
Abstract
One of the best ways to improve new learning and increase memory strength is by reprocessing the recently acquired information, for example, by thinking of it again. Synaptic plasticity, the process by which neurons change the strength of their connections with each other, is fundamental for learning and memory formation. Yet, at present, it is unclear how reprocessing information drives synaptic plasticity to support memory improvement. A new study suggests that reprocessing enhances memory formation by recruiting more synapses to represent the new memory, thus increasing its strength.
In this mini-review, I will use fresco painting as a metaphor for memory formation to illustrate the importance of early memory processing and the synaptic changes that support them. Frescos are painted with watercolors over wet plaster. As the plaster dries, it reacts with the colors’ pigments, leading to a durable coloration of the wall or ceiling surface. This time-limited reaction gives an hours-long window for creating most of the fresco, a transience that is also captured in the technique's name (fresco means fresh in Italian). Fresco painting is a useful analogy for memory since both are representations of the world, created over time, and are more amendable to changes and susceptive to disruptions when new (Figure 1).

Early memory formation as a fresco. Akin to memory, a fresco is a representation of reality that takes time to produce and is easier to change or disrupt when new. In this analogy, encoding corresponds to a painting session, which for our purposes, was the only opportunity to see the models. Early consolidation follows, and it matches with the changes made when the subjects were no longer present, but the paste was still wet.
In a recent publication, 1 we studied the neuronal mechanisms that support early stages of memory formation: memory encoding and consolidation. In memory science, encoding refers to several cognitive and neural processes that take place when we acquire new information, specifically at the time we are exposed to the thing-to-be-remembered. In the fresco metaphor, encoding corresponds with the initial stage of the painting session, which, for our purposes, was the only opportunity to see the work's models (Figure 1). Memory consolidation refers to the many processes that stabilize memories; some occur minutes to hours after encoding, while others arise across days to weeks after.2,3 Here, we are concerned only with the early stages of consolidation. In our metaphor, early consolidation, being an umbrella term for many processes, corresponds with all the changes made when the painting's models were no longer present, but the paste was still wet. It is both the hardening of the paste with pigment and the addition of more brush strokes. Early consolidation is a period known to endow greater strength to mental/neuronal processes that increase memory retention, but it is also more sensitive to disruptions, such as from competing information or neuroactive drugs.2–5
Early consolidation is sometimes referred to as “synaptic consolidation,” indicating the centrality of synaptic plasticity to processes occurring over these short time scales.2,3 In short, synaptic plasticity is the change in the strength of neuronal connections.3,6 Since neural representations of the world are brought about through the firing of sets of neurons, which depends on their synaptic connectivity, changes in this connectivity can alter those representations, thereby supporting learning and memory formation. In the fresco analogy, synaptic plasticity could be matched with changing the color of the paste by application of the watercolors.
We used a well-established learning task suited for studying cellular and circuit mechanisms of learning: the classical (Pavlovian) threat conditioning paradigm, which involves the presentation of a neutral stimulus (here, an auditory tone) and an aversive stimulus (an electric foot shock). The thing-to-be-learned here is that the tone is associated with the shock and, therefore, that it merits a defensive response. In memory science terms, the training session corresponds with the encoding phase, after which consolidation ensues. In the most minimalistic terms, encoding corresponds with just the time(s) of tone and shock delivery(ies), and the rest of the training session would be considered as peri-encoding. 7 Finally, recall tests are instances when subjects are presented again with the tone, where the extent of the freezing response indicates memory strength. 8
In one set of experiments, we trained mice with a single pairing and tested freezing responses to the tone at two later time points: first, early on- just a minute and a half after the pairing, still during the training session; and again, a day later (Figure 2(A)). The results showed that the mice froze during both the early and late tone presentations and that freezing was higher a day after training (Figure 2(A)). We argue that during the tone-shock pairing or shortly afterward, the mice formed an initial association between the tone and shock, and this is why they displayed defensive freezing when the tone was presented again. If freezing to the second tone resulted simply from high vigilance facilitated by the shock, 9 we would have seen a similar freezing response after non- associative presentations of the shock and tone, which we did not. 1 Finally, the improvement in freezing is likely because consolidation occurred after encoding.

