
Abstract
The rabbit is occasionally used for inhalation and intranasal safety assessment studies, but there are no detailed descriptions of the anatomy or histology of the rabbit nose. To address this deficit, the nasal cavities of thirty-two control adult rabbits were sectioned and examined to provide mapping of the main epithelial types and histological structures present within the cavity and turbinates. Four levels of the nasal cavity were prepared and examined using anatomic landmarks. Level I was sectioned immediately posterior to the incisors, Level II at the first palatal ridge, Level III immediately anterior to the first upper premolar teeth, and Level IV immediately anterior to the first upper molar. Level I was lined predominantly by squamous epithelium with small amounts of thick transitional epithelium, and examination is recommended only for studies involving test article administration via instillation. Level II was lined primarily with transitional and respiratory epithelia, whereas Levels III and IV were lined with respiratory and olfactory epithelia and often contained nasal-associated lymphoid tissue. The vomeronasal organs were evident only in Level II. The similarities and differences of these features are compared with those of other common laboratory species (rat, mouse, dog, and cynomolgus monkey) and man.
Introduction
As the portal of entry to the respiratory tract, the nasal cavity is a primary site for injury induced by inhaled substances, whether microorganisms or xenobiotics. Although the nasal cavity is not usually examined in great detail in conventional toxicity studies in which substances are administered orally or by parenteral routes, it is carefully examined histologically when drugs are administered by inhalation (Greaves 2007) or intranasal instillation. Over the past decade, inhalation has increasingly been used as a drug delivery route to treat a range of acute and chronic health conditions in humans. The advance in inhalation therapy has resulted in more scientific information on the structure and function of the nose at anatomic, microscopic, and molecular levels. There is extensive literature about the anatomic and histological features of the nasal cavity of mice (Herbert and Leininger 1999; Mery et al. 1994), rats (Boorman et al. 1990; Harkema 1990; Harkema 1991; Harkema et al. 2006; Mery et al. 1994; Uraih and Maronpot 1990; Young 1981), and monkeys (Harkema 1990; Harkema 1991; Harkema et al. 2006; Macri et al. 2007). Although relatively uncommon, the rabbit has been used as the species of choice in some inhalation toxicological studies. Identification of toxicologically induced changes in the nasal turbinates of any animal species relies on the pathologist having a thorough understanding of anatomic and histologic properties of the normal nasal cavity of that species. Currently there is neither a detailed description of the rabbit nasal cavity nor a systematic approach to sectioning the rabbit nose in the published literature. The purposes of the present paper are to describe in detail a standard method of preparing the rabbit nasal cavity for histopathologic examination and describe the normal microscopic anatomy of the rabbit nasal cavity.
Material and Methods
Animals
Thirty-two New Zealand white rabbits of each sex, six to eight months of age, were used to describe the histological aspects of the nasal cavity. The rabbits were free of pathogens and respiratory disease (based on health and pathology reports by the supplier). Animals were from the control groups of two different inhalation studies and had acetate buffer at pH 5.0 administrated via instillation into the opening of their nares for twenty-eight days. Humane care was provided in compliance with the United States Department of Agriculture federal regulations.
All rabbits were euthanized by exsanguination following intravenous injection of sodium pentobarbital. The head was separated from the body, and all extraneous tissues, including the mandible, brain, eyes, skin, and muscle, were removed.
Fixation
The head was partially immersed in 10% neutral-buffered formalin, and fixative was introduced into the nasal cavity by retrograde perfusion through the nasopharynx, using a syringe. The nasal cavity was then fully immersed in 10% neutral-buffered formalin and fixed with the head for a minimum of seven days prior to sectioning.
Decalcification
The fixed nose was wrapped in gauze and immersed in a hydrochloric acid/water 25:1 decalcification solution for eight days. The decalcification solution was renewed at least four times during the process.
Sampling
Following decalcification, the nose was rinsed in running water overnight. Transverse sections (up to 5 mm) were made at four levels and processed in 28 × 35 × 6 mm cassettes with the cranial surface facing down. Landmarks for sectioning were based on the upper incisor teeth, palatal ridges, and first upper premolar and first upper molar teeth (Figure 1 ). Level I was sectioned immediately posterior to the incisors, over the incisive papilla. Level II was sectioned at the first palatal ridge. Level III was sectioned immediately anterior to the first upper premolar teeth. Level IV was sectioned immediately anterior to the first upper molar (Figure 1 ).
