
Abstract
Characteristic susceptibility to environmental and pharmaceutical exposure may occur during periods in life of marked histophysiological changes of the immune system. Perinatal development is such a period; pregnancy followed by lactation is potentially another one. Here, we explored the influence of pregnancy and lactation on the model immunotoxic compound di-n-octyltin dichloride (DOTC) in rats using clinical and histopathological parameters. Female rats were exposed to 0, 3, 10, or 30 mg DOTC/kg feed during pregnancy and up to 20 (at weaning) or 56 days after delivery. Age-matched nonmated females were exposed during the same time periods. DOTC at the level of 10 and 30 mg/kg decreased thymus weight and affected thymus morphology in the lactating rats. In addition, DOTC decreased the numbers of neutrophils in the lactating rats. These effects were no longer apparent at day 56 despite continuous exposure to DOTC. This explorative study indicates that the innate and adaptive immune system may be especially sensitive to immunotoxicants during pregnancy and lactation.
The immune system is a highly dynamic system that is closely intertwined and influenced by the neural and endocrine system. Generally, immunotoxicity to xenobiotics and immune efficacy of drugs are investigated in young-adult animals with a mature and balanced immune system. However, a characteristic susceptibility of the immune system during embryonic and postnatal development to xenobiotic-induced insults is at present widely recognized and addressed as developmental immunotoxicology (Holladay 1999; Van Loveren and Piersma 2004; Ladics et al. 2005; Luebke et al. 2006; Dietert and Holsapple 2007). This increased sensitivity is due to the complex and rapid development of the immune system during the perinatal period. In toxicology, immune development has been characterized on the basis of critical windows, each with a specific vulnerability. The increased sensitivity during early development is reflected in the much lower exposure levels needed to induce immune alterations compared with exposure levels producing overt effects in adults. Moreover, early exposure to xenobiotics appeared more likely to produce persistent immune effects, whereas effects following adult exposures tended to resolve over time (Luebke et al. 2006). Finally, the type of effects due to perinatal exposure may differ completely from those induced during adult life (Kuper et al. 2007).
The developing immune system is not the only period during which the immune system goes through a series of complex and rapid changes. Pregnancy and lactation are periods in adult life during which the immune system changes dramatically and in a unique way (McLean, Mosley, and Gibbs 1974; Forster, McLean, and Gibbs 1979; Kendall and Clarke 1994; Endo and Kanayama 1998). In brief, early pregnancy in rats was associated with an increase in cell density in the thymus and spleen, whereas late pregnancy and lactation were associated with a decrease in thymus weight and cellularity, although the thymus medulla was expanded. Lactation delayed the return of the thymus to the prepregnancy state (Kendall and Clarke 1994). Hormonal and neural changes can partly explain these changes. Progesterone, prolactin, and estradiol levels in plasma fluctuate in early, mid, and late pregnancy (Fernandez-Twinn et al. 2003; Brusco et al. 2008). Nerve growth factor, nearly absent in the medulla of the adult thymus, was high in the medulla of the thymus at late pregnancy (Aloe et al. 1997). Thus, early and late pregnancy and lactation may be different windows, each potentially with their own sensitivity to immunotoxic xenobiotics and immune-active drugs. In addition, the changes in the thymus can be attributed to the adaptation of the maternal immune system to the semi-allogeneic fetus (half of the genes are supplied by the father). This is supported by the observation that changes are less distinct when mating occurs between genetically identical (inbred) animals.
A well-studied immunotoxic compound is di-n-octyltin dichloride (DOTC). DOTC induced thymus atrophy in rats (Seinen and Willems 1976; Grundel, Grupe and Gores 1984; Miller, Scott, and Foster 1984). The atrophy was caused by a decrease in the number of CD4/CD8 double-positive thymocytes in the cortex and led to a decrease in peripheral lymphocytes and a diminished T-lymphocyte–dependent immunity in rats (Pieters et al. 1992). Exposure during the developmental phase of the lymphoid system, more specifically postnatally, induced the most severe effects (Seinen, Vos, and Van Krieken 1977; Smialowicz et al. 1988).
In the present study, female rats were exposed via their feed to DOTC during mating, pregnancy, and lactation; additional groups of rats were exposed until day 56 after delivery. Subsequently, the thymus, spleen, and peripheral blood were examined at day 20 (at the end of the lactation period) and day 56 after delivery. The DOTC-induced effects in these females were compared with those in similarly exposed, nonpregnant and nonlactating females.