Early memory reprocessing during threat learning improves long-term retention. (A, B) Classical threat conditioning involves pairing a tone and shock, after which mice display defensive freezing responses to the tone. We found that mice exhibited freezing to the tone already 90 seconds after pairing and that freezing increased a day later (A). In addition, the presentation of the tone shortly after pairing increased long-term memory retention (A, B). Schematics are based on data from Ref. 1
A second set of experiments had a more intriguing result. We found that mice presented with the tone for a second time at training displayed more freezing at the 24-hour test than animals that were not presented with the tone again (Figure 2(A) and (B)). To explain this improvement in freezing, we suggested that when the mice were presented with the tone a second time, it was processed in relation to the shock, hence elaborating it and enhancing the memory's consolidation. 1 This line of explanation agrees with a large amount of data indicating that early memory reprocessing increases retention strength5,10 (but see). 11 A prime example of such a memory-facilitating reprocessing event is “retrieval practice,” where trying to recall recently learned information is known to improve mnemonic performance. 5 Retrieval practice is thought to work through elaborating and contextualizing the new memory and creating additional retrieval routes.
We wanted to visualize the cellular processes taking place during threat memories’ encoding and early consolidation. Therefore, we worked out a method to condition mice while imaging their brains with standard two-photon (2P) microscopy, which provides exceptionally high spatio-temporal resolution of activity within individual neurons, down to the level of their synapses. We applied the same training protocol used in a conditioning box- a single pairing followed by another tone- during 2P imaging. But what should we look at? Ideally, we would like to image a process that is important for threat learning in a relevant brain region. We had an opportunity to do so by imaging synaptic plasticity in the primary motor cortex (a cortical region involved in movement control 12 that is essential for threat learning together with other brain regions)1,8,13,14 (Figure 3(A)). Specifically, we imaged layer 5 pyramidal neurons (L5 PNs), which are excitatory output neurons, and we focused on their synapses (i.e. their dendritic spines, the receiving-end of a synapse) located in cortical layer 1, a compartment of the cortex that is especially important for learning as it preferentially receives higher order information. 15
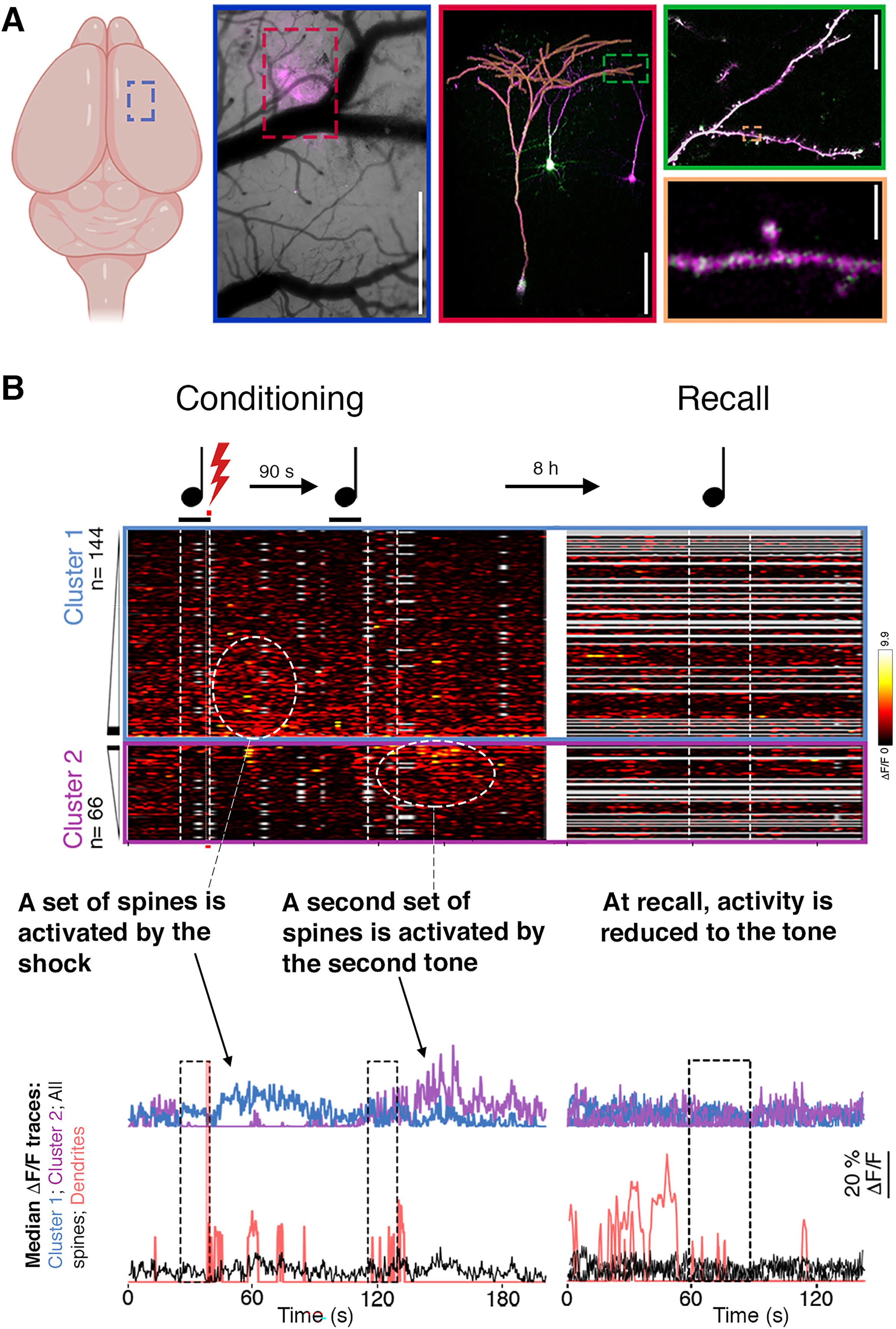
