
Abstract
The principle of microbeam radiotherapy (MRT) is the delivery of a single high-dose fraction to a large treatment area divided into several smaller fields, to reduce the treatment's overall toxicity. Since the fundamental concept of MRT was first established, a notable tissue-sparing effect (TSE) has been confirmed in a large variety of species and tissue types, although the underlying biological mechanism in this process remains unclear. By coupling high-precision MRT with an ex-vivo mouse spermatogenesis model, we demonstrated the significant testicular TSE for maintaining spermatogenesis following MRT. To our knowledge, this was the first TSE identified in reproductive tissue. Our high-precision microbeam analysis also revealed that an efficient TSE for spermatogenesis relies on the size of the non-irradiated germ stem cell pool in the irradiated testicular tissues, suggesting the involvement of stem cell migration/competition. These findings of testicular TSE indicate that radiation infertility is not dose-dependent, but instead depends on micro-dosimetric conditions, and that the limit of the TSE is a potential threshold for radiation-induced male infertility.
INTRODUCTION
Human spermatogenesis takes approximately 60–70 days for a spermatogonial stem cell (SSC) to become a fully functional sperm. Mammalian spermatogenesis requires the maintenance of a stem cell pool, amplification of spermatogonia, reductive division to haploid cells (meiosis), and the morphological transformation of the haploid cells into spermatozoa (spermiogenesis) (Clermont, 1972). This differentiation process is one of the most important developmental events for propagating life; thus, it should be highly robust. However, male fertility can be easily impacted by environmental, chemical, and physical stressors, including ionising radiation. During this long process in the testes, radiation exposure can easily lead to reproductive toxicity. In fact, doses of radiation >0.35 Gy cause the eradication and complete absence of sperm, which may be reversible (De Felice et al., 2019). The time needed for recovery increases as dose increases, and doses in excess of 2–6 Gy may result in a permanent loss of fertility (Fukunaga et al., 2017). Such radiation-induced male infertility is considered a tissue reaction.
Radiation-related effects at the tissue level appear to be dose-dependent; however, notable differences exist in the responses observed, depending on whether the radiation exposure is uniform. Microbeam radiotherapy, or MRT, is able to deliver a single high-dose fraction to a large treatment area, divided into several smaller fields, to reduce the treatment's overall toxicity (Fukunaga et al., 2021). The term MRT means the use of parallel 50–150 keV x-ray beams of micron width dimensions for potential therapeutic approaches (Slatkin et al., 1992). The German radiologist Alban Köhler was the first to report clinical observations of a tissue-sparing response during grid radiotherapy in which spatially fractionated radiation was delivered using a grid-like pattern of beams (Köhler, 1909). Zeman et al. (1959) found an extraordinary tolerance response in mouse brain tissue using microbeams. Since Slatkin et al. (1992, 1995) established the fundamental concept of MRT and demonstrated a noteworthy tissue-sparing response in rat brain tissue during MRT in the 1990s, extraordinary tolerance responses in normal tissues following MRT have been confirmed, namely, the tissue-sparing effect, or TSE, showing the great potential to offer significant improvements in clinical outcomes (Fukunaga, 2021). However, the underlying radiobiological mechanisms at the tissue level in response to non-uniform radiation fields have yet to be fully determined (Fukunaga and Prise, 2018).
This article outlines our recent work on fertility preservation with testicular TSE following MRT and the perspectives for extrapolation to radiation protection systems.
TISSUE-SPARING EFFECT FOR MAINTAINING SPERMATOGENESIS
Using a combination of a synchrotron-generated x-ray microbeam and a unique ex-vivo testicular tissue culture method (Fig. 1A), we have investigated non-uniform radiation-induced effects on the process of spermatogenesis (Fukunaga et al., 2018). The Acr-GFP transgenic mice express a meiosis-specific GFP, which can be visualised and graded according to developmental stage during spermatogenesis. Testicular tissues from 7 days postpartum mice were separated into small fragments (approximately 1–2 mm3 in size) to establish organ cultures and placed on sterile agarose gel stands half-soaked in αMEM medium containing 10% KnockOut™ Serum Replacement (KSR) medium. The GFP expression of the spermatogenic cells confirms the progression of spermatogenesis in the cultured tissues.