Exploration of Increased Sensitivity of Pregnant/Lactating Rats for the Immunotoxicant DOTC
Test material
DOTC (C16H34Cl2Sn, CAS no. 3542-36-7; batch number 1002585, purity 95%) was purchased from ABCR GmbH&Co KG (Karlsruhe, Germany). The appropriate amount of test substance was mixed with the feed and stored in a freezer <–18°C.
Animals and maintenance
The welfare of the animals was maintained in accordance with the general principles of the European Communities (Directive 86/609/EEC) and the Netherlands legislation (the Experiments on Animals Act 1997), governing the use of animals in toxicity experiments. Parental F0-rats, Wistar outbred ([Crl:WI] WU BR), were obtained from Charles River Deutschland (Sulzfeld, Germany). At the commencement of the treatment period, the F0 rats were approximately 9 to 10 weeks old. Housing conditions were conventional, in macrolon cages with stainless-steel grid covers and wood shavings as bedding material, in a controlled environment (temperature 20–24°C, humidity 40–70%, 12-h light-dark cycle, about 10 air changes per hour). In addition, paper material was provided for nesting material. The animals were fed a commercial rodent diet (RM3 diet; SD Special Diets Services, Witham, England). Feed and water were available at libitum.
Experimental design
The study comprised two main groups:
Pregnant and lactating females: Groups of female animals were fed the test or control diets starting 2 weeks prior to mating and throughout mating, gestation, and lactation until weaning of the F1 animals. The dams were sacrificed at weaning (day 20 after delivery). Additional groups were exposed until day 56 after delivery.
Nonpregnant and nonlactating females: A parallel group of nonmated female animals (same age as the pregnant/lactating groups) were fed the test or control diets.
The animals received DOTC at concentrations of 0, 3, 10, or 30 mg DOTC/kg RM3 diet. The body weight of each animal was recorded at initiation and at least once per week thereafter. In addition, the animals were weighed on their scheduled necropsy date to calculate the correct organ to body weight ratios. Feed consumption was measured per cage weekly. At necropsy, thymus and spleen were weighed and collected for histopathological evaluation. Because of the explorative nature of the study, the results were analyzed only with a Student’s t test.
Hematology
Hematology was conducted at necropsy. Blood was sampled from the abdominal aorta of the rats while under CO2/O2 anesthesia. Hemoglobin, packed cell volume, red blood cells, reticulocytes, total and differential white blood cells, and thrombocytes were measured with the Advia 120 Haematology Analyser. Mean corpuscular volume, mean corpuscular hemoglobin, and mean corpuscular hemoglobin concentration were calculated from hemoglobin, packed cell volume, and red blood cells. Prothrombin time was measured using the Normotest for EDTA blood (Nyegaard and Co., A/S, Norway).
Histology
Neutral, phosphate-buffered 4% (v/v) formaldehyde was used to preserve the spleen and one lobe of the thymus. These tissues were embedded in paraffin wax and sectioned at 5 µm. Cross sections of the spleen and longitudinal sections through one lobe of the thymus were stained with hematoxylin and eosin and examined microscopically.
Changes Related to Pregnancy/Lactation
Changes in peripheral blood composition
In lactating females, red blood cell counts and hemoglobin were decreased when compared with the nonmated control females (Table 1). The number of reticulocytes and the mean corpuscular volume were increased. The total number of white blood cells was not significantly altered at the end of the lactation period, but the ratio between the different cell types was distinctly altered. The absolute number and percentage of neutrophils were increased, and the percentage of monocytes was increased as well, whereas the percentages of lymphocytes and eosinophils were decreased. These data are in line with previously described hematology during pregnancy (Liberati, Sansone, and Feuston 2004; Honda et al. 2008).
Hematology of pregnant/lactating Wistar rats at day 20 after delivery, compared with age-matched nonmated controls.