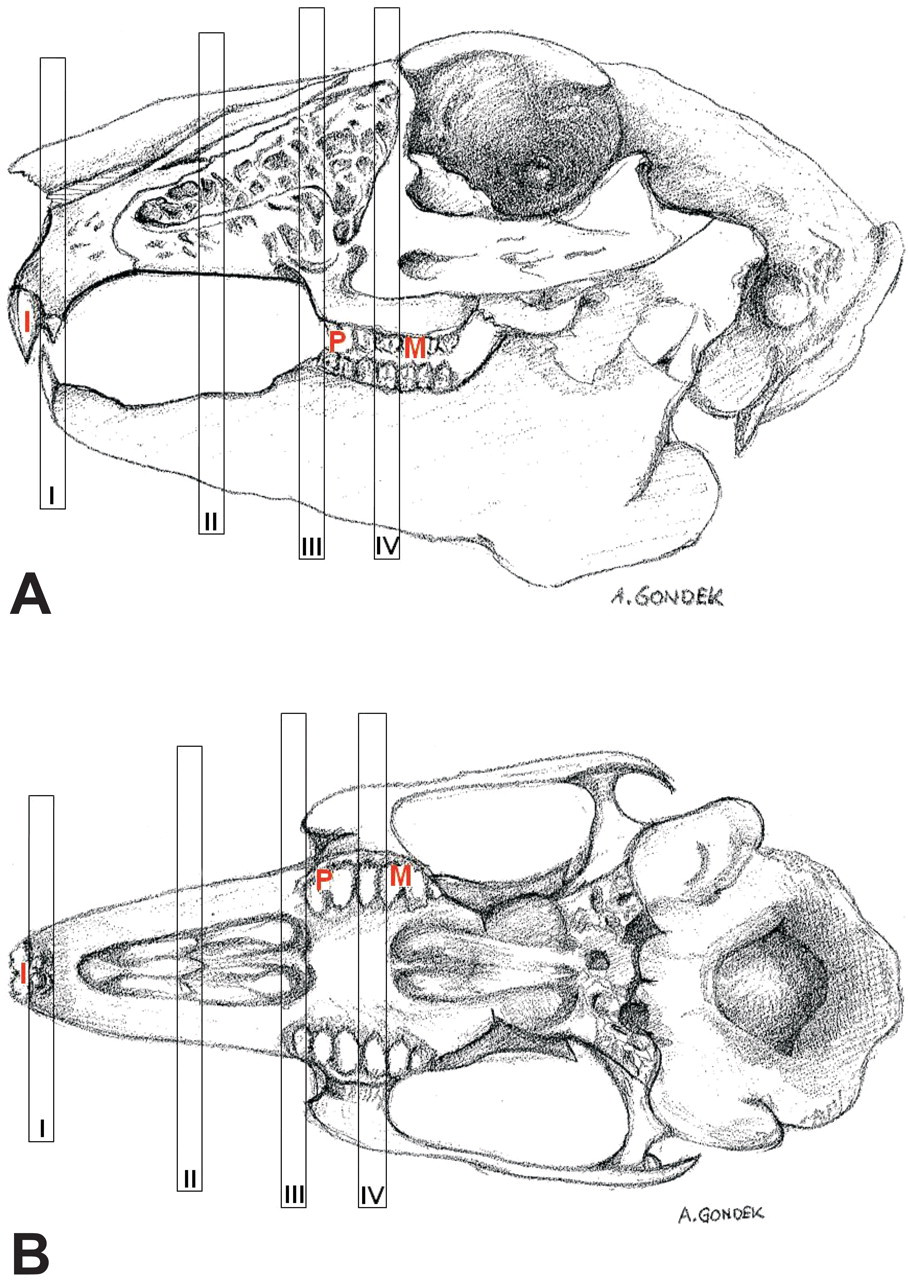
Diagrammatic representation of the dental landmarks for sampling the nasal cavity of the rabbit. A, lateral view; B, ventro-dorsal view; I, incisor tooth; P, first premolar; M, first molar; I, Level I; II, Level II; III, Level III; IV, Level IV.
Processing and Sectioning
Tissues were processed and embedded in paraffin wax using conventional methodology. Five-micron–thick sections were microtomed from the anterior face of each level, mounted on 75 × 25 × 1 mm glass slides, stained with hematoxylin and eosin (H&E), and examined microscopically.
Results
Level I
This first level was obtained immediately posterior to the upper incisor teeth (Figures 1A, 1B). The roots of the incisors are shown ventrally and inserted in the incisive bone (Figure 2A–C). Clusters of serous glands are located ventral to the nasal cavity and dorsal to the incisive bone (Figure 2C). The nasal cavity is divided sagittally into equal halves by the nasal septum. A plate of hyaline cartilage supports the nasal septum and extends dorsally and then laterally, supporting the outer surface of the nose. It then extends medially as the cartilage of the nasoturbinate and divides the nasal cavity into two nasal meatus, the right meatus and the left meatus.
