
Abstract
The C57BL/6 mouse strain (or derivation of this strain) is used as a background for many transgenic mouse models. This strain has a relatively low susceptibility to chemically induced hepatocarcinogenesis compared with other commonly used experimental mouse strains. In the present study, the authors treated C57BL/6 mice with 25, 50, and 75 mg/kg of diethylnitrosamine (DEN) for 4 or 8 weeks by intraperitoneal injection to investigate the dose-response pattern of preneoplastic and neoplastic lesion formation in the liver. DEN induced preneoplastic lesions and cytokeratin 8/18–positive foci in a dose-dependent manner. In the 75 mg/kg for 8 weeks treatment group, hepatocellular adenoma, cholangioma and hemangioma, and cytokeratin 19–positive foci were also induced, but a significant decrease in body weight was observed. The suitable DEN treatment range for this strain was concluded to be from 75 mg/kg for 4 weeks (total amount = 300 mg/kg) to 50 mg/kg for 8 weeks (total amount = 400 mg/kg). These results should prove useful for future studies investigating hepatocarcinogenesis in both the background C57BL/6 strain and other transgenic mouse models derived from it.
Keywords
Introduction
Mice are used extensively in carcinogenicity bioassays and mechanistic toxicology studies. However, striking interstrain differences exist in laboratory mice with respect to liver tumor development. For example, the C3H mouse strain is highly susceptible to spontaneously and chemically induced liver tumors, whereas the C57BL/6 mouse strain is comparatively resistant to liver tumor development (Drinkwater and Ginsler 1986; Maronpot 2009; Harada et al. 1999). With the increased interest in the use of genetically engineered mice of the C57BL/6 background to study the mechanisms of hepatic carcinogenesis, the lower liver tumor susceptibility of this strain of mice complicates its use in these mechanistic studies (Harvey et al. 1993). Diethylnitrosamine (DEN) has been used extensively in the past as a carcinogen and/or lesion initiator in animal model systems of carcinogenesis. DEN itself induces hepatic neoplastic and preneoplastic lesion in mice (Goldfarb et al. 1983). To induce hepatic tumors by DEN, infant mice are often used as the developing liver is susceptible especially to genotoxic chemical carcinogens (Vesselinovitch et al. 1984; Fu et al. 2000). The distinct initiation-promotion steps of the multistage hepatocarcinogenesis model are well recognized. Again, while DEN is extensively used as an initiator, a certain “time of initiation” effect has been reported on the ability of promoters to effectively facilitate the clonal expansion of preneoplastic cells (Lee 2000). For example, male B6C3F1 mice treated at 30 days of age with a single dose of DEN followed by chronic exposure to phenobarbital, a strong hepatic tumor promoter, exhibited an enhancement of hepatic tumor incidence and number compared with DEN-only–treated mice (Diwan et al. 1984; Ward et al. 1983). However, the efficacy of phenobarbital as a tumor promoter appears weak or lacking in the C57BL/6 strain (Bursch et al. 2005; Diwan et al. 1986; Lee et al. 1989). Furthermore, the initiation of B6C3F1 mice at day 15 of age with a single dose of DEN followed by chronic treatment with phenobarbital resulted in an inhibition of hepatic tumor formation in other studies (Klaunig et al. 1987, 1988; Lee 2000). Such inhibitory effects of phenobarbital on hepatocarcinogenesis have also been recognized in C57BL/6 mice (Weghorst and Klaunig 1989). These data suggest that both the timing and dosage of DEN treatment are important considerations for the two-stage hepatocarcinogenesis model.
For the evaluation of mechanisms of hepatocarcinogenicity, in vivo short-term or medium-term liver bioassay systems are frequently used. In these short- or medium-term bioassay systems, preneoplastic lesions are evaluated as endpoint markers for hepatocarcinogenicity. In mouse hepatocarcinogenesis, altered hepatocellular foci (AHF) have been used as markers of preneoplastic lesions, since no reliable pathological marker has been reported. Recently, Kakehashi et al. (2010) reported that immunohistochemically stained cytokeratin 8/18 (CK8/18)–positive foci might be a useful candidate as marker of preneoplastic lesions in B6C3F1 and C57BL/6J mice. Their report said that the CK8/18-positive reaction was recognized not only in all basophilic AHF but also in all hepatocellular tumors (Kakehashi et al. 2010). Furthermore, cytokeratin 19 (CK19) has been described as a prognostic marker of early neoplastic lesions in the rat liver carcinogenesis (Andersen et al. 2010). Therefore, we evaluated whether CK8/18- and CK19-positive foci would be of utility in the C57BL/6 mouse.