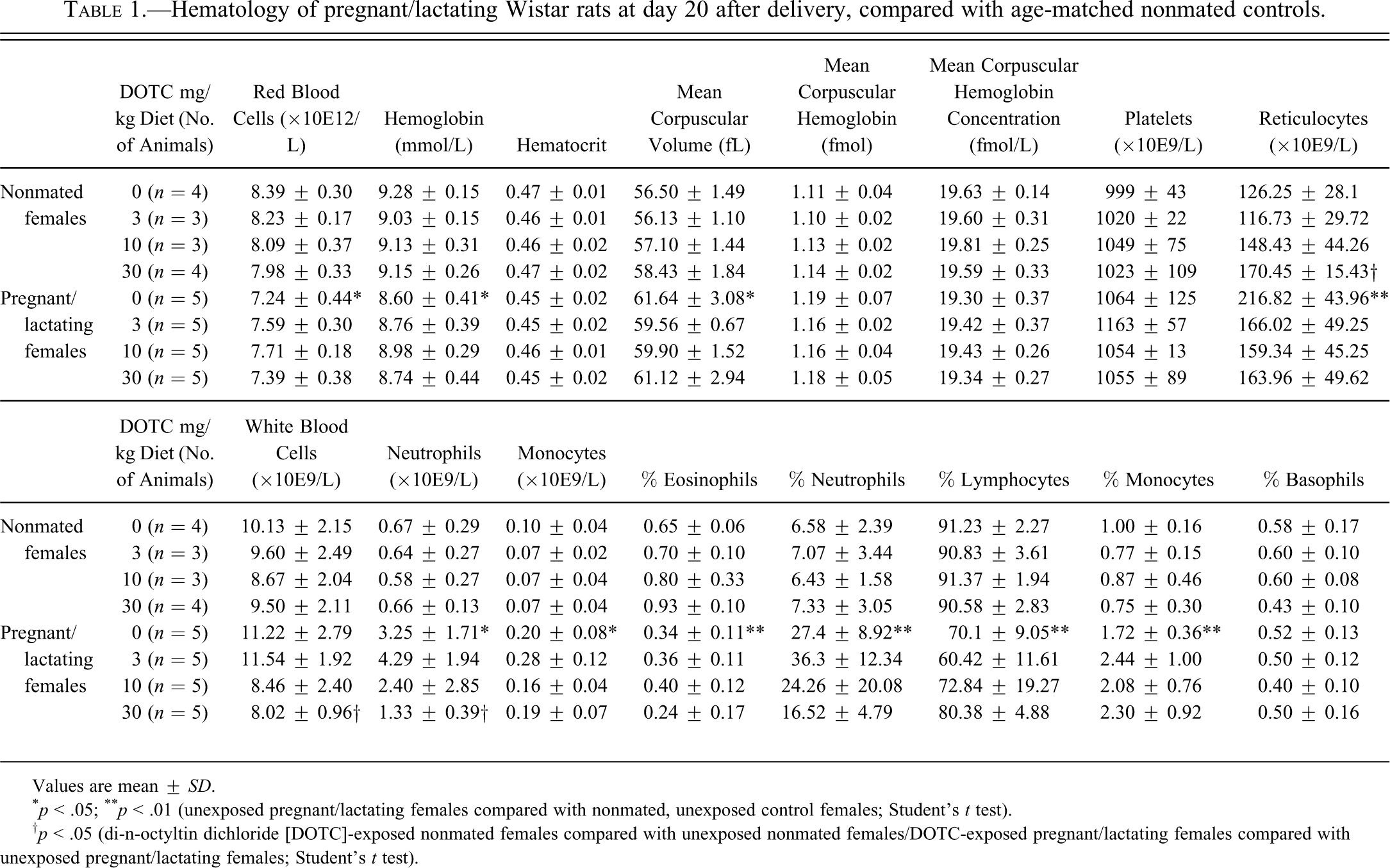
Values are mean ± SD.
* p < .05; ** p < .01 (unexposed pregnant/lactating females compared with nonmated, unexposed control females; Student’s t test).
† p < .05 (di-n-octyltin dichloride [DOTC]-exposed nonmated females compared with unexposed nonmated females/DOTC-exposed pregnant/lactating females compared with unexposed pregnant/lactating females; Student’s t test).
Changes in the thymus and spleen
At the end of the lactation period, the weight of the thymus was significantly decreased when compared with the thymic weight of the nonmated control females (Table 2). The decreased thymic weight correlated with a reduction in overall size of the thymus and a lower cellularity in the cortex. The medulla appeared to be expanded, relative to the cortex (Figure 1A,B). Similar changes in the thymus have been described in rats during their late pregnancy by Kendall and Clarke (1994).
Relative thymus and spleen weight of pregnant/lactating Wistar rats at day 20 and 56 after delivery, compared with age-matched nonmated controls.