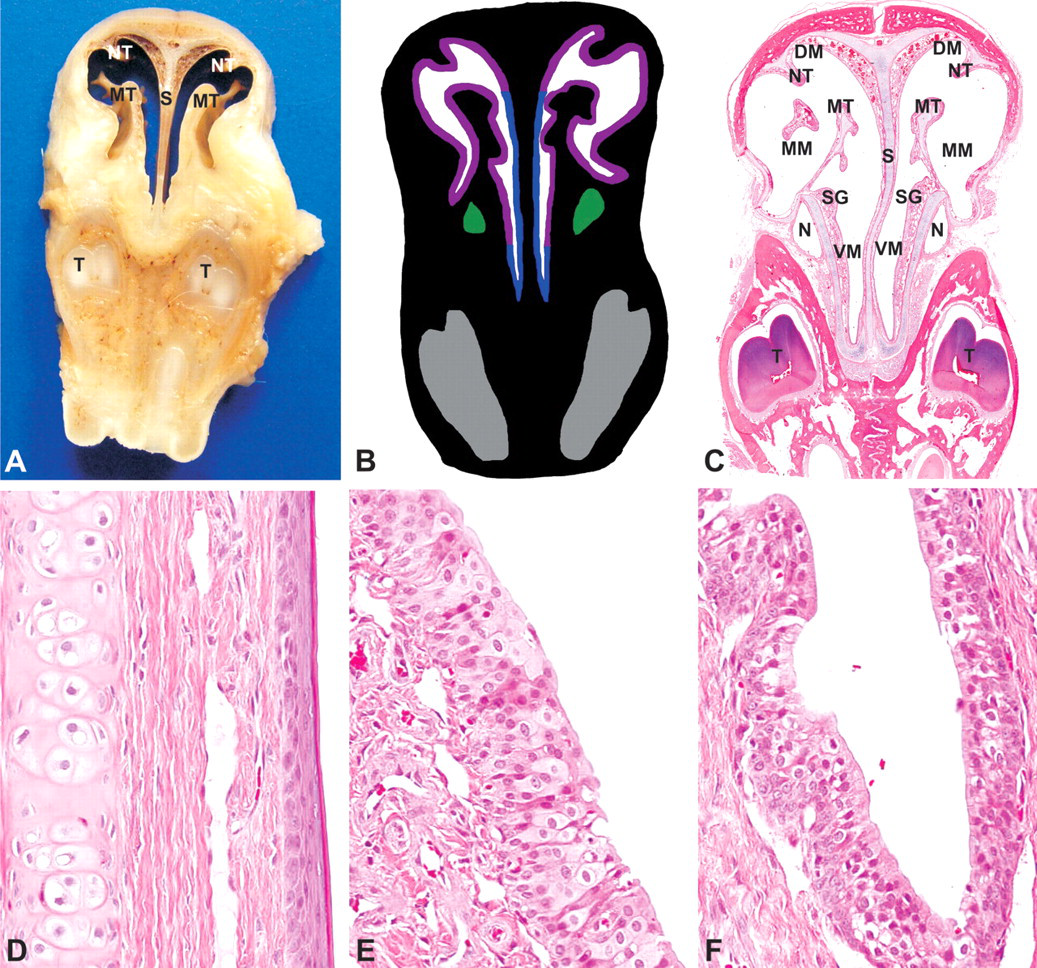
Rabbit, nasal turbinates, Level I-(A) Gross cross section. (B) Cross-section representation of Level I. Blue, squamous epithelium; purple, transitional epithelium; green, nasolacrimal duct; grey, tooth. (C) Subgross cross section, hematoxylin and eosin (H&E). NT, nasoturbinate; MT, maxilloturbinate; DM, dorsal meatus; MM, middle meatus; VM, ventral meatus; S, nasal septum; N, nasolacrimal duct; SG, serous glands; T, tooth. (D) Squamous epithelium, H&E, 200×. (E) Transitional epithelium, H&E, 200×. (F) Nasolacrimal duct, H&E, 200×.
At this level, the nasal cavity is lined by squamous epithelium and transitional epithelium (Figures 2B, D–E). Squamous epithelium lines most of the nasal septum. This epithelium is composed of four to six layers of squamous epithelial cells. It is overlaid by approximately 8 µm of keratin (Figure 2D). Serous glands with fine, intracytoplasmic, eosinophilic granules are occasionally present in the lamina propria.