Few studies have examined the nature of the dose-response relationship between DEN and the formation of preneoplastic/neoplastic lesions in postweaned treated C57BL/6 mice because of this strain’s low susceptibility to liver tumor development. However, as C57BL/6 mice are the background strain for many kinds of transgenic mice, it is important for future studies to establish a reproducible model for the induction of liver tumors and preneoplastic lesions in this strain. The observed different response of lesions induced in the initiated infant mouse to tumor promoters compared with that seen in the adult mouse points to a need to further develop and characterize a model for hepatic lesion induction in the postweaned C57BL/6 mouse (Klaunig et al. 1987, 1988; Weghorst and Klaunig 1989; Lee 2000). In the present study, we treated male C57BL/6 mice with several protocols of intraperitoneal (i.p.) injections of DEN (25, 50, and 75 mg/kg for 4 or 8 weeks) to investigate preneoplastic and neoplastic lesion formation in the liver.
Materials and Methods
Chemical
DEN was purchased from Sigma-Aldrich (St. Louis, MO).
Animals
A total of eighty 21-day-old male C57BL/6 mice were purchased from Charles River (Wilmington, MA) and housed in polycarbonate cages in an animal facility certified by the Association for Assessment and Accreditation of Laboratory Animal Care. Maintained in accordance with the National Institutes of Health (NIH) Guide for the Care and Use of Laboratory Animals, all of the mice were fed NIH-07 diet rodent chow and received water ad libitum. The animals were allowed to acclimate for 1 week prior to the experiments.
Experimental Design
After acclimation, the 4-week-old mice were randomly divided into eight experimental groups. In the 4-week treatment group, mice were treated with DEN dissolved in saline by i.p. injection at the dosages of 0, 25, 50, and 75 mg/kg on experimental days 0, 7, 14, and 21 (groups 1–4). In the 8-week treatment group, i.p. injections were given as above on experimental days 0, 7, 14, 21, 28, 35, 42, and 49 (groups 5–8). At experimental day 233, mice were killed by CO2 asphyxiation and exsanguinations, weighed, and necropsied. The livers were removed in toto, weighed, and examined for the presence of grossly visible lesions. They were then separated by lobe and cut into 1- to 2-mm slices. One from each lobe was fixed in formalin for 48 to 72 hours and then embedded in paraffin to give a total of three paraffin blocks per animal. Serial sections from each block were stained with hematoxylin and eosin (H&E), CK8/18, and CK19 immunohistochemistry. Hepatic preneoplastic and neoplastic lesions were classified according to the Pathology of the Mouse (Harada et al. 1999).
Quantitative Analysis of Altered Hepatocellular Foci
Quantitative analysis of AHF was accomplished using a two-dimensional evaluation. Whole H&E staining sections were scanned by ScanScope CS (Aperio Technologies, Vista, CA). Images were then viewed with Aperio’s image viewer software (ImageScope), on which the quantitative analyses were performed. The number and volume of each foci (eosinophilic, clear, and basophilic) and total volume of liver sections were quantified. Because large hepatic tumors were recognized in two animals (one animal in the 25 mg/kg for 8 weeks treatment group and the other in the 75 mg/kg for 8 weeks group), these areas were not included in the total volume of liver sections of these animals. The multiplicity of AHF is reported as numbers per centimeter squared and the volume of foci as the percentage of total liver volume (%).
Immunohistochemistry for CK8/18 and CK19
For staining of formalin-fixed paraffin-embedded tissue sections, polyclonal guinea pig CK8/18 antibody (dilution 1:600; LifeSpan BioSciences, Seattle, WA) and polyclonal rabbit CK19 (dilution 1:50; Abcam, Cambridge, MA) antibodies were used. After deparaffinization, microwave antigen retrieval was performed in citrate buffer (pH 6.0) at 95°C for 30 minutes, and then sections were incubated with 3% hydrogen peroxide for 15 minutes to block endogenous peroxidase. For blocking of nonspecific antibody binding, the sections for CK8/18 were incubated in the Avidin/Biotin Blocking Kit (Vector Laboratories, Burlingame, CA) and then in blocking buffer, phosphate-buffered saline (PBS) with 1% bovine serum albumin, 1.5% normal goat serum, and 0.02% NaN3. For CK19, PBS with 1% skim milk and 2.5% normal horse serum was used. Sections were incubated overnight at 4°C with primary antibodies diluted in blocking buffer. The following day, sections for CK8/18 were washed in PBS, incubated with the appropriate secondary antibody (biotinylated anti-guinea pig supplied with the Vectastain ABC kit), and immunoreactivity revealed by the avidin-biotin method (Vectastain ABC Kit [guinea pig IgG]; Vector Laboratories). For CK19, incubation was with an ImmPRESS anti-rabbit Ig (peroxidase) kit (Vector Laboratories). Antigen visualization was accomplished with 3,3′-diaminobenzidine tetrahydrochloride. The numbers and areas of foci and total areas of liver sections were measured by the same way as detailed above.