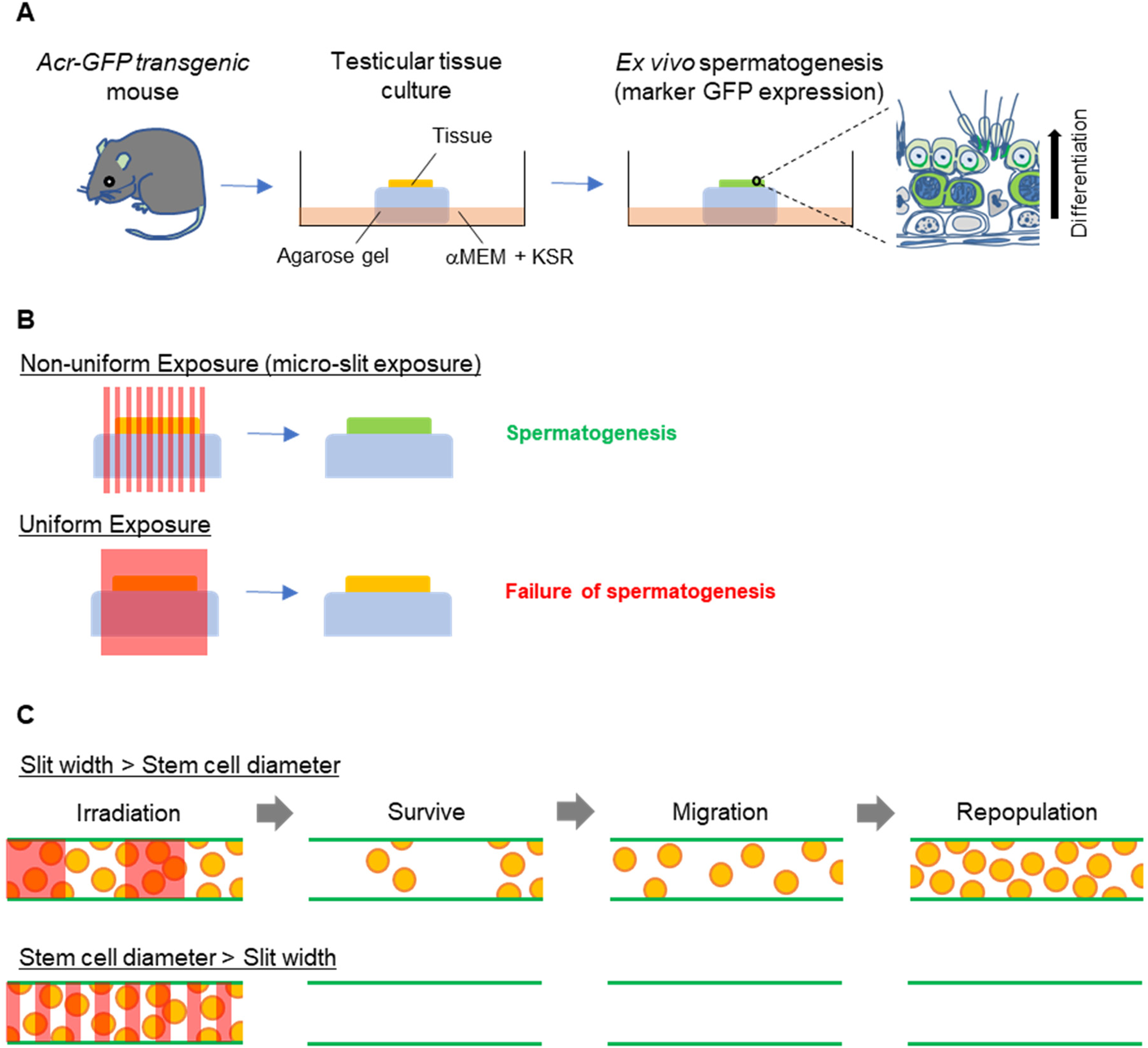
Tissue-sparing effect for maintaining spermatogenesis following microbeam radiotherapy. (A) Schematic representation of our ex-vivo spermatogenesis model. (B) The tissue-sparing effect of spatially fractionated radiation for maintaining spermatogenesis is shown. In the lower scenario, all stem cells are irradiated in a uniform radiation field, and no tissue-sparing response is observed. In the upper scenario, some of the stem cells in the non-exposed areas survived in micro-slit radiation fields. Then, the significant tissue-sparing effect occurs to maintain spermatogenesis. (C) Spermatogonial stem cell migration in the seminiferous tubule in response to spatially fractionated radiation fields. Two 50% micro-slit patterns were compared for the seminiferous tubules in which there are germ stem cells and the niche. They are irradiated at the same dose at the whole tissue level. In the lower scenario, all stem cells are irradiated in a narrow-slit radiation field, and no tissue-sparing response is observed. In the upper scenario, some of the stem cells in the non-exposed areas survive in the wide-slit radiation fields. Then, they migrate and repopulate, and the significant tissue-sparing effect occurs to maintain tissue functions. The red areas show the irradiated areas at the tissue level. Reproduced from Fukunaga (2021) with permission. © 2024 Radiation Research Society.
