
Abstract
Radiation risks for cancer are classified into probabilistic effects because the induction of oncogenic mutations in a single epithelial cell is thought to be causally related to the process of carcinogenesis. However, two observations remain unanswered by the theory. One is a parallel shift of mouse survival curves towards younger ages after a radiation exposure, and the other is a decreasing trend of relative risk for cancer with an increase in postexposure time. Both are readily explained if it is assumed only that a radiation exposure causes a shift of the spontaneous tumours to appear earlier as a result of tissue damage which favours a transformed cell(s) to grow faster. In line with this notion, there are reports showing that exposures to high doses of acute radiation did not increase the proportion of mice dying from tumours, while their lifespans were shortened. In contrast, although there are reports showing a dose-related increase in the proportion of tumour deaths, the studies were either prematurely terminated or the ages were adjusted, and they are not considered to support the tumour induction model of radiation. As it seems that radiation carcinogenesis is a type of tissue response, it should be more appropriately classified as a deterministic effect of radiation. It is added that this article deals mainly with the data obtained under high-dose and high–dose-rate exposure conditions.
A GAP BETWEEN EPIDEMIOLOGY AND BIOLOGY
It has long been known that an exposure to ionising radiation increases the risk of developing cancer, but no explanation has been provided to explain two puzzling observations: (1) There is a decreasing trend in the relative risk (RR) with an increase in time following a radiation exposure (Grant et al., 2017), and (2) the parallel shift of mouse survival curves towards younger ages following an exposure to radiation (the shape of the curve remains the same: e.g. Tanaka et al., 2003). Radiation biologists are primarily concerned with stem cell mutagenesis which involves DNA repair mechanisms and are indifferent to these observations. In this context, radiation biologists and radiation epidemiologists are isolated from each other. The present manuscript is an effort to try to bridge the gap between these two viewpoints.
Why the relative risk for cancer should decrease
It is well known that the RR for cancer declines with an increase in time since an exposure to radiation (e.g. Grant et al., 2017). Unfortunately, although the declines were commonly observed both in human epidemiology and in mouse experiments, they did not attract interest from radiation biologists because radiation biologists usually do not examine mouse data from an epidemiologic viewpoint, i.e. using the mouse·day approach to evaluate risks. In examining this data, it was observed that this decrease in the RR could be the result of a radiation-induced shifting of the mortality curve towards younger ages (Fig. 1A and B). To test this possibility, solid tumour mortality data from mice were valuable (e.g. Sasaki and Fukuda, 2005). Plotting the solid tumour mortality rate against the attained age revealed that the shape of the mortality curves for the irradiated population looked very similar to that of the control group but was shifted towards younger ages (Fig. 1C), and the shifted time was proportionally related to the dose. The findings suggest that this is a window to allow us to understand the declining trend of the RR.