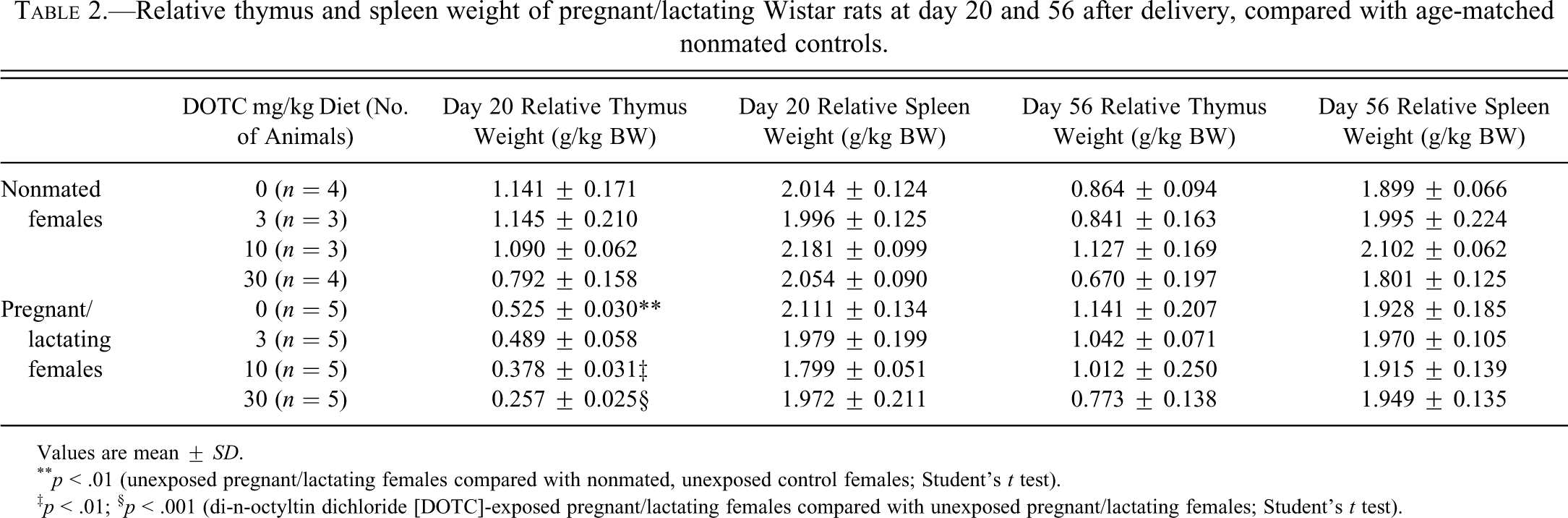
Values are mean ± SD.
** p < .01 (unexposed pregnant/lactating females compared with nonmated, unexposed control females; Student’s t test).
‡ p < .01; § p < .001 (di-n-octyltin dichloride [DOTC]-exposed pregnant/lactating females compared with unexposed pregnant/lactating females; Student’s t test).
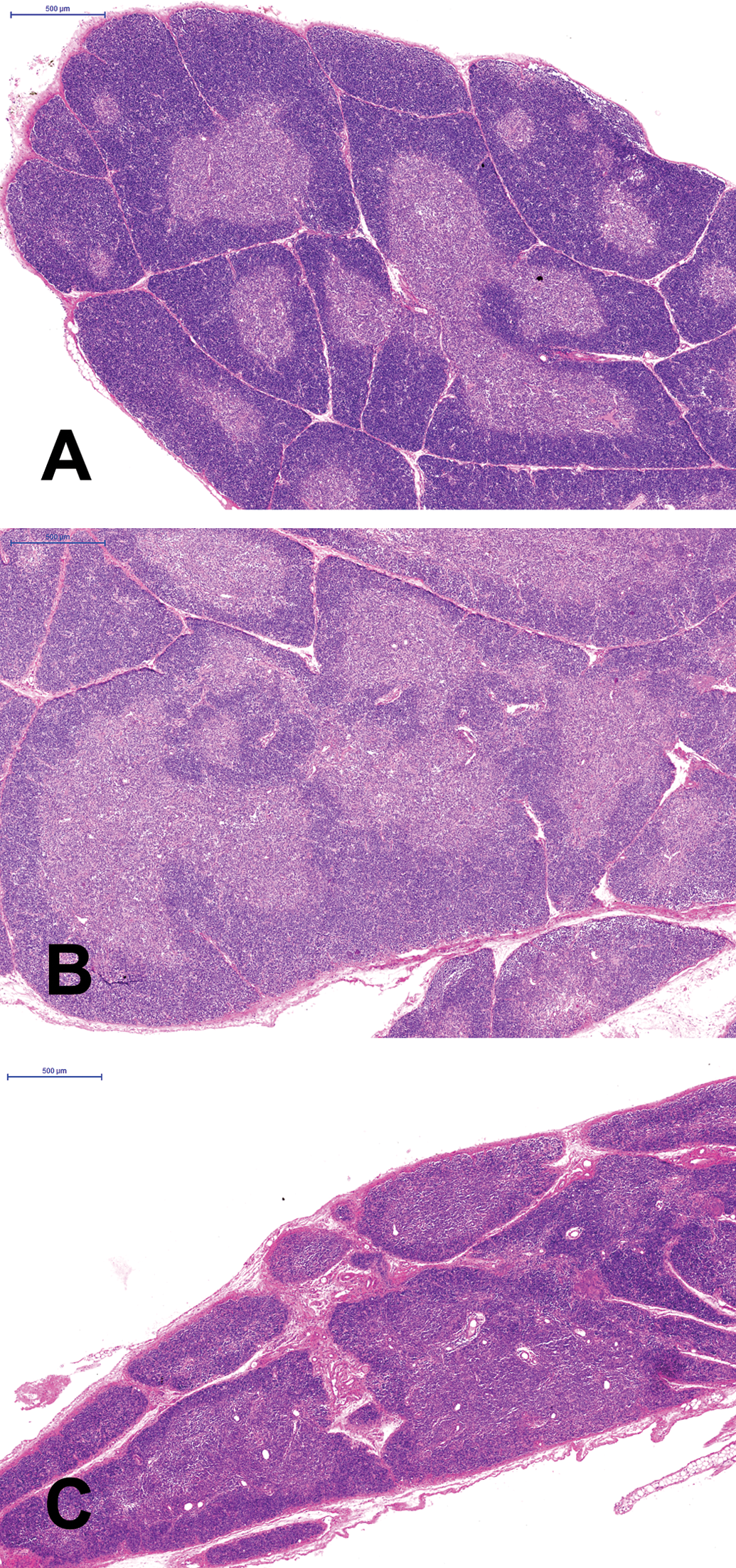
Thymus of a nonmated (nonpregnant/lactating) control Wistar rat (A), a pregnant/lactating Wistar rat (B), and a pregnant/lactating rat exposed to 30 mg/kg di-n-octyltin dichloride
At the end of the lactation period, the weight of the spleen was not affected. Histopathological analysis showed that the spleen of the lactating control females contained far less hemosiderin pigment than their age-matched nonlactating controls (Figure 2A,B). Although