Transitional epithelium, up to six cells thick, lines the lateral walls and medial and dorsal surfaces of the nasal turbinates. It is composed of up to two layers of basal cells, underlying four layers of polyhedral to cuboid cells (Figure 2E). The nasolacrimal duct is located ventrolaterally in the nasal turbinate and is lined with pseudostratified columnar epithelium (Figures 2B, C, E).
Level II
This level was taken through the first palatal ridge (Figure 1A, B). There are three distinct meatus: a dorsal meatus, a middle meatus with adjacent maxillary sinuses, and a ventral meatus. Also present at this level are two distinct turbinates: the nasoturbinate and the maxilloturbinate.
The roots of the upper incisor teeth are prominent at Level II and are located laterally (Figures 3A–C). Adjacent and medioventral to the incisor teeth are the nasolacrimal ducts, which are lined with simple columnar epithelium (Figures 3B–C).
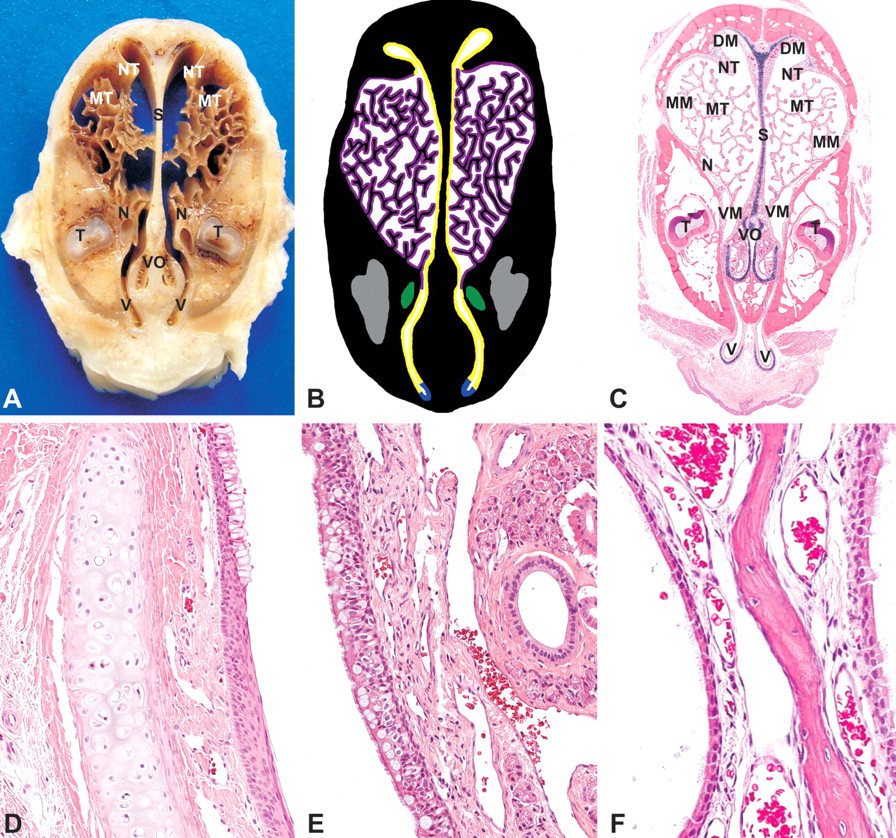
Rabbit, nasal turbinates, Level II-(A) Gross cross section. (B) Cross-section representation of Level II. Blue, squamous epithelium; yellow, respiratory epithelium; purple, transitional epithelium; green, nasolacrimal duct; grey, tooth. (C) Subgross cross section (H&E). NT, nasoturbinate; MT, maxilloturbinate; DM, dorsal meatus; MM, middle meatus; VM, ventral meatus; S, nasal septum; N, nasolacrimal duct; T, tooth; V, vestibule; VO, vomeronasal region. (D) Vestibule (squamous to respiratory epithelium transition), H&E, 200×. (E) Respiratory epithelium (septum), H&E, 200×. (F) Transitional epithelium, H&E, 200×.
At this level, the vestibule is located at the most ventral part of the nasal cavity, dorsal to the soft palate (Figure 3A–C, E). It has a sac-like shape and extends below the incisive bone. An inverted C-shaped cartilage ring supports the most ventral portion of the vestibule. The vestibule is lined with up to eight layers of slightly keratinized squamous epithelium. Nasal-associated lymphoid tissue (NALT) is often present in the vestibular mucosa.