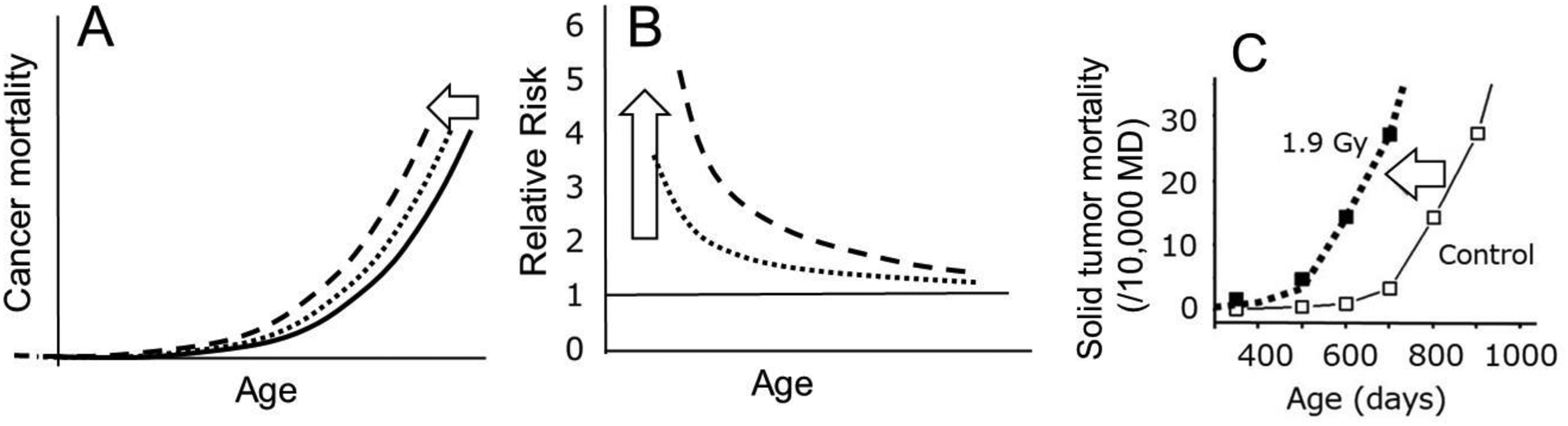
(A and B) Conceptual presentation to indicate that a parallel shift of the mortality curve towards younger ages necessarily gives rise to a highly elevated RR shortly after the exposure which is followed by its rapid decline. Different lines indicate different doses. (C) Mouse data for solid tumour mortality show that a simple shift of the control curve towards younger ages fits the observed points of the irradiated group (closed symbols) almost exactly (Nakamura, 2023). In this study, B6C3F1 female mice were exposed to 1.9 Gy of gamma rays (0.98 Gy min−1) at 7 days of age (Sasaki and Fukuda, 2005).
The next task is to try to understand the biological nature of this parallel shift in the survival curves. In the oncogenic mutagenesis model, it is assumed that a proportion of the irradiated individuals is affected and that the proportion increases with an increase in the radiation dose. Thus, there would be an excess of cancers that probably did not emerge in one's life if one were not exposed to radiation. However, in this scenario, there must be two subpopulations, one affected by an irradiation and the other not affected, and the mean life span is shortened in the exposed group. Under such conditions, the distribution of individual lifespans must become wider, and hence the mean slope of the survival or mortality curve should be shallower than that of the control curve. In other words, under such conditions, a parallel shift of the curve cannot be realised. To solve this problem, it suffices to suppose that all individuals in the irradiated group were affected. In other words, a radiation exposure would affect the future tumour bed to facilitate spontaneously transformed cell(s) into forming tumours earlier than usual rather than induce oncogenic mutations which lead to cancer development.
Simple parallel shift of the control curve also applies to human data
Since we can see a good fit of solid tumour mortality of irradiated mice by shifting the control curve towards younger ages (Fig. 1C), it was next of interest to see if the same results could be realised with human data. For this purpose, solid cancer mortality data from atomic bomb survivors were used.
Specifically, first, the cancer mortality rate at age 70 in the control group was multiplied by the RR at 1 Gy at an attained age of 70, which represents the cancer mortality rate of the 1-Gy–exposed group at 70 years of age. Then, shifting the mortality curve of the control group towards the left so that it may pass through this point of mortality can show a hypothetical age-dependent cancer mortality at 1 Gy. The shifted curve was found to fit closely to the epidemiologically estimated values (Fig. 2) and thus satisfied concerns of whether an earlier onset model would also work well in humans.