the numbers of animals were small, the lactating females appeared to have less pronounced periarteriolar lymphocyte sheets than the nonmated controls.
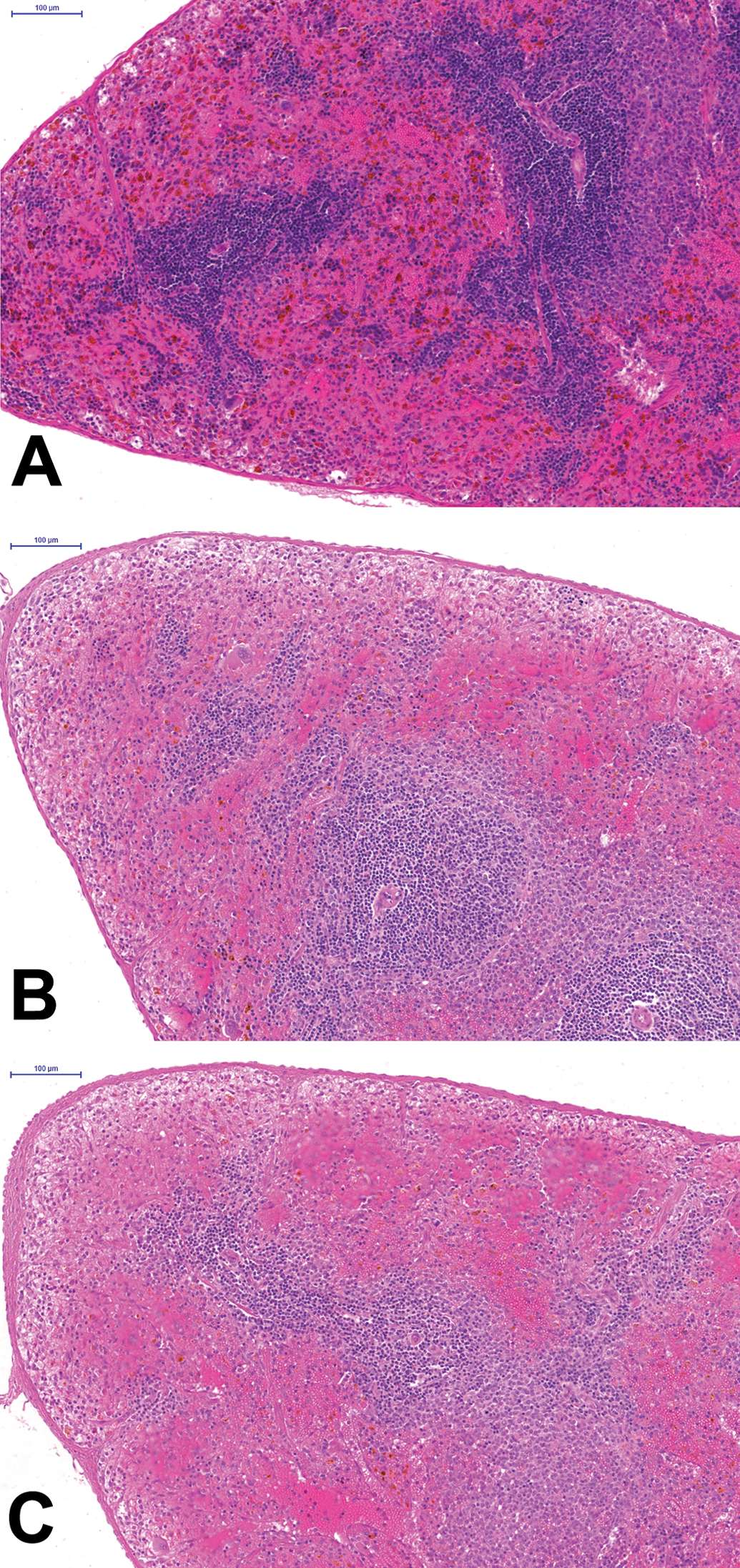
The spleen of the nonpregnant control (A) contains much more hemosiderin in the red pulp than the pregnant/lactating rat (B). In addition, the white pulp appears smaller in size. The small size of the white pulp appears even more prominent in the di-n-octyltin dichloride–exposed animal, but the interindividual variability was considerable. Hematoxylin and eosin stain.
DOTC Affected the Lactating Females but Not the Nonmated Females
DOTC induced a decrease in neutrophil counts in lactating females
The absolute number of white blood cells was lower in lactating females fed 30 mg/kg DOTC (statistically significant) and 10 mg/kg DOTC than in the lactating control females (Table 1). In line with this observation, a decrease in the numbers of circulating neutrophils was observed. DOTC had no effect on the white blood cell counts in nonlactating females (Table 1).
DOTC decreased thymus weight, cortex size, and cellularity in lactating females
The weight of the thymus was significantly and dose-dependently reduced in the lactating females at 10 and 30 mg/kg DOTC compared with the lactating control females (Table 2). Likewise, the thymic cortex of the lactating females was reduced in size (Figure 1C), at 10 and 30 mg/kg DOTC in a dose-dependent way in all animals. Spleen weight and morphology (Figure 1B) were not affected by DOTC in the diet (Figure 2C).
Are Pregnancy and Lactation Indeed Periods of Increased Sensitivity to Immunotoxic Agents?