The paired vomeronasal organ is visible near the ventral aspect of the septum and is partially surrounded by a descending hook-shaped cartilage (Figure 3C). Two types of epithelium compose the vomeronasal organ: olfactory and respiratory epithelia. The olfactory epithelium lines the medial aspect with nerve fiber bundles extending into the lamina propria. The lateral aspect of the organ is lined with ciliated, pseudostratified respiratory epithelium. Numerous heterophils transmigrating the respiratory epithelium of the vomeronasal organ or forming microabscesses were frequently observed and may represent background pathology related to instillation of the vehicle. The lamina propria of the respiratory mucosa contains glands that extend into the lamina propria of the nasal septum.
The nasal septum is supported by a plate of hyaline cartilage in an inverted anchor shape, which terminates in the nasoturbinate (Figure 3C). It is covered by a pseudostratified columnar, ciliated respiratory epithelium (Figure 3E). Goblet cells are more prominent in the most ventral surface. The lamina propria contains variable numbers of glands. The respiratory epithelium often has a scalloped profile at the dorsal aspect of the nasoturbinate.
There are three maxilloturbinates. The most dorsal is the most complex (Figure 3A–C), with as many as fifty or more branches. The maxilloturbinates as well as the lateral walls of the middle meatus are lined with transitional epithelium, which ranges in appearance from cuboidal to almost squamous cells. The epithelium is sparsely ciliated and lacks goblet cells (Figure 3F). Glands are present in the lateral wall of the middle meatus. The nasolacrimal duct is lined with squamous epithelium.
Level III
This level was taken immediately anterior to the first premolar teeth (Figure 1A, B). The two halves of the nasal cavity may or may not communicate at this level depending on the location of the section relative to the fully formed pharyngeal duct. The turbinates are lined with respiratory and olfactory epithelia. The paranasal sinuses are lined by a respiratory epithelium, made up of ciliated, pseudostratified, columnar cells, but goblet cells are absent. The lamina propria of this epithelium is occupied by two populations of glands, a periodic acid-Schiff–positive and a periodic acid-Schiff–negative population. Respiratory epithelium, without glands, also lines the lateral and ventral aspects of the ethmoid turbinates and ventral portion of the nasal septum (Figure 4D). The medial and dorsal aspects of the ethmoid turbinates, as well as the majority of the nasal septum, are lined with olfactory epithelium. The olfactory epithelium is a pseudostratified, columnar epithelium with up to ten layers of nuclei. The lamina propria of the olfactory epithelium contains prominent nerve bundles, glands, and blood vessels (Figure 4D).
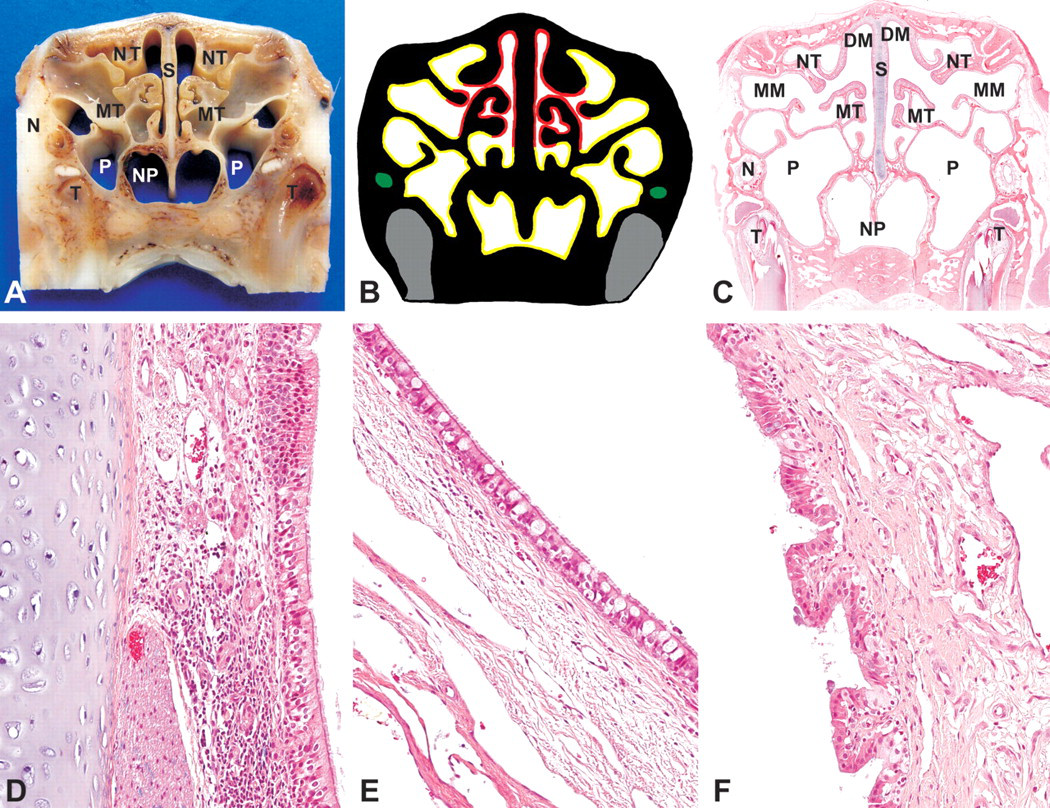
Rabbit, nasal turbinates, Level III-(A) Gross cross-section. (B) Cross-section representation of Level III. (C) Subgross cross section (H&E). NT, nasoturbinate; MT, maxilloturbinate; DM, dorsal meatus; MM, middle meatus; S, nasal septum; N, nasolacrimal duct; T, tooth; NP, nasopharyngeal duct; P, paranasal sinuses. (D) Septum mucosa—respiratory to olfactory epithelium, H&E, 200×. (E) Respiratory epithelium (nasopharyngeal duct), H&E, 200×. (F) Squamous to cuboidal epithelium (nasolacrimal duct), H&E, 200×.