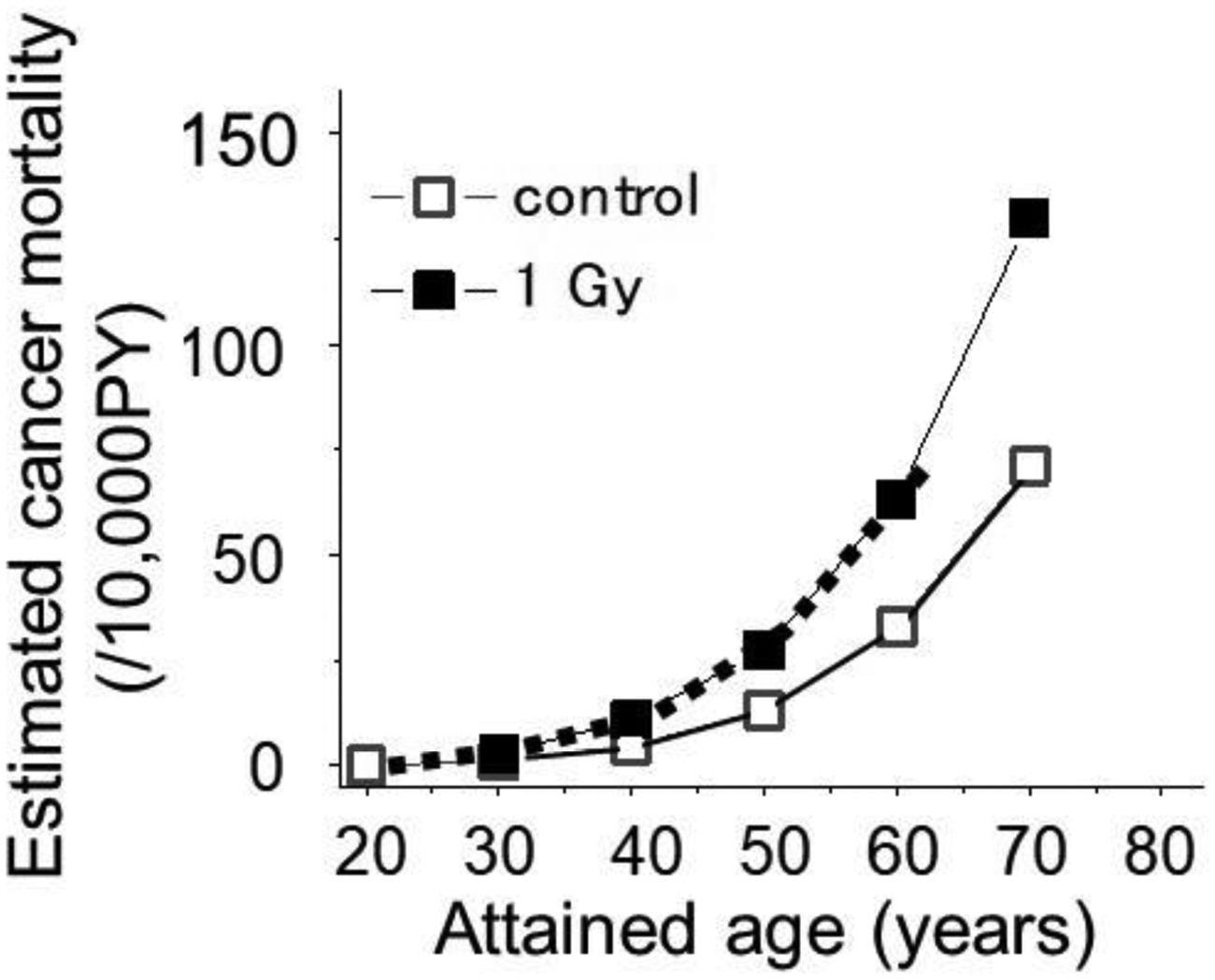
Solid cancer mortality data of atomic bomb survivors who were 10 years old at the time of exposure (Nakamura, 2023). A simple shift of the control curve (open symbols with a solid line) towards younger ages by 8.5 years (dotted curve) fit closely to the epidemiologically estimated mortality following an exposure to 1 Gy (closed symbols).
In this context, it should be mentioned that the present model does not apply to some types of malignancies such as childhood leukaemia and thyroid cancers, which show a short latency period and do not show an age-related rapid increase in the mortality rate.
In the literature, there are a substantial number of mouse studies which show a dose-related increase in the proportion or incidence of solid cancers. Such data are not compatible with the present earlier-onset model, and the discrepancy is due to the premature termination of the experiments or to age adjustments made when analysing the data. In both instances, the results do not seem to lend support to the mutagenesis hypothesis.
