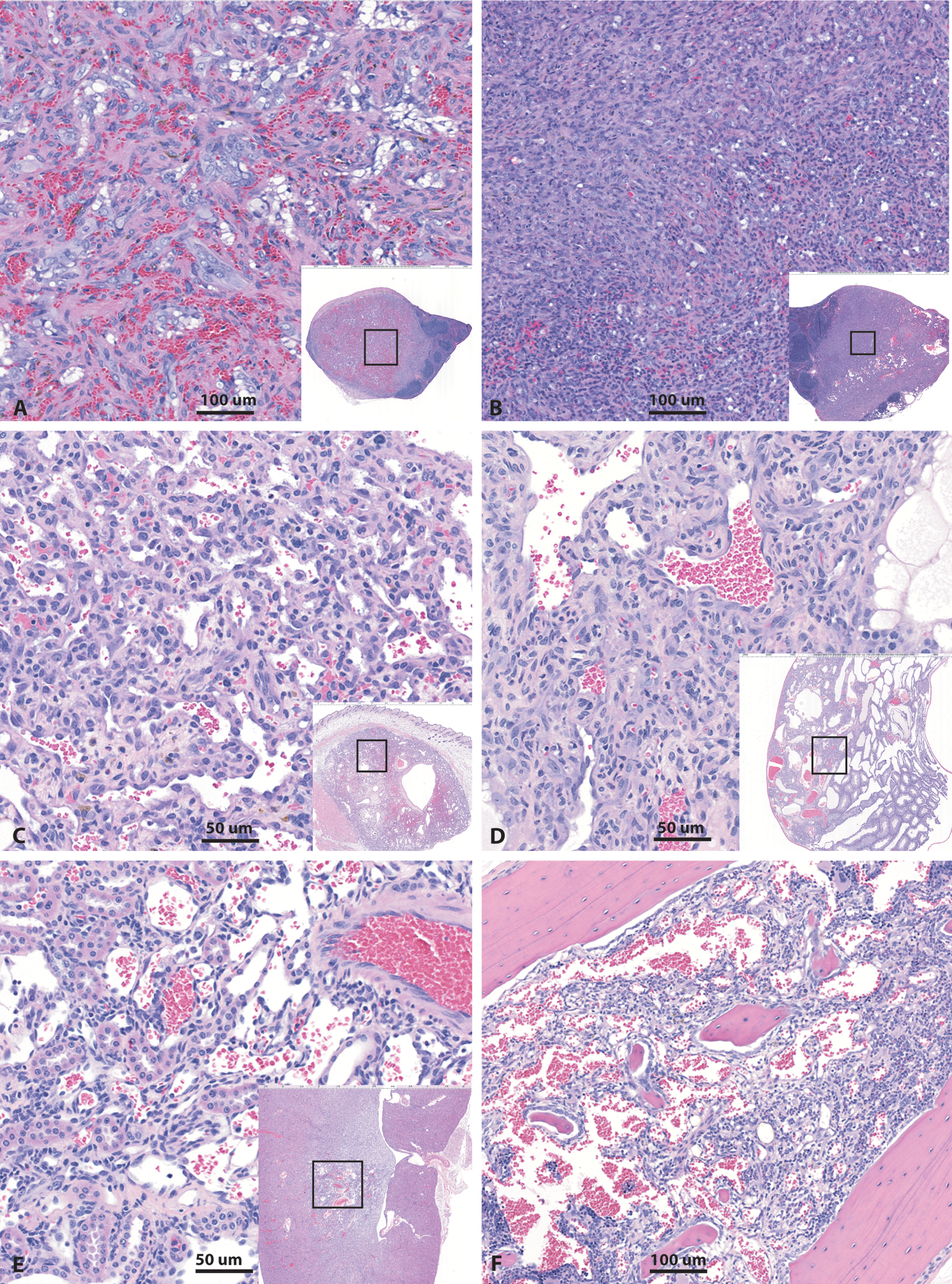
Representative photomicrographs of hemangiosarcomas noted in rasH2 mice in 6-month carcinogenicity studies. These tumors were most common in the spleen (A, B), while isolated occurrences were noted in skin (C), testis (D), kidney (E), bone marrow (F), and occasionally other organs, independent of the spleen. Note the two distinct patterns of splenic hemangiosarcomas–neoplastic cells forming vascular channels (A) and the more solid fibrous component (B). In other organs, the hemangiosarcomas typically formed vascular channels. Insets represent low magnification images of the neoplasm, while the boxed area is the magnified portion of the region. H&E stain.