Statistical Analysis
Statistical analysis was performed with Statistical Analysis System (SAS) version 9.1.3 (SAS Institute Inc., Cary, NC). For the incidence of hepatic lesions, Fisher’s exact test was used. For the remainder of the data, testing for homogeneity was conducted using Bartlett’s analysis. For data found to be homogeneous, Dunnett’s multiple analysis was performed. If the data were not homogeneous, Steel’s multiple analysis was conducted. The significance of differences was determined based on probability levels of 1% and 5%. The data presented are means or means ± standard deviation.
Results
General Observation, Body and Liver Weight
The final body weight and absolute and relative liver weights are shown in Table 1 . During the experimental period, one mouse in the 4 weeks 50 mg/kg treatment group (group 3), one mouse in the 4 weeks 75 mg/kg treatment group (group 4), and two mice in the 8 weeks 50 mg/kg treatment group (group 7) died. The causes of death could not be determined at necropsy. One mouse each in the 4 weeks 25 mg/kg (group 2), 4 weeks 75 mg/kg (group 4), 8 weeks 50 mg/kg (group 7), and 8 weeks 75 mg/kg treatment (group 8) groups were subjected to moribund sacrifice when emaciation was observed. Final body weights were significantly decreased in the groups 4, 7, and 8 as compared with the control group. Measurement of organ weight revealed that the relative liver weight was significantly increased in group 8 as compared with the control group, whereas the absolute liver weight in group 4 was significantly decreased. Decreased absolute liver weight in this group was attributed to decreased body weight, as there was no difference in relative liver weight.
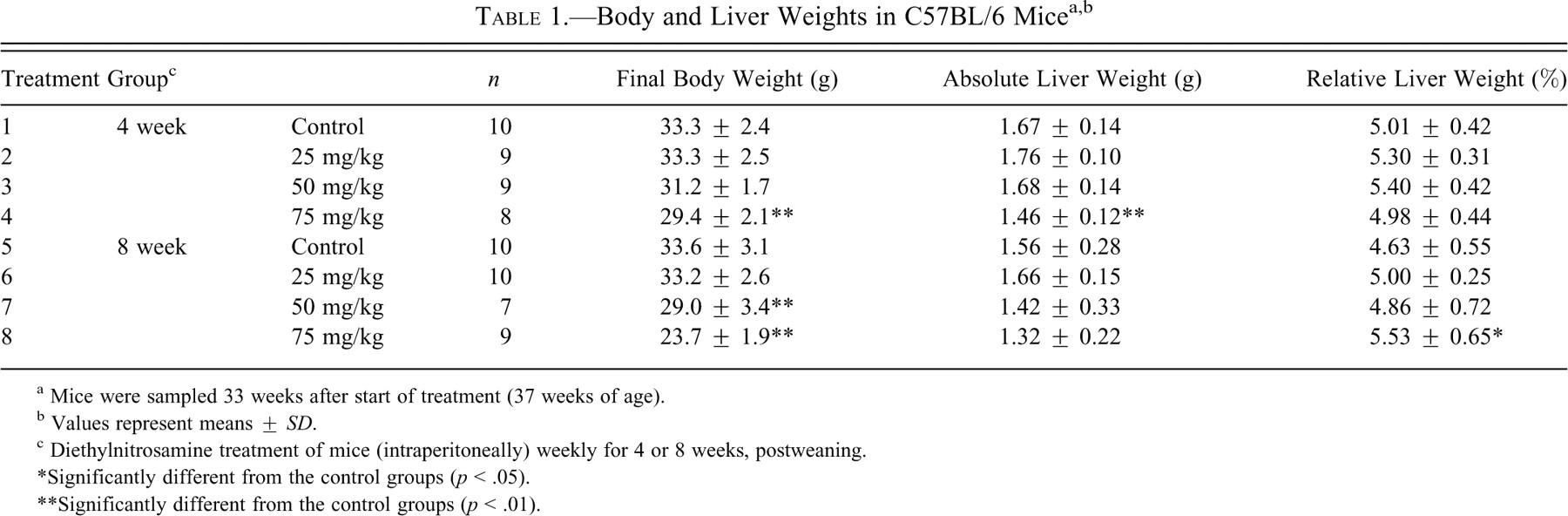
a Mice were sampled 33 weeks after start of treatment (37 weeks of age).
b Values represent means ± SD.
c Diethylnitrosamine treatment of mice (intraperitoneally) weekly for 4 or 8 weeks, postweaning.
*Significantly different from the control groups (p < .05).
**Significantly different from the control groups (p < .01).
Histopathological Examination
Table 2 summarizes the results of the histopathology examination. Hepatocellular adenomas, cholangiomas, and hemangiomas were observed in group 8 (Table 2; Figure 1 ). A hepatocellular adenoma was also identified in group 6 (Table 2). In all DEN treatment groups, eosinophilic, clear, and basophilic AHF were identified (Figures 1–3). The incidences of clear and basophilic AHF increased in a dose-dependent manner and were statistically significant as compared with control values for groups 3, 4, 6, 7, and 8 (Table 2). Eosinophilic AHF were also positively correlated to dose, but only those identified in group 8 were statistically significant. In addition, biliary cysts were identified in most of the group 8 animals (Table 2; Figure 1).