Prematurely terminated observations
In the past, and even today, irradiated animals were not supported until reaching a natural death but were sacrificed around the time when deaths started to occur in the control group. It is obvious that such an early termination of the observation only shows a cross-sectional picture of the complete process of radiation carcinogenesis. One example is shown in Fig. 3 where irradiated male B6C3F1 mice were sacrificed at 13 months of age. The proportion of mice bearing liver tumours may appear to increase sharply in relation to the dose, but this is only seen at this unique time point because the incidence of liver tumours in unirradiated mice may reach nearly 50% when they are fed until their natural death. In other words, Fig. 3 represents a cross-sectional view that tumours started to occur earlier in the exposed group, and the amount of shifted time is proportionally related to the radiation dose.
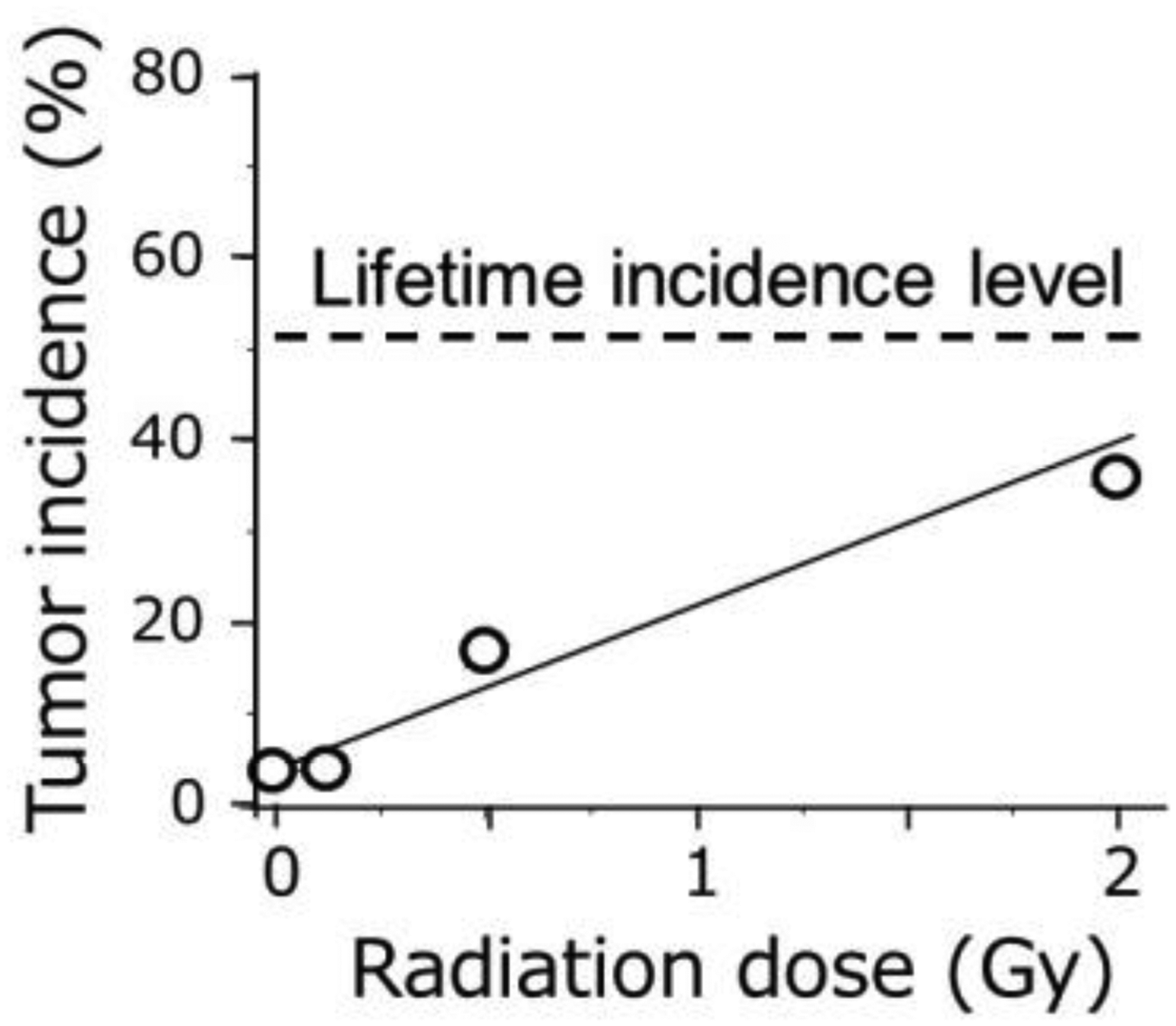
Incidence of liver tumours in B6C3F1 male mice (Takahashi et al., 1992). Mice were sacrificed at 13 months of age when over 90% of the animals were still alive. Because the lifetime incidence of liver tumours in the unirradiated mice is about 50% and does not significantly increase following irradiation (Tanaka et al., 2007), the dose–response slope observed here should become gradually shallower due to gradual increase in the tumour incidence in the control group with an extension of the observation period.