NALT is occasionally evident in the nasal septum, primarily at the transition from respiratory to olfactory epithelium (Figure 4D) and in the nasopharyngeal duct mucosa, mostly ventrolateral to the lumen. The nasopharyngeal duct is lined with ciliated, pseudostratified, columnar epithelium with goblet cells (Figure 4E). The nasolacrimal ducts are located laterally at this level and are lined with a simple columnar epithelium. Its lamina propria is highly vascular (Figure 4F). Ventral to the nasolacrimal ducts are the roots of the first upper premolar teeth (Figure 4A–C).
Level IV
Level IV was sectioned immediately anterior to the first upper molar (Figure 1A, B). The roots of the first molar are lateral to the nasopharyngeal duct (Figure 5A–C). The nasopharyngeal duct is fully formed ventrally and does not communicate with the nasal cavity. Pseudostratifed, ciliated, respiratory epithelium with goblet cells lines the nasopharyngeal duct (Figure 5E). Paranasal sinuses are still evident and are still lined with ciliated respiratory epithelium that lacks goblet cells. Numerous glands are present within its lamina propria (Figure 5F). The nasal septum and most of the ethmoid turbinates are lined with olfactory epithelium. Numerous large, fibrous nerve bundles, in addition to glands and blood vessels, compose the lamina propria of the olfactory epithelium (Figure 5D). NALT may be present in the nasopharyngeal mucosa (Figure 5E) and in the transitional zones of respiratory to olfactory epithelium. In the dorsal part of the section the olfactory lobes of the brain may or may not be visible.
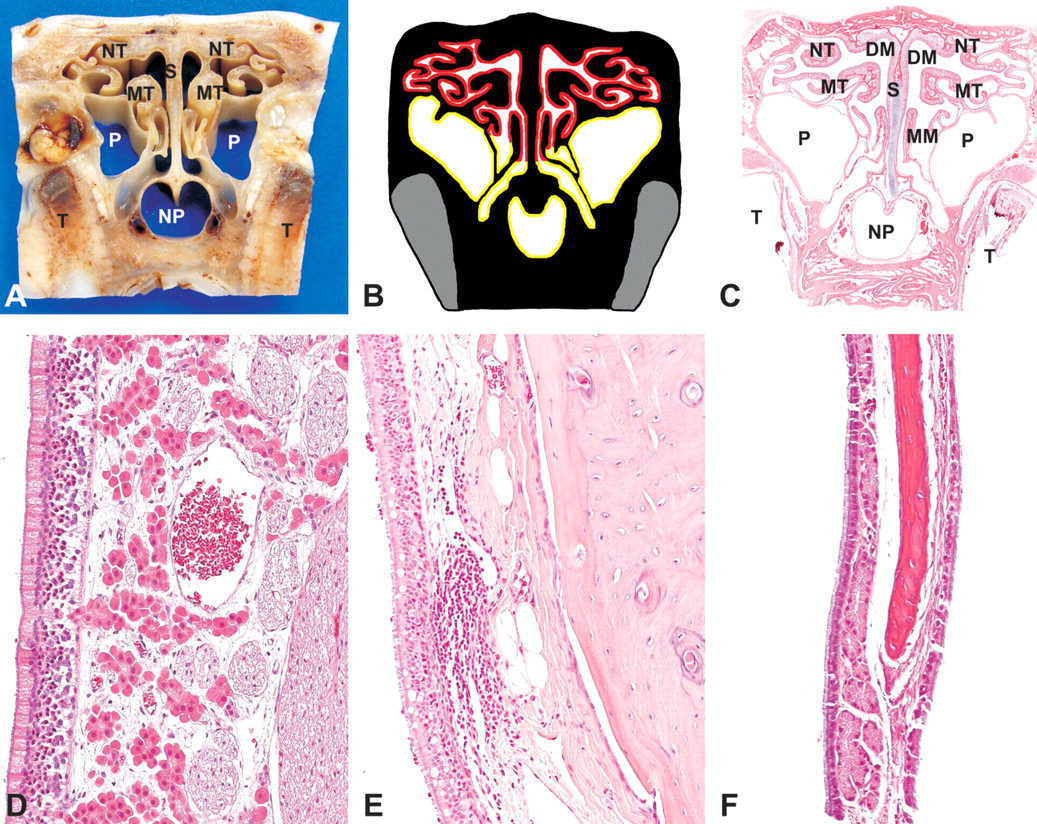
Rabbit, nasal turbinates, Level IV-(A) Gross cross-section. (B) Cross-section representation of Level IV. Yellow, respiratory epithelium; red, olfactory epithelium; grey, tooth. (C) Subgross cross-section (H&E). NT, nasoturbinate; MT, maxilloturbinate; DM, dorsal meatus; MM, middle meatus; S, nasal septum; T, tooth; NP, nasopharyngeal duct; P, paranasal sinus. (D) Olfactory epithelium, H&E, 200×. (E) Respiratory epithelium (nasopharyngeal duct), H&E, 200×. (F) Respiratory epithelium (paranasal sinus), H&E, 200×.
Discussion
The purpose of this article was to describe a method for preparing consistent sections of the rabbit nasal cavity for histopathologic examination. Landmarks for trimming levels I and II of the rabbit are similar to those recommended for rats (Boorman et al. 1990; Uraih and Maronpot 1990; Young 1981), but anatomic and histological differences exist between the species. Unlike in rats, Level I in rabbits is lined predominantly with squamous epithelium with minimal amounts of transitional epithelium. It resembles the cranial aspect of the rat nose anterior to the incisors, which is not routinely evaluated in rat