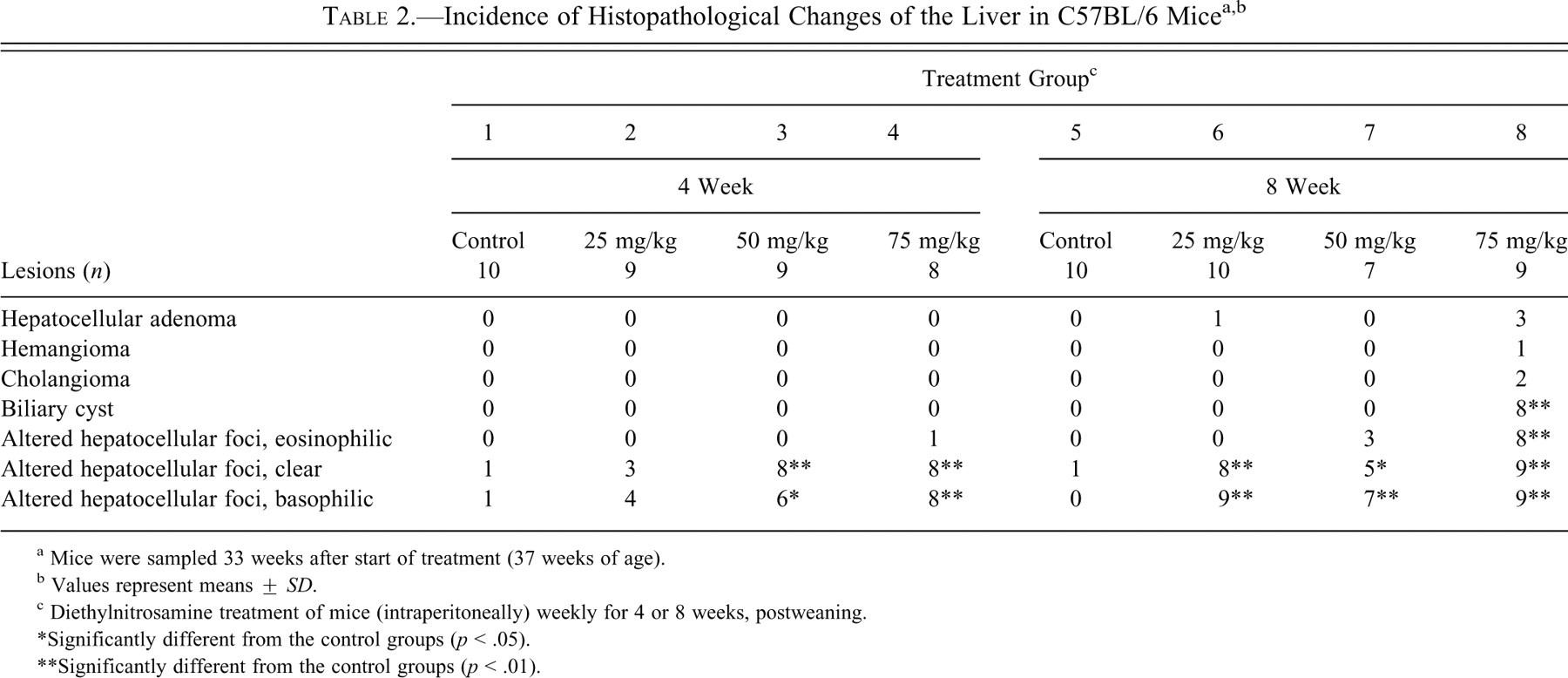
a Mice were sampled 33 weeks after start of treatment (37 weeks of age).
b Values represent means ± SD.
c Diethylnitrosamine treatment of mice (intraperitoneally) weekly for 4 or 8 weeks, postweaning.
*Significantly different from the control groups (p < .05).
**Significantly different from the control groups (p < .01).
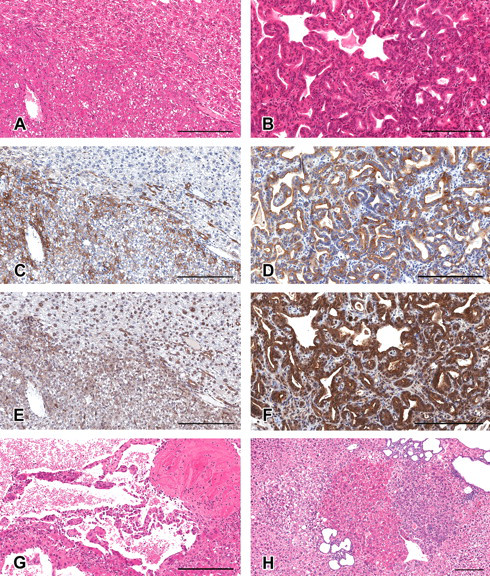
Formation of altered hepatocellular foci (AHF) and tumors in the liver of C57BL/6 mice treated with diethylnitrosamine. (A–G) The lesions of a hepatocellular adenoma (A, C, and E), a cholangioma (B, D, and F), and a hemangioma (G). (H) Typical basophilic, clear, and eosinophilic AHF, along with a biliary cyst. (A, B, G, and H) Hematoxylin and eosin stained. (C, D) Immunohistochemically stained for cytokeratin 8/18. (E, F) Immunohistochemically stained for cytokeratin 19. (A, C, and E) and (B, D, and F) are serial sections. All figures are for livers of mice treated at the dosage of 75 mg/kg for 8 weeks. Scale bars are 200 µm.