Another problem in describing the carcinogenic effects of radiation is age adjustment. The rationale behind the idea was to compensate for the shortened lifespan of the irradiated animals since cancer mortality occurs primarily later in life (Ullrich and Storer, 1979). In other words, it was determined what would have happened if there was no life shortening in the exposed group. Specifically, first adjust the number of surviving individuals in the exposed group to match that of the control group in each age group. Second, multiply the corrected number by the observed mortality for specific tumours. It is now understood that the observed mortality in the exposed group is in fact equal to the mortality of the control group, but the age was advanced by X days which is the amount of time the mortality curve was shifted towards the younger ages. Thus, the apparent dose-related increase in the age-adjusted cancer mortality represents the effect of a time shift in tumour development. For more details, see Nakamura (2024).
REPORTS WHICH SHOWED NO INCREASED CANCER MORTALITY
In contrast to the reports which showed an apparent increase in the cancer mortality or incidence, there are also publications which are based on lifetime observations and which failed to show such an increase. For example, Grahn and Sacher (1958) and Storer (1965) showed that the proportion of mice which died from tumours did not increase following exposures to radiation. More recently, Tanaka et al. (2007) reported pathologic data of a large-scale study of mice chronically exposed to 8 Gy (both the exposed and control groups consisted of 500 male and 500 female B6C3F1 mice). There was no significant increase in the proportion of mice dying from major types of malignancies.
RADIATION DOES NOT INDUCE CANCERS BUT ACCELERATES THE GROWTH OF SPONTANEOUS CANCERS
The mutation induction theory of radiation carcinogenesis supposes that only a fraction of the irradiated population is affected and that the fraction increases with radiation dose. In contrast, the present earlier-onset hypothesis assumes that all tumours which occurred in the exposed group were affected. Evidence which supports the latter hypothesis comes from a recent mouse study in which animals were sacrificed every 100 days to examine the occurrence of tumour cells (not macroscopic tumours). Interestingly, malignant lymphoma cells appeared at about the same age in both irradiated and non-irradiated groups (Tanaka et al., 2017), but death from malignant lymphoma occurred earlier in the irradiated group by about 100 days (Tanaka et al., 2007). Radiation-induced inflammation is a possible factor which could lead to an earlier onset of tumour development, and it appears that there are multiple pathways (and factors) involved in the process (Nakamura, 2021).
CANCER RISK FROM RADIATION SHOULD BE DETERMINISTIC
The present review clearly suggests that the induction of oncogenic mutations does not play a key role in radiation carcinogenesis but probably that radiation-induced tissue damage and subsequent inflammation could play a role (Fig. 4). If we accept the earlier-onset model, it will be necessary to change the paradigm and to reclassify the carcinogenic effects of radiation from a probabilistic effect to a deterministic effect because the earlier onset of cancer mortality resulted from the induction of some types of tissue responses, and a tissue response belongs to the category of deterministic effects. An apparently linear non-threshold response for radiation-related cancer risk in terms of RR would have to be understood from a new viewpoint.