Hepatocellular adenomas were well demarcated and slightly compressed the adjacent hepatic parenchyma. The cells within the neoplasm were uniform, well differentiated with uniform nuclei forming regular hepatocyte plates, and lacking lobular architecture (i.e., portal triads, central veins, etc.). The hepatocyte cytoplasm ranged from eosinophilic, basophilic, and clear to vacuolated.
Cutaneous papillomas were solitary, mostly exophytic, well-circumscribed neoplasms with single or multiple finger-like projections and a fibrovascular connective tissue core. The overlying epidermis was composed of hyperplastic stratified squamous epithelium and there was no evidence of dermal invasion (Figure 1).
Gastric squamous cell papillomas were usually solitary, exophytic masses arising within the squamous/non-glandular stomach and composed of a thin fibrovascular connective tissue core. The epithelium was frequently hyperplastic with normal maturation and no evidence of proprial invasion.
Harderian gland adenomas were usually isolated, variable sized, well-demarcated tumors composed of acinar glands lined by a single layer of finely vacuolated/foamy cells with projection into the lumen (Figure 1). Harderian gland adenocarcinomas were expansile infiltrative tumors, poorly demarcated with disruption of normal architecture. The neoplastic cells formed solid glandular structures often piling up to form multiple layers, with a variable degree of cytoplasmic vacuolation, and prominent oval to round nuclei. The mitotic figures were increased and occasional necrotic foci were noted within the neoplasm (Figure 1).
The intestinal neoplasms noted in these rasH2 mice were adenomas (Figure 3).
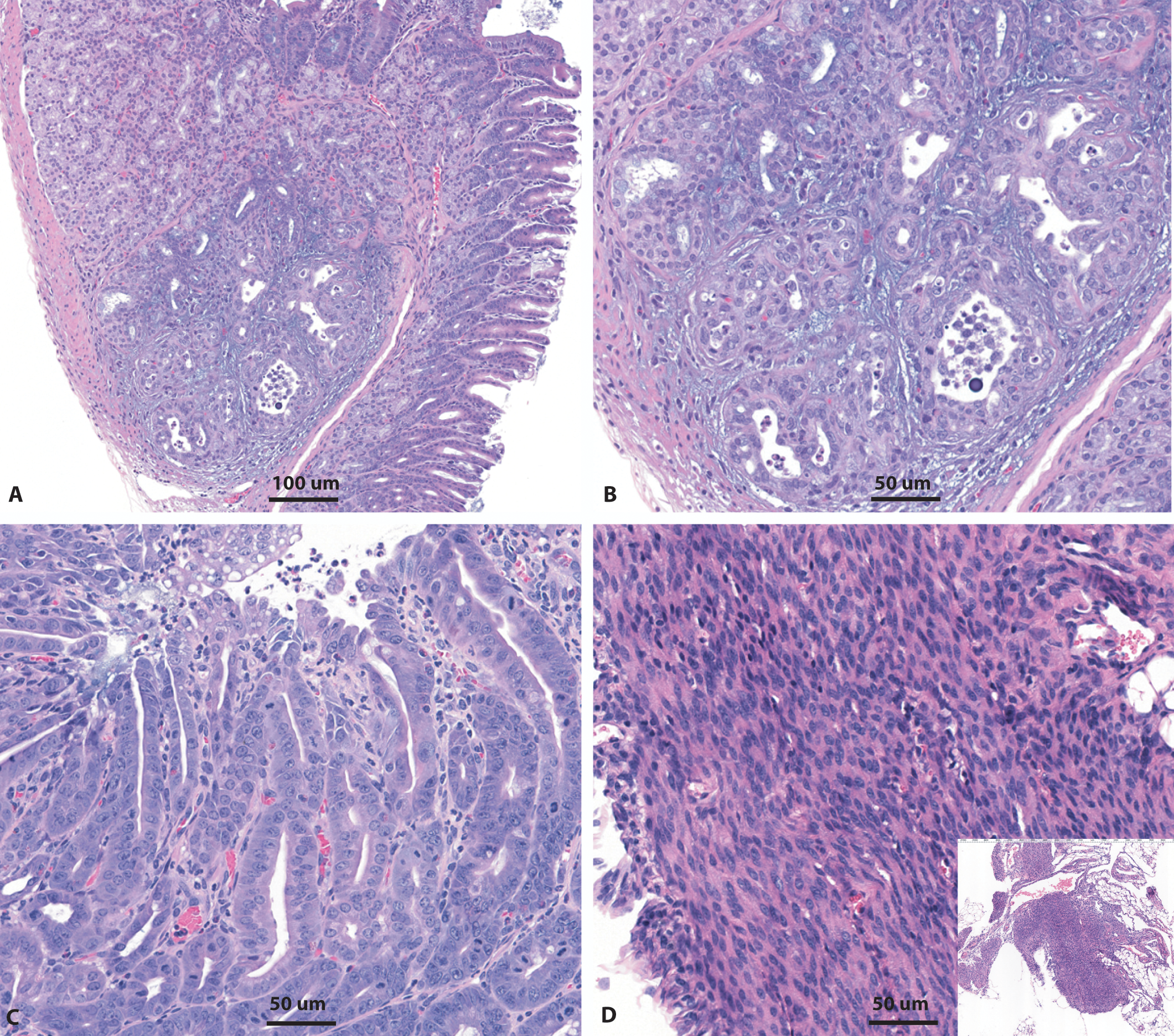
Representative photomicrographs of rare neoplasms (<1% incidence) noted in rasH2 mice in 6-month carcinogenicity studies. Adenocarcinoma within the Brunner’s gland in the duodenum (A, B), small intestinal adenoma (C), and a mesenteric leiomyoma (D) are shown. Insets represent low-magnification images of the neoplasm. H&E stain.
Tumor Incidence in the Commonly Used Positive Control, MNU