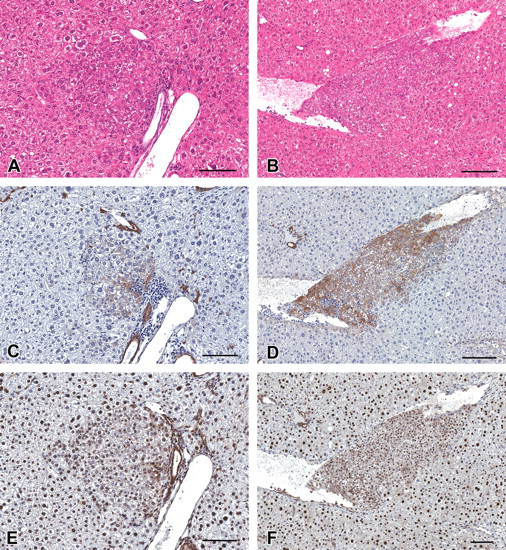
Histology and immunohistochemistry of basophilic foci in serial sections in the liver of C57BL/6 mice treated with diethylnitrosamine. (A–F) Basophilic AHF (A, C, and E) and invasive basophilic AHF (B, D, and F). (A, B) Hematoxylin and eosin stained. (C, D) Immunohistochemically stained for cytokeratin 8/18. (E, F) Immunohistochemically stained for cytokeratin 19. An immunohistochemically strongly positive reaction for CK8/18 was recognized in invasive basophilic AHF (D). Scale bars are 200 µm.
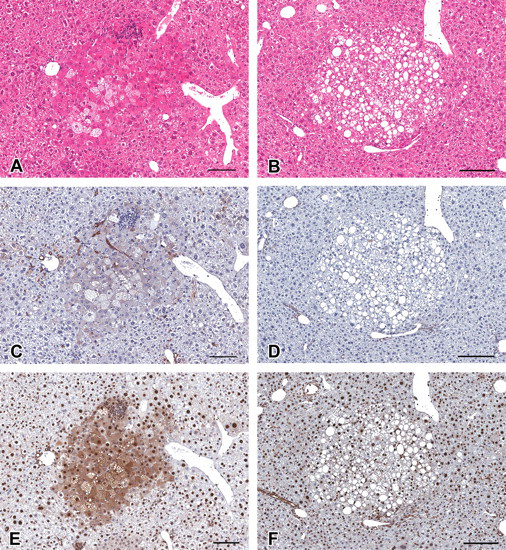
Immunohistochemistry for CK8/18 and CK19, and hematoxylin and eosin (H&E) staining in serial sections in the liver of C57BL/6 mice treated with diethylnitrosamine. (A–F) Eosinophilic altered hepatocellular foci (AHF; A, C, and E), and clear AHF (B, D, and F). (A, B) H&E stained. (C, D) Immunohistochemically stained for cytokeratin 8/18. (E, F) Immunohistochemically stained for cytokeratin 19. Note the strongly positive reaction for CK19 in eosinophilic AHF (E). Clear AHF were not positive for cytokeratin 8/18 and 19. Scale bars are 200 µm.
Quantitative Analysis of Multiplicity and Volume of the AHF in H&E Stained Sections
A quantitative morphometrical evaluation of AHF more clearly revealed that the number and volume of AHF increased in correlation with DEN dose levels (Table 3 ). In the 8-week treatment groups, the volume of total foci was significantly increased from the low-dose group compared with the control group, and basophilic AHF accounted for more than 50% of the total AHF. In the 4-week treatment groups, the volume of total foci was significantly higher in group 3, and more than half of the total AHF were basophilic. The numbers data followed a similar trend as the above reported AHF volume data.