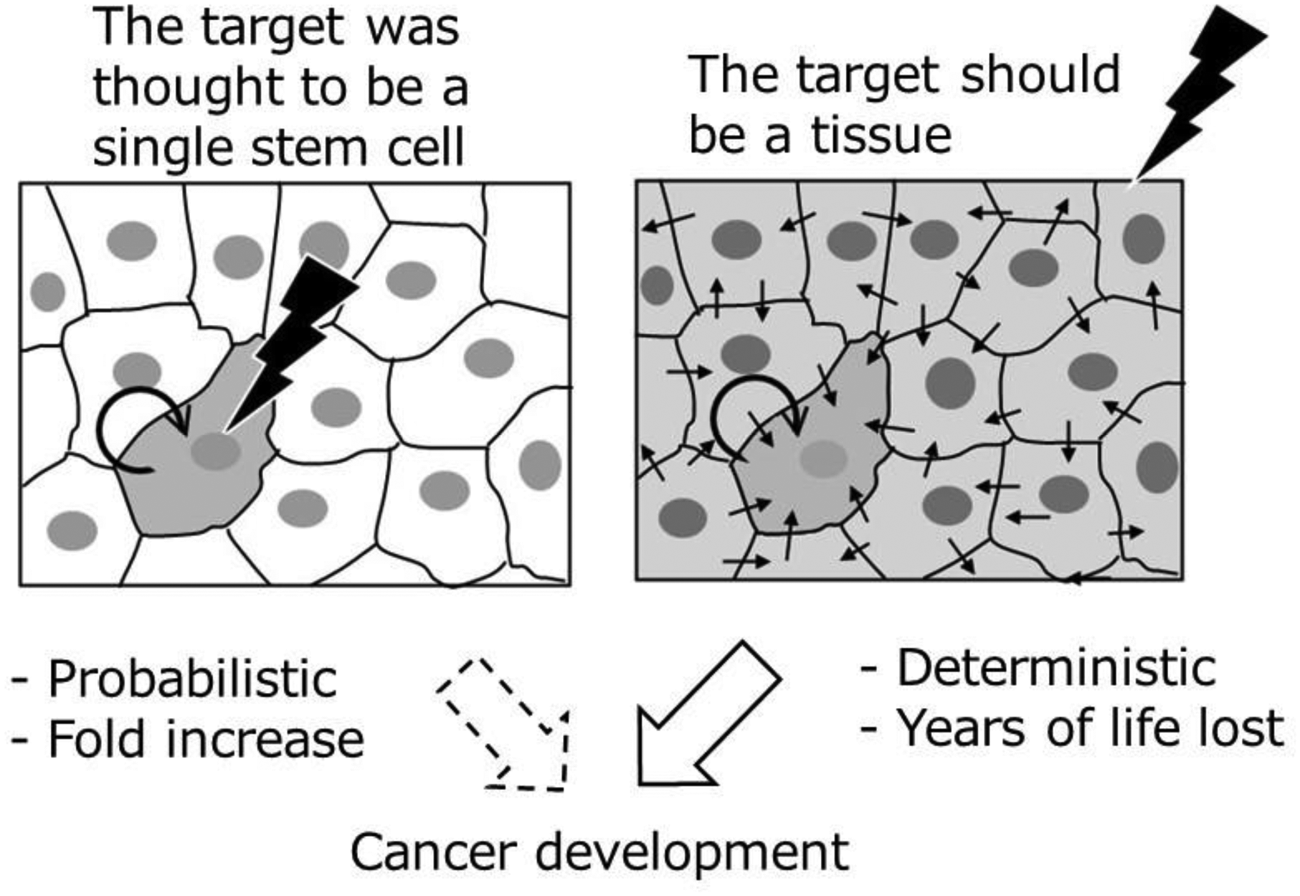
A schema which indicates differences in the target of radiation carcinogenesis. The left panel shows an irradiated single stem cell, and the right panel shows an irradiated tissue. The small arrows indicate possible factors which contribute to intercellular communications, and the thick circular arrows indicate the future multiplication of a transformed cell.
Footnotes
ACKNOWLEDGEMENTS
The author is grateful to Dr. Leon Kapp for his careful reading of the manuscript. The Radiation Effects Research Foundation (RERF), Hiroshima and Nagasaki, Japan, is a public interest foundation funded by the Japanese Ministry of Health, Labor and Welfare (MHLW) and the US Department of Energy (DOE). This publication was supported by RERF Research Protocol 1-75. The views of the author do not necessarily reflect those of the two governments.