Early memory reprocessing activates an additional set of dendritic spines. (A) Different levels of brain organization. Moving from left to right, the images zoom in on smaller and smaller functional components in the brain- from the brain itself, through a cortical region (framed in blue), a single pyramidal neuron (framed in red), a dendrite (green), and a spine (the receiving-end of a synapse; beige). (B) Heat maps (top) and median traces (bottom) of calcium spine activity during training and recall. Note that the tone-shock pairing induced the activation of a set of synapses and that reprocessing due to the second presentation of the tone activated a second set of synapses. This activation was not evident as early as 1 hour after training. Data in A and B were adapted from Ref. 1
Imaging synaptic activity during threat training provided our most notable results. We found that a set of synapses of L5 PNs was activated immediately after the shock application (Figure 3(B)). However, a second set of synapses, not triggered by the shock, were subsequently activated by the second tone (Figure 3(B)). In contrast, when the tone was presented an hour after training, no activation of additional synapses was detected (Figure 3(B)). Our explanation for these results is as follows: upon the tone-shock pairing, the association of these stimuli was rapidly encoded by the activation of a subset of synapses and the plasticity their activation induced. Once the second tone was presented, after the formation of the initial tone-shock association, the animals were prompted to process the tone again, this time in relation to the shock; this led to the recruitment of the other set of synapses. The fact that additional synapses were activated, instead, for example, reactivating the synapses already triggered by the shock, suggests a synaptic mechanism by which early memory reprocessing elaborates the memory and increases its strength. Finally, that the tone triggered synaptic activation only early during learning (and not an hour later) is in line with the importance of early periods of memory formation for its eventual strength.4,7 To complete the fresco analogy, we could say that early consolidation processes are the ones occurring after the painting's models have left. The recruitment of additional synapses corresponds with adding new brush strokes to the piece, while other consolidatory processes correspond with other reactions of the paste and paint.
Footnotes
Acknowledgments
The author thanks members of the Rudy lab for discussions. R. Machold for comments on the manuscript.
Author contributions
Declaration of conflicting interests
The author declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Institutes of Health (grant number R01NS110079 to B. Rudy).