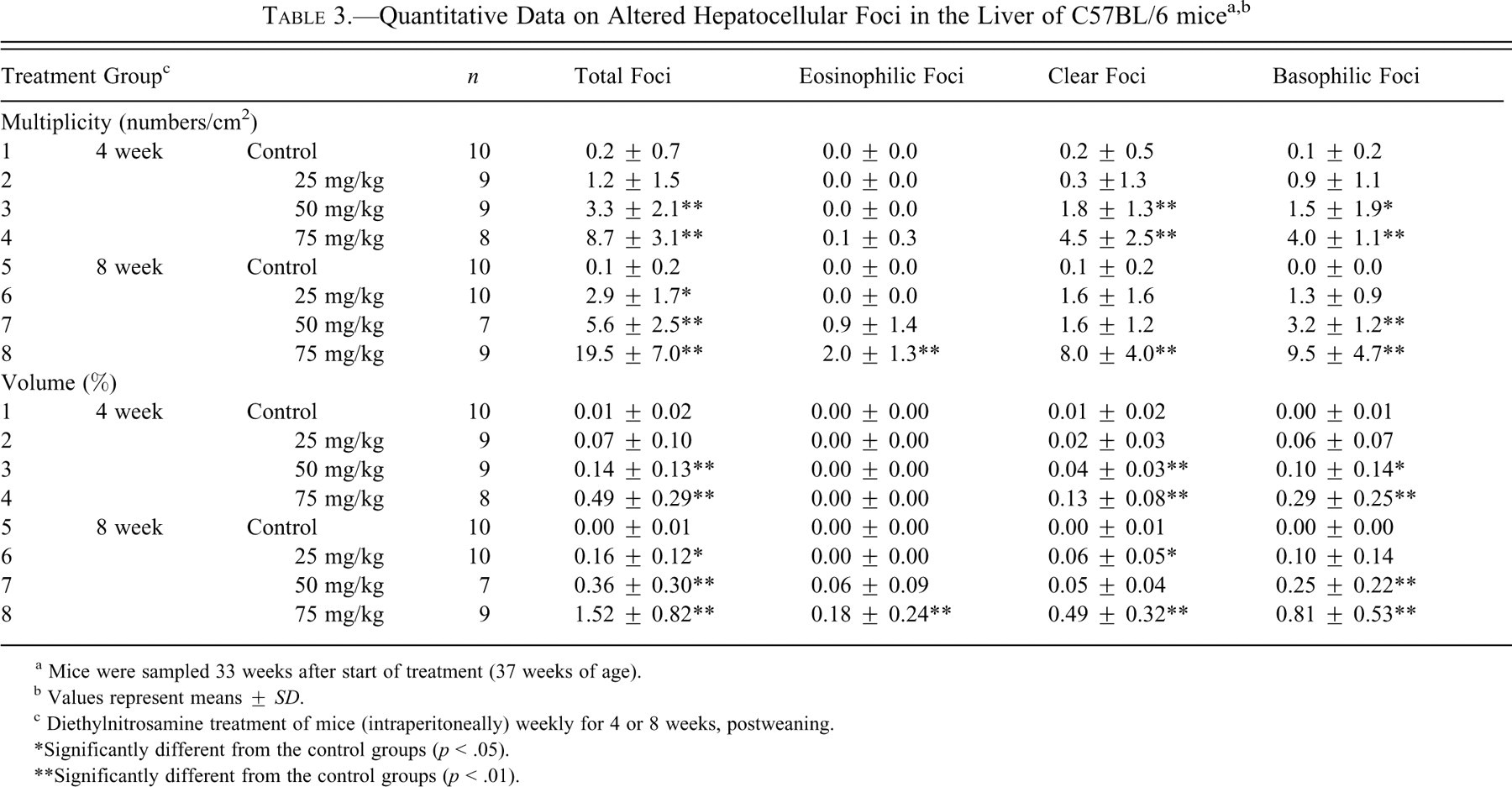
a Mice were sampled 33 weeks after start of treatment (37 weeks of age).
b Values represent means ± SD.
c Diethylnitrosamine treatment of mice (intraperitoneally) weekly for 4 or 8 weeks, postweaning.
*Significantly different from the control groups (p < .05).
**Significantly different from the control groups (p < .01).
Immunohistochemistry for CK8/18
Immunohistochemically positive reactions for CK8/18 were observed in bile ducts in all animals. As Kakehashi et al. (2010) reported, the CK8/18-positive reaction was recognized in basophilic AHF (Figure 2) and hepatocellular adenoma (Figure 1). In addition, its positive staining was observed in eosinophilic AHF (Figure 3) and cholangioma (Figure 1). The positive reaction for CK8/18 in both basophilic and eosinophilic AHF was significant but less prominent compared with its expression in the tumors. No clear AHF were stained by CK8/18 (Figure 3). Interestingly, basophilic AHF invasive to hepatic veins were strongly stained (Figure 2). In this study, however, CK8/18-negative basophilic AHF and hepatocellular adenoma were also observed (Figure 4 ). The area and number of CK8/18-positive foci in the livers of DEN-treated C57BL/6 mice increased in a dose-dependent manner, and statistical significance was observed at 50 mg/kg and greater (Table 4 ).