MNU, a known alkylating genotoxic carcinogen, is often used as the positive control to confirm the sensitivity of the rasH2 mouse model to chemical carcinogens. We therefore evaluated the tumor incidence in MNU-treated (positive control) rasH2 mice and compared our data to the published literature. The target organs, with respect to the observed MNU-related tumors, were comparable in the Pfizer studies and previous reports. In the Pfizer studies, the most common MNU-related tumors in both sexes were gastric papillomas (63.2–71.3%), lymphomas (65.5–66.7%), cutaneous papillomas (29.9–46.0%), gastric squamous cell carcinomas (29.9–34.5%), pulmonary bronchiolo-alveolar adenomas (8.0–18.4%), small intestinal adenocarcinomas (8.0–11.5%), cutaneous squamous cell carcinomas (6.9–10.3%), harderian gland adenomas (3.4–6.9%), and small intestine adenomas (2.3–4.6%; Table 4). MNU-related increases in incidence of squamous cell papillomas, squamous cell carcinomas, hemangiosarcomas, and so forth were also sporadically noted in the female reproductive tract including vagina, cervix, and uterus (data not shown) in both Pfizer studies and previous reports.
Some of the unique MNU-related features in the Pfizer studies relative to previous reports include higher incidence of the more common MNU-related tumors such as lymphomas, gastric papillomas, and cutaneous papillomas. Tumors such as gastric squamous cell carcinoma, cutaneous squamous cell carcinomas, and small intestinal adenocarcinomas were also noted at higher incidences (Tables 4, 5). Similar to previous reports, MNU treatment in the Pfizer studies failed to increase the incidence of splenic hemangiosarcomas, the second most common spontaneous tumor in rasH2 mice.
The most common and consistent MNU-related neoplasm in the Pfizer studies and published literature were gastric papillomas and lymphomas. The incidences across studies ranged from 33–87% and 47–87% for gastric papillomas and lymphomas, respectively. Overall, MNU was a robust positive control eliciting a tumorigenic response comparable to previous reports.
Discussion
The limitations of lifetime rodent bioassays include several factors such as protracted study duration (2–3 years), study size (> 600 animals), cost (~1 MM USD), high morbidity and mortality rates during study, lack of clarity on mechanism of tumorigenesis, and questionable relevance to human risk assessment (Alden et al. 2009). The acceptance of short-term (i.e., 6-month) alternate mouse transgenic models such as rasH2 and p53+/– mice for cancer biohazard testing of pharmaceuticals by the FDA and ICH has been an important milestone from drug development and human risk assessment perspectives (Friedrich and Olejniczak 2011).