Recently, our microbeam analysis revealed a significant TSE in the culture testicular tissues following MRT and for maintaining spermatogenesis (Fukunaga et al., 2019) (Figs. 1B and 2). To our knowledge, this is the first study to show significant TSE as a way of fertility preservation. Furthermore, by changing the micro-slit width, we found that non-irradiated stem cells are required for significant TSE and that the migration step of the non-irradiated SSCs in the irradiated testes tissue likely underpins an effective TSE (Fig. 1C). This indicated that in the testicular tissue, tissue repair/remodelling in response to spatially fractionated radiation fields depends not only on the dose–volume parameters but also on biological parameters such as non-irradiated stem cell survival, migration, and repopulation. These may also occur in other systems.

Immunohistochemical images of ex-vivo testicular culture tissues following exposure to micro-slit x-ray microbeams. Immunohistochemical images of ex-vivo testicular culture tissues 1 h after 10-Gy 50-μm-slit x-ray microbeam irradiation. Staining for γ-H2AX, anti-GENA (anti-germ cell-specific antigen antibody), and Hoechst (for DNA) is shown as red, green, and blue, respectively. The red arrows indicate the micro-slit irradiated areas. Scale bar, 200 μm. Reproduced from Fukunaga et al. (2020a) with permission. © 2024 Radiation Research Society.
In 2020, in a follow-up to our initial report (Fukunaga et al., 2019), we showed that the TSE in irradiated testicular tissue was more effective when more SSCs survived after exposure to spatially fractionated radiation (Fukunaga et al., 2020a). These findings indicated that the distribution of radiation delivered to the testes at the microscale level is important for preserving male fertility, indicating that stem cell migration/competition is possibly involved in the underlying mechanisms of the TSE. In addition, the testicular TSE of spatially fractionated x-rays has considerable potential for future clinical applications, as it responds to a wide range of x-ray energies, although further mechanistic studies in vivo are required (Fukunaga et al., 2020b).
There are two types of radiation effects, namely, tissue reactions and stochastic effects, which are considered to be dose-dependent (ICRP, 2007). When it comes to tissue reactions, when a number of individuals are exposed to the same dose of radiation and certain symptoms (tissue responses) appear in 1% of them, that dose is considered to be the threshold. However, based on our results, we would like to propose a possible paradigm shift (Fig. 3). Although the current model assumes that a threshold dose exists for radiation-induced male infertility as a tissue reaction, our data show that germ stem cell viability is more important than the dose. In other words, the limit of TSE is a type of threshold. This novel concept may, of course, be common not only in radiation-induced male infertility, but also in other tissue reactions.