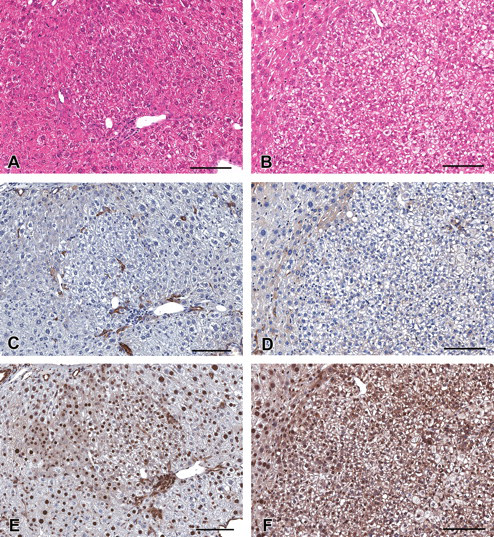
Histology and immunohistochemistry of cytokeratin 8/18 (CK8/18)–negative lesions in serial sections in the liver of C57BL/6 mice treated with diethylnitrosamine. (A–F) CK8/18-negative basophilic altered hepatocellular foci (A, C, and E) and a CK8/18-negative hepatocellular adenoma (B, D, and F). (A, B) Hematoxylin and eosin stained. (C, D) Immunohistochemically stained for cytokeratin 8/18. (E, F) Immunohistochemically stained for cytokeratin 19. Scale bars are 200 µm.

a Mice were sampled 33 weeks after start of treatment (37 weeks of age).
b Values represent means ± SD.
c Diethylnitrosamine treatment of mice (intraperitonally) weekly for 4 or 8 weeks, postweaning.
*Significantly different from the control groups (p < .05).
**Significantly different from the control groups (p < .01).
Immunohistochemistry for Cytokeratin 19
Immunohistochemically positive reactions for CK19 were observed in the bile duct in all animals and cholangiomas (Figure 1). The expression pattern of CK19 was similar to that for CK8/18, that is, positive reactions were observed in hepatocellular adenomas and basophilic and eosinophilic AHF but not in clear AHF (Figures 1–3). However, the numbers of CK19-positive foci were fewer than for CK8/18-positive foci (Table 4). Interestingly, some eosinophilic AHF were strongly stained (Figure 3). The expression of CK19 in the CK8/18-negative basophilic AHF and hepatocellular adenomas was very weak or not recognized (Figure 4). The area and number of CK19-positive foci in the livers of DEN-treated C57BL/6 mice were increased in a dose-dependent manner, but statistical significance was observed only in group 8 (Table 4).
Discussion
In the present study, a distinct dose-dependent response in preneoplastic liver lesion development following DEN treatment for 4 or 8 weeks was observed in 4-week-old C57BL/6 mice. DEN induced clear and basophilic AHF in the 25-mg/kg and above treatment groups. In addition, eosinophilic AHF were seen in the groups with 50 mg/kg after 8 weeks of DEN treatment and at 75 mg/kg after 4 weeks of DEN treatment. More than half of the identified AHF were basophilic type. CK8/18-positive foci were observed at 50 mg/kg and greater, and in mice treated with DEN at the dosage of 75 mg/kg for 8 weeks, hepatocellular adenomas, cholangiomas and hemangiomas, and CK19-positive foci were also observed. In addition, biliary cysts were further identified in most of the mice in this group.
In rats, several studies have investigated the relationship between DEN treatment and the multistage steps of the hepatocarcinogenesis process (Gray et al. 1991; Tsuda et al. 2003; Williams et al. 1993). In contrast, similar studies in the mouse have been limited (Vesselinovitch et al. 1984). In the present study, we demonstrated that DEN induced preneoplastic lesions in a dose-dependent manner. The mice treated with DEN at the dosage of 75 mg/kg for 8 weeks (total = 600 mg/kg) developed hepatocellular adenoma, cholangioma, and hemangioma during the 33-week duration of the experiment. Biliary cysts were also identified in most of the mice in this group. Both these biliary cysts and cholangiomas indicated that DEN affected not only hepatocyte proliferation but also bile duct pathology. The mean final body weight of animals in this group was significantly lower (29.5%) than in the control group, indicating that this dose exceeded the maximum tolerated dose. When mice were administered 75 mg/kg for 4 weeks (total = 300 mg/kg) or 50 mg/kg for 8 weeks (total = 400 mg/kg), basophilic and clear as well as eosinophilic AHF were identified. Both the mice in the 50 mg/kg for 4 weeks treatment group and the 25 mg/kg for 8 weeks treatment group received a total of 200 mg/kg, and no difference in AHF formation was present between the two (Student’s t-test, data not shown). Transgenic mice are becoming more frequently employed in the field of hepatocarcinogenesis research. Previous studies using DEN initiation in transgenic mice treated the animals at 15 days of age (Kang et al. 2007; Tamano et al. 1994). While treatment of preweaning, infant mice show an increased sensitivity in liver lesion to many hepatocarcinogens (Vesselinovitch et al. 1984), the lesions induced with the 15-day-old treatment are phenotypically and perhaps genotypically different than those seen following chronic treatment in the adult. Lee (2000) demonstrated that DEN treatment in the preweaning period induced only basophilic cell–type tumors and never eosinophilic and clear cell types in the B6C3F1 strain. Others (Klaunig et al. 1987, 1988; Fu et al. 2000) have shown an inhibitory response of lesion growth and formation in preneoplastic foci by tumor-promoting compounds such as phenobarbital in 15-day-old DEN-initiated mice. These results suggested that the initiation process (DEN treatment) must occur in the postweaning period for the initiation promotion with classical tumor promoters to occur in mice. It is well known that C57BL/6 mice are less sensitive to DEN-induced hepatocarcinogenesis compared with the C3H strain (Bursch et al. 2005); previous studies have shown it is difficult to detect the effects of promoters in a traditional initiation-promotion model in the C57BL/6 strain. Our results demonstrate that DEN does induce hepatic preneoplastic lesions in C57BL/6 mice in a dose-dependent manner, when DEN treatment dosages are of sufficient concentration and duration.