inhalation toxicity studies (Mery 1994). In rabbits, the mucosa lining the ventral surface of the nose is continuous with the mucosa that overlays the external surface of the nares, and it is lined with highly keratinized squamous epithelium. Unlike other laboratory animals, this mucosa in rabbits contains some hair follicles. The human nasal cavity also contains hair follicles (Harkema et al. 2006). The second type of squamous epithelium (the thinner four- to six-layered squamous epithelium) that is evident at Level I sections from the rabbit nose is similar to the squamous epithelium that lines the vestibule in this and other laboratory species. Since Level I is recommended only to evaluate resistant squamous epithelium, evaluation of this level is recommended only to evaluate irritancy in studies involving direct instillation of the test material into the nares.
The transitional epithelium in the rabbit nose at Level I is relatively thick and more closely resembles that of the monkey nose than that of the rodent. In rats and mice, the transitional epithelium is one or two cells thick and pseudostratified (Montiero-Riviere and Popp 1984), whereas in monkeys it is thick, with up to five cell layers, and stratified (Harkema et al. 1987). The stratified nature of the transitional epithelium at this level should not be confused with squamous metaplasia in rabbits and monkeys. A thinner, pseudostratified, transitional epithelium lines the maxilloturbinates in rabbits at Level II and resembles that seen in the rodent nose (Monteiro-Riviere and Popp 1984). As in rats, but unlike primates, the transitional epithelium in rabbits is restricted to the lateral walls of the nasal cavity. In monkeys, transitional epithelium is present on both the lateral and septal walls of the anterior nasal cavity (Harkema et al. 1987).
The characteristic ciliated respiratory epithelium that is seen in Level I sections from rats (Boorman et al. 1990; Herbert and Leininger 1999; Young 1981) is not present in Level I sections of rabbits, but it occurs in all other levels. Respiratory epithelium with goblet cells is present in Levels II through IV. As in rats and mice, goblet cells are predominantly located in the rabbit mucosa lining the septum and nasopharynx (Harkema et al. 1989; Herbert and Leininger 1999).
The olfactory epithelium, which is evident in Levels III and IV in rabbits, is morphologically similar to that described for other species (Boorman et al. 1990; Harkema 1991; Harkema et al. 2006; Uraih and Maronpot 1990). In rabbits, this epithelium occupies a significant portion of the nasal cavity, and in this respect it is more similar to rats, mice, and dogs than to primates (including man), which have relatively little olfactory epithelium (Harkema et al. 2006).