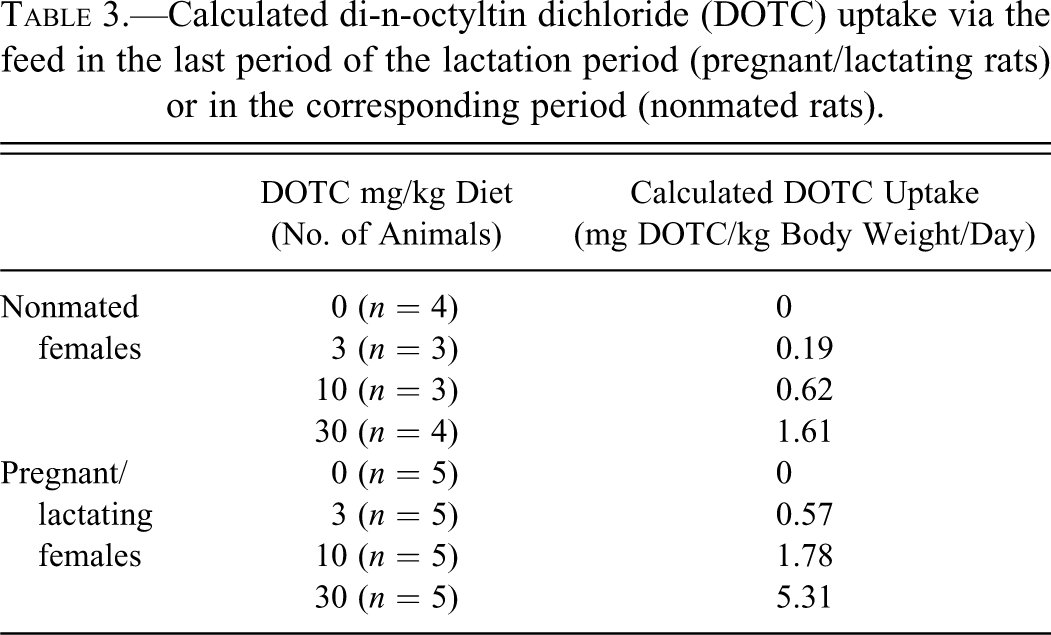
The results of the DOTC study showed a distinct increased sensitivity of lactating female rats for the immunotoxicant, when compared with age-matched, nonmated (nonpregnant/nonlactating) females. A large litter size can prolong or increase pregnancy-related effects during lactation (Kendall and Clarke 1994), but litter size was not affected by DOTC. However, there is an aspect to be considered when the animals are exposed to the test substance via the diet. In the last 2 weeks of the lactation period (between days 7 and 21 postdelivery), feed consumption increases considerably, and the lactating females are thus exposed to an increased amount of the test substance in the diet. In the present study, DOTC uptake by the lactating rats of the mid-dose group (10 mg DOTC/kg diet) was comparable to that of the high-dose nonmated rats, 1.78 and 1.61 mg DOTC/kg body weight/day, respectively (Table 3). Nevertheless, DOTC induced thymic changes in the lactating rats of the mid-dose group but not in the high-dose nonmated females. Thus, the increased feed consumption did not explain entirely the increased sensitivity of the lactating rats for the immunotoxic actions of DOTC. Moreover, DOTC nullified the observed increase of circulating neutrophils in the lactating females, an effect that may be unique for pregnancy and lactation periods because it was not observed in the nonmated females and to our knowledge has not been reported previously. Both observations suggest that DOTC exposure has a more pronounced and/or specific impact on the acquired and innate immune system of pregnant/lactating females than on the nonmated animals. Altered sensitivity to immunotoxicants or immune-modulating drugs can be expected in humans as well. The immune system of humans changes during pregnancy, reflected by the frequent improvement of symptoms of rheumatoid arthritis, only to flare up postpartum. Although the mechanisms of this phenomenon are still poorly understood, it is hypothesized that it is due to the immunologic changes occurring during pregnancy (Amin et al. 2011).
Calculated di-n-octyltin dichloride (DOTC) uptake via the feed in the last period of the lactation period (pregnant/lactating rats) or in the corresponding period (nonmated rats).
About 2 months after delivery, the immune parameters tested were fully recovered to the prepregnancy state and comparable with those in the nonmated females. Feed consumption of the females, which had been pregnant and lactating, returned to normal. Despite continuous exposure, DOTC-induced effects were not observed in these females. However, it remains to be elucidated if immune functioning had also fully recovered.
In the study with DOTC, the effects were measured at the end of the lactation period, when the immune system is expected to return to the prepregnancy state. It is possible that the effects of DOTC in the mated females would have been more profound when they were measured during pregnancy. It is concluded that the explorative study with DOTC in pregnant/lactating female rats has provided indications that pregnancy and lactation are sensitive periods in the adult life, which warrants further investigation.
The authors declared no potential conflicts of interests with respect to the authorship and/or publication of this article. The authors received no financial support for the research and/or authorship of this article.