CK8 and CK18 are known to be some of the intermediate filaments and associated with the plasma membrane (Ku et al. 2007). CK8/18 are expressed on the cell membrane of adult hepatocytes and hepatobiliary ductal cells (Ku et al. 2007). It has been reported that CK8/18 is diffusely expressed in hepatocellular carcinomas of human liver (Stroescu et al. 2006). Recently, Kakehashi et al. (2010) demonstrated that CK8/18 were positively expressed in all basophilic AHF and hepatocellular tumors in the livers of C57BL/6J mice treated with DEN. In addition, CK8/18-positive foci were also induced in the liver of RasH2 mice treated with DEN and fenofibrate (Kawai et al. 2008). In our study, we could demonstrate a CK8/18-positive reaction not only in basophilic AHF and hepatocellular adenoma but also in eosinophilic AHF (Figures 1 and 2). No clear AHF were stained by CK8/18. Positivity for CK8/18 was slight in AHF but was strong in the tumors and basophilic AHF invasive to hepatic veins (Figures 1 and 3). Kakehashi et al. (2010) reported that CK8/18 was overexpressed in hepatocellular carcinomas, especially in oval cells. These data indicate that CK8/18 expression might be related to hepatic tumor progression or malignancy.
In the present study, CK8/18-negative basophilic AHF and hepatocellular adenomas were recognized (Figure 3). Histopathologically, we could not find any difference between CK8/18-positive basophilic AHF and CK8/18-negative basophilic AHF. Kawai et al. (2010) got a similar result from a male ICR mouse study. These results differ from the results that Kakehashi et al. (2010) reported, namely, that all basophilic AHF and tumors were stained positively for CK8/18. However, all AHF recognized in the Kakehashi et al. study were basophilic type because they administrated DEN in the preweaning period. In our study, in contrast, not only basophilic but also eosinophilic and clear-type AHF were observed because we treated in the postweaning period. There is the possibility that the origin cells of AHF or the mechanisms of the AHF formation were different between preweaning and postweaning initiation, but this remains unclear. Further investigations are needed to answer this question.
CK19 is also one of the intermediate filaments and is expressed in biliary epithelial cells but not normally in hepatocyte in adult liver. In humans, hepatocellular carcinomas that are positively stained for CK19 by immunohistochemistry are reported to have a worse prognosis than CK19-negative hepatocellular carcinomas (Andersen et al. 2010; Lee et al. 2006). Andersen et al. (2010) reported that the expression of CK19 was related to the malignancy of rat liver neoplastic lesions and CK19 was a prognostic marker of early neoplastic lesions in the rat liver carcinogenesis (Andersen et al. 2010). In this study, CK19-positive reactions were observed in both neoplastic and preneoplastic lesions, especially in the eosinophilic AHF (Figure 3). To our knowledge, this is the first report that CK19 is expressed in mouse hepatic preneoplastic and neoplastic lesions. Unfortunately, because there were only four hepatocellular adenomas and no carcinomas observed in the current study, the relationship between CK19 expression and the malignancy of mouse liver neoplastic lesions was unclear in this study. Further investigations are needed to resolve this question.
In conclusion, we demonstrated that DEN induced preneoplastic lesions in a dose-dependent manner in C57BL/6 mice. In addition, CK8/18-positive foci appeared to be appropriate marker lesions for evaluation of hepatocarcinogenesis. We conclude that suitable DEN treatments range from 75 mg/kg for 4 weeks (total = 300 mg/kg) to 50 mg/kg for 8 weeks (total = 400 mg/kg). The results suggest that the C57BL/6 strain has the potential to be suitable for postweaning period models, paving the way for future studies using the C57BL/6 strain as the background in transgenic models.
Footnotes
Financial support for this study was provided by NIH/NCI (R01-CA100908 to J.E.K.). All authors disclose that there are no conflicts of interest that could inappropriately influence the outcome of the present study.