In line with regulatory acceptance of the transgenic mouse models, recent trends in the use of these models in carcinogenicity testing have been encouraging. In the last decade (2002–2011), 135 out of 506 mouse carcinogenicity protocols submitted to the FDA were transgenic mouse protocols (ranging from 11–52% per year). Of these 135 transgenic mouse protocols, 78 (57.8%; range 9.1–92.9% per year) were from studies using rasH2 mice. Interestingly, the trends in the use of p53+/– mice and rasH2 mice have reversed in the past 5 years, with an increase in rasH2 mouse protocol submission to the FDA (50–85% per year) relative to p53+/– mice (0–31.3% per year; personal communication). The trend toward greater use of the rasH2 model likely reflects the FDA’s view that the p53+/– mouse is suitable only for pharmaceuticals that are positive in at least one genotoxicity assay.
Despite the positive signs, the adoption of these alternate transgenic mouse models by the pharmaceutical industry has been slow (Friedrich and Olejniczak 2011). Some of the factors contributing to limited use of these models by the biopharmaceutical industry have been concerns regarding lack of historical data, application of consistent diagnostic criteria, use of nonstandardized terminologies in published data, and lack of comfort using a novel model (Long et al. 2010). To address these issues especially that of generating a large historical database, we conducted comprehensive retrospective analysis of the spontaneous tumor incidence in rasH2 mice from internally sponsored 6-month carcinogenicity studies.
Overall, the incidences of tumors in the Pfizer internal rasH2 studies were similar to the compiled incidence from all the published rasH2 studies of similar duration (i.e., 6-month studies). Despite variables such as the wide range of the study conduct dates (i.e., over 10 years), geographical locations, and sponsoring institutions, the tumor spectrum in the published literature was remarkably similar to the Pfizer studies. For example, the common spontaneous tumors noted in rasH2 mice in the Pfizer studies were pulmonary adenomas/carcinomas, splenic hemangiosarcomas, cutaneous squamous cell papillomas, hepatocellular adenomas, and harderian gland adenomas. A few exceptions noted in the Pfizer studies relative to the literature were the low incidence of lymphomas and gastric papillomas and slightly higher incidence of pulmonary tumors. The stability of the inserted human transgene in the mouse genome over several generations (Suemizu et al. 2002; Tamaoki 2001) and the remarkable similarity in the tumor incidences in multiple rasH2 studies over a long interval of time (i.e., decade) demonstrate the robustness and minimal drift in this animal model.
Although the inserted human c-ha-ras transgene is expressed in most tissues of rasH2 mice, tumors develop only in select organs without any direct correlation between transgene expression and tumorigenesis. The mechanistic basis for the higher predilection for pulmonary tumors, hemangiosarcomas, and epithelial tumors in rasH2 mice is unclear. Although protein levels of the human transgene product (p21) and activating mutation of the human transgene play important roles in tumorigenesis, tumorigenesis in rasH2 mice can be independent of mutational activation (Mitsumori 2002). Higher tumor incidence in specific organs or cell types can be a function of the genetic background of the parental strains. The background strains of rasH2 mice are BALB/c and C57BL/6. The spontaneous tumor spectrum in rasH2 mice was roughly in accordance with the background strains of BALB/c and C57BL/6 (Mitsumori 2002; Tamaoki 2001). In fact, the reported incidences of lung adenomas and gastric papillomas in the wild type littermates of the rasH2 mice around 7–9 months of age were 1.7% and 0.6%, respectively (Mitsumori 2002; Tamaoki 2001). Thus, the low spontaneous tumor incidence in untreated rasH2 mice and greater susceptibility to chemical carcinogens highlights the model’s sensitivity to chemical carcinogens.
MNU, an alkylating carcinogen, is the commonly used positive control to demonstrate the enhanced sensitivity of the rasH2 mouse model. To evaluate if MNU causes an increase in the background tumor incidence or causes induction of tumors in organ systems distinct from those in which background tumors are seen, we surveyed the literature for tumor incidences in MNU-treated (single dose) 6-month rasH2 studies. These data were then compared with the data generated in the MNU positive control groups in our 6-month carcinogenicity studies. The most common tumors induced by MNU (gastric papillomas, lymphomas, cutaneous papillomas, and gastric carcinomas) were similar between the two data sets. The incidences of these common MNU-related tumors were slightly higher in Pfizer studies compared to previous reports. These differences in MNU sensitivity may be a function of the preparation and immediate administration of MNU in the laboratory setting. MNU is a highly volatile agent with a short half-life; therefore, careful preparation and immediate administration are critical elements in eliciting a positive tumor response. Overall, MNU was a robust positive control with consistent induction of tumors such as lymphoma (47–87% range) and gastric papilloma (33–87% range), as previously reported (Usui et al. 2001). This high positive tumor rate permits the use of only 10–15 animals/sex in MNU-positive control groups (Morton et al. 2008). For the positive control cohorts, histologic evaluation of thymus alone is recommended while collecting only stomach and tissues with macroscopic findings for further evaluation if needed. The FDA has endorsed this approach for at least 2 pharmaceutical candidates.