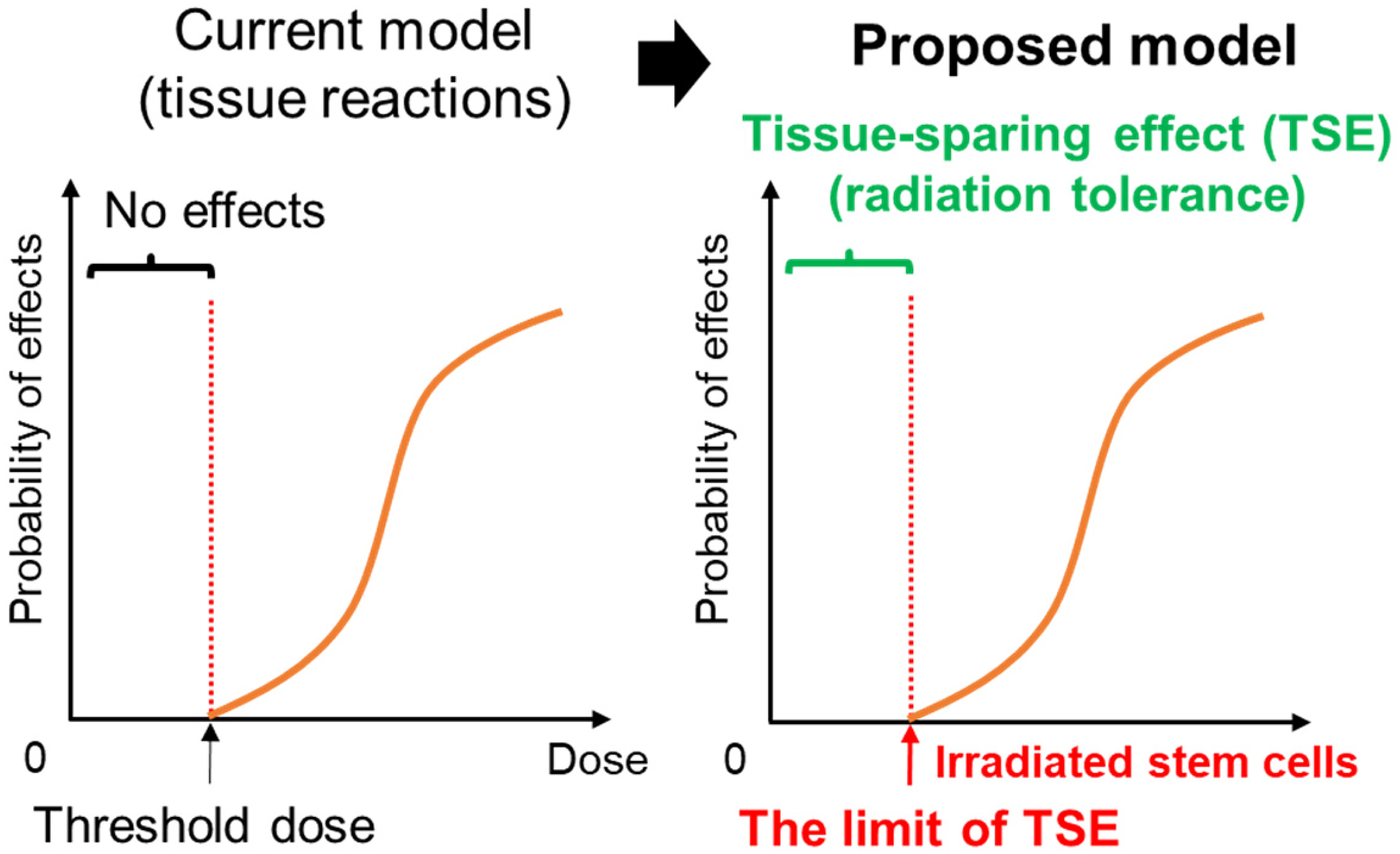
Possible paradigm shift. The current model assumes that a threshold dose exists for radiation-induced male infertility as a tissue reaction. Exposure to doses above the ‘threshold dose’ leads to radiation-induced male infertility. However, our data show that the limit of the TSE is the radiation tolerance threshold for radiation-induced male infertility. Exposure of stem cells in amounts exceeding the ‘limit of TSE’ leads to radiation-induced male infertility.
According to ICRP Publication 131, ICRP considers the characteristics of stem cells and the niche to be one of the most important factors for radiation-induced biological effects (ICRP, 2015). Microbeam studies and the available literature demonstrate that the concept of stem cell migration has great potential for predicting tissue-level responses, either during or after spatially fractionated radiotherapy (Fukunaga, 2021). In particular, with the technical combination of the ex-vivo spermatogenesis model and high-precision microbeam techniques, the TSE findings for maintaining spermatogenesis indicate the need for a reconsideration of the conventional dose–volume concept.
In conclusion, we demonstrated the significant testicular TSE of MRT for maintaining spermatogenesis. This was the first TSE identified in reproductive tissue. According to our results, the testicular TSE is not a dose–volume effect, and radiation-induced male infertility is not dose-dependent but instead depends on stem cell viability. Our findings may contribute to the improvement of radiological protection systems, as well as to the establishment of a novel radiotherapy to preserve male fertility.
Footnotes
ACKNOWLEDGEMENTS
We would like to express our sincere gratitude to all our collaborators in the world, past and present, for their continuous support.