The vomeronasal organ is a paired tubular diverticulum located in the ventral portion of the nasal septum of most mammals (e.g., dogs, cats, rodents, and rabbits), and it is thought to contribute to olfaction (Harkema 1991). It is poorly developed in higher primates and is absent in adult humans (Harkema 1991). The vomeronasal organ that is seen in Level I sections from rats (Boorman et al. 1990; Herbert and Leininger 1999; Young 1981) is not present in Level I but is present in Level II sections of rabbits. As in rats, both respiratory and olfactory epithelia form the rabbit vomeronasal organ (Uraih and Maronpot 1990). The occurrence of heterophils/neutrophils or any other inflammatory cell transmigrating the vomeronasal mucosa and/or forming microabscesses has not been described as a normal feature in any other species. Although the cause of the heterophil movement and abscess formation in the rabbit vomeronasal mucosa is unknown, nasal instillation of the vehicle (acetate buffer at pH 5.0) may be a factor.
Sections of the rabbit nose at Level II show unique histological features. Plates and rings of hyaline cartilage are prominent underlying the vestibule and surrounding the vomeronasal organ.
Another unique histological feature in rabbits is the presence of complex maxilloturbinates, with multiple branching. Other small laboratory rodents also have complex maxilloturbinates, although they are much less branching (Harkema et al. 2006). In rabbits and rodents the maxilloturbinates provide better protection of the lower respiratory tract, by better filtration, absorption, and disposal of airborne particles and gases, than does the less complex human nose (Harkema et al. 2006).
Different from other species, the roots of the incisor teeth in rabbits are ventral at Level I but lateral at Level II. The roots of incisor teeth are always located laterally at Level I in rats (Uraih and Maronpot 1990) and mice (Herbert and Leininger 1999).
Rabbits have one pair of paranasal sinuses (maxillary sinuses), which are located in the lateral wall of the nasal cavity. Histologically, the rabbit paranasal sinuses resemble those of rats, with similar ciliated, columnar epithelium and a lamina propria with prominent glands (Boorman et al. 1990).
In the majority of laboratory species, NALT is located at the entrance of the nasopharyngeal duct (Spit et al. 1989). In rats, NALT is restricted to the ventral aspects of the lateral walls at the nasopharyngeal duct opening (Harkema 1991). In monkeys, NALT is more abundant than in rats and is located on both lateral and septal walls of the proximal nasopharynx (Haley 2003). Nasal-associated lymphoid tissue also occurs in the rabbit nose and is located mostly in the vestibule, nasopharyngeal duct mucosa, and transitional zones (respiratory to olfactory epithelium). Based on personal experience, NALT is more widespread in rabbits than rats (but less abundant than in monkeys and dogs). Although the function of NALT and its place in the general mucosal-associated lymphoid system have not yet been characterized, NALT in laboratory animals may have an important function in regional immune defense of the upper airways (Harkema 1991).
The rabbit nasolacrimal duct originates at the medial canthus of the eye, is first seen nearest its origin in Level III, and terminates ventromedially in the nasal turbinate of Level I. At its origin, the nasolacrimal duct is lined with squamous epithelium, but elsewhere the duct is lined by pseudostratified nonciliated columnar epithelium. Unlike that of rabbits, the rat nasolacrimal duct terminates at the ventromedial wall of the nasal vestibule (Uraih and Maronpot 1990). Stratified squamous epithelium is normally found at the origin and termination of the rat nasolacrimal duct, and pseudostratified noncliated columnar epithelium is found elsewhere (Uraih and Maronpot 1990).
Conclusion
We have demonstrated a method for evaluating the rabbit nasal cavity in safety assessment studies and compared the histological features with those of other commonly used laboratory species and man. The technique uses easily recognizable anatomic landmarks for trimming and produces four consistently reproducible sections representing the entire structure and all epithelial types. Level I is predominantly lined with squamous epithelium, with minimal amounts of thick transitional epithelium. This transitional epithelium should not be confused with squamous metaplasia. Level II is lined mostly with transitional and respiratory epithelia. The vomeronasal organs and vestibule are evident only in Level II. Levels III and IV are lined with respiratory and olfactory epithelia, and NALT is relatively common in these levels.
Footnotes
Acknowledgment
The authors wish to thank Janet Hammell and Cyndi Pillsbury for the assistance with trimming, processing, and slide preparation; Aleksandra Gondek for drawings; and Helen Wilkinson for image preparation.
Acknowledgements
The author(s) declared no potential conflicts of interests with respect to the authorship and/or publication of this article. The author(s) received no financial support for the research and/or authorship of this article.