Another facet of this animal model, noted in the Pfizer studies and in the literature, was the tumor spectrum (i.e., organ susceptibility) after treatment with MNU. MNU-related tumor spectrum was slightly discordant with the spontaneous/background tumor spectrum. For example, the most common spontaneous tumors in rasH2 mice were pulmonary adenomas, hemangiosarcomas, hepatocellular adenomas, and cutaneous papillomas. MNU administration did not result in a proportional increase in the incidence of these background tumors. For example, the incidence of hemangiosarcomas, the second most common spontaneous tumor in rasH2 mice, did not change after treatment with the alkylating carcinogen MNU. The most common MNU-related tumors were gastric papillomas, lymphomas, cutaneous papillomas, and gastric squamous cell carcinomas. Similarly, urethane (another alkylating genotoxic carcinogen) treatment was associated with a tumor spectrum distinct from MNU, that is, high incidence of lung tumors and splenic hemangiomas / hemangiosarcomas (Ozaki et al. 2005). Although the basis of these differences between spontaneous tumors and different genotoxic agents is unclear, earlier reports have highlighted organ susceptibility and mutational pattern as the two main differences between MNU-related tumors and spontaneous tumors in rasH2 mice (Ando et al. 1992; Saitoh et al. 1990). Thus, it is important to keep in mind that the organ susceptibility in rasH2 mice is dependent on the type of the positive control used (MNU or urethane).
An ideal animal model, as defined by the National Research Council, should be based on “appropriateness as an analog, transferability of information, genetic uniformity, background knowledge of biological properties, generalization of the results, ease of experimental manipulation and ecological consequences and ethical implications” (Wall and Shani 2008). The rasH2 mouse model satisfies most of these requirements as they have been shown to be as effective if not better than the model they have replaced, that is, 2-year mouse bioassay; are robust and reproducible with respect to the tumor incidence; have been shown to be genetically stable over several generations; and have low mortality rates and spontaneous tumor incidence, which could compromise long-term studies. Despite the positive attributes of rasH2 mouse carcinogenicity studies, it is important to note that these studies in transgenic models are always a part of an integrated, weight of evidence evaluation that considers mechanisms of action, genotoxicity, structure activity relationships, toxicokinetic information, and results from other in vitro and in vivo bioassays (MacDonald et al. 2004).
In summary, a 6-month rasH2 mouse carcinogenicity study is an accepted alternative to a 2-year rodent bioassay for cancer hazard identification of genotoxic and non-genotoxic small pharmaceutical molecules when combined with one 2-year rat carcinogenicity study. The rasH2 mouse expresses multiple copies of the human c-ha-ras gene that is aberrantly expressed in many human neoplasms. The rasH2 model predicts known human carcinogens with similar accuracy as 2-year rodent studies, but demonstrates fewer positive results that are considered irrelevant or of low relevance for human risk assessment. Other advantages include reduced animal use, high survival at study termination, greater flexibility in study scheduling, and overall resource savings. The rasH2 mouse model should be considered to support registration of small molecule pharmaceuticals that require carcinogenicity assessment. The standard study design for a rasH2 mouse study includes 3 treatment groups and a vehicle control group composed of 25 animals/sex/group. A positive control group (usually MNU or urethane) composed of at least 10 animals/sex is required. It is acceptable to examine microscopically only the thymus in the positive control (MNU-treated) cohort. Limited historical control data and lack of confidence in interpretation of positive results likely have slowed the use of the rasH2 mouse model by sponsors. The historical control data for spontaneous neoplasms and neoplasia induced by MNU should assist in the interpretation of future rasH2 mouse studies. Overall, the remarkable similarity in the tumor incidences in multiple rasH2 studies over a long interval of time (i.e., decade) further highlights the robustness and minimal drift in this animal model.
Footnotes
Acknowledgments
The authors wish to express their sincere thanks to Drs. Roy Kerlin and Christopher Houle and Ms. Kimberly Ebersole in critically reviewing the article and Walter Bobrowski for help with the images.
The author(s) declared no potential conflicts of interests with respect to the authorship and/or publication of this article. The author(s) received no financial support for the research and/or authorship of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.