
Abstract
Atractylodes lancea, commonly known as Kod-Kamao in Thai, a traditional medicinal herb, is being developed for clinical use in cholangiocarcinoma. β-eudesmol and atractylodin are the main active components of this herb which possess most of the pharmacological properties. However, the lack of adequate toxicity data would be a significant hindrance to their further development. The present study investigated the toxic effects of selected concentrations of β-eudesmol and atractylodin in the heart, liver, and endocrine systems of zebrafish embryos. Study endpoints included changes in the expression of genes related to Na/K-ATPase activity in the heart, fatty acid-binding protein 10a and cytochrome P450 family 1 subfamily A member 1 in the liver, and cortisol levels in the endocrine system. Both compounds produced inhibitory effects on the Na/K-ATPase gene expressions in the heart. Both also triggered the biomarkers of liver toxicity. While β-eudesmol did not alter the expression of the cytochrome P450 family 1 subfamily A member 1 gene, atractylodin at high concentrations upregulated the gene, suggesting its potential enzyme-inducing activity in this gene. β-eudesmol, but not atractylodin, showed some stress-reducing properties with suppression of cortisol production.
Introduction
β-eudesmol and atractylodin (Figure 1a and b, respectively) are the major volatile components of the rhizome of Atractylodes lancea (Thunb.) DC. 1 Both are reported to be the main ingredients that exhibit most of the pharmacological activities of the herb.2,3 Studies of β-eudesmol and atractylodin in cholangiocarcinoma (CCA) cell line CL-6 have demonstrated the potent inhibitory activity of both compounds with mean (±SD) IC50 values of 21.5 ± 2.12 and 24.0 ± 1.98 μM, respectively. 3 Cell cycle arrest at the G1 phase and induction of cell apoptosis through activation of caspase 3/7 was considered one of the possible mechanisms by which the compounds exert their inhibitory actions on CCA cells. 4 The crude ethanolic extract of A. lancea induced apoptosis in only 5.5% of CL-6 cells, while β-eudesmol and atractylodin induced apoptosis in 37.84% and 27.21% of the cells, respectively. β-eudesmol and atractylodin had more selective cytotoxic effects against CL-6 cells than normal cells. 5 The IC50 values of β-eudesmol and atractylodin in CL-6 cells were 39.33 ± 1.15 and 41.66 ± 2.51 µg/ml, while those in the human embryonic fibroblast cells (OUMS) were 53.15 ± 3.9 and 64.33 ± 2.08 µg/ml, respectively. 4

Structure of (a) β-eudesmol and (b) atractylodin.
As A. lancea is being developed for clinical use in CCA, 6 evaluation of the toxicity profiles of β-eudesmol and atractylodin are essential to avoid any future objections toward its safety. Different types of toxicity studies need to be carried out to cover as many organs and/or systems of an organism as possible. Complete and in-depth toxicity data could help overcome the hurdles that prevent the formulation from reaching the market, as well as avoid withdrawal of the formulation from the market after it has been registered.7,8 Some of the major toxicity data that lead to the failure of the drug to reach the market and/or withdrawal from the market are cardiotoxicity, hepatotoxicity, and toxic effect on endocrine systems. This study, therefore, aimed to investigate the toxic effects of β-eudesmol and atractylodin on the expressions of the genes related to Na/K-ATPase in the heart, including the fatty acid-binding protein 10a (fabp10a) and cytochrome P450 family 1 subfamily A member 1 (cyp4501a1) in the liver using the zebrafish model. In addition, the effects on cortisol production in zebrafish embryos were also investigated.
Methods
Compounds
β-Eudesmol and atractylodin, in their pure form (99.99%), were bought from Wako Pure Chemical Industries Ltd (Osaka, Japan). Solvents used for dissolving β-eudesmol and atractylodin were ethanol (EtOH) and dimethyl sulfoxide (DMSO), respectively. Preparation of different concentrations of the test compounds was done by diluting the stock solutions in embryo medium (E3). The highest concentrations of EtOH and DMSO in the final test solutions were 1.11% and 0.6%, respectively, not exceeding the lowest permitted concentrations that cause phenotypic and molecular level defects in the zebrafish embryos.9–11
Zebrafish embryo collection
Ethical approval to report this case was obtained from the Ethics Committee for Animal Research of the National Science and Technology Development Agency, Thailand (Number 005/2561). Zebrafish were maintained and handled according to the recommendations provided in OECD Test No. 236: FET. 12 To get the eggs, adult male and female fish were made to mate in spawning tanks an hour before the collection of the eggs. The eggs were then collected and raised in E3 water. Naturally fertilized eggs were selected for the tests using a stereomicroscope (Olympus SZX7, Olympus Corporation Shinjuku Monolith, Tokyo, Japan).
Treatment of embryos with test compounds
Appropriate concentrations of the test compounds were selected based on the preliminary experiment. The concentrations selected for β-eudesmol were 12.5, 50, and 100 µM and for atractylodin were 12.5, 25, and 50 µM. The control consisted of only the solvent in the vehicle. Naturally fertilized eggs aged 0.75–1.25 h post-fertilization (PF) were selected and transferred to a 12-well plate (30 embryos/well). The test solutions, either β-eudesmol (12.5, 50, and 100 µM) or atractylodin (12.5, 25, and 50 µM) and the control were added to respective wells. The plate was then incubated at 28.5°C for 72 h. An equal number of embryos from each concentration was transferred to a centrifuge tube, and the exposure media was completely removed. The embryos were washed three times with type II deionized water for use in further experiments.
RT-qPCR analysis
Total mRNA of the embryos was extracted using the RNeasy Plus Kit (Qiagen, Hilden, Germany) following the company's instructions. One microgram of mRNA was reverse transcribed into cDNA using iScript™ Reverse Transcription Supermix for RT-qPCR (Bio-rad, CA, USA). cDNA (500 ng) was used for RT-qPCR using Bio-Rad CFX96 thermal cycler and iTaq™ Universal SYBR® Green Supermix (Bio-rad). 13 The primer sequences used are presented in Table 1. The reaction process included incubation at 50°C for 2 min, denaturation for 10 min at 95°C, and extension of 40 cycles of 15 s at 95°C, 30 s at 60°C, and 30 s at 72°C. Melt curve analysis was immediately performed, starting at 65°C and increasing to 0.5°C per cycle for 60 cycles. The change in gene expression was determined according to the Livak method. 14
Nucleotide sequences of primers used in the R-qPCR.

ELISA assay of whole-body cortisol
Whole-body cortisol of the embryos was extracted as previously described. 15 In order to obtain a sufficient amount of cortisol, 70 embryos/concentration were used. Cortisol was extracted by adding diethyl ether in embryos homogenized in 1xPBS, and the concentration was determined using a commercial ELISA kit (Fish cortisol ELISA kit, MyBioSource, CA, USA). Samples were prepared in threefold dilutions following optimization tests; each assay was performed in duplicates. Fifty microliters each of the standard and the sample were added to each well, followed by an antibody (1x, 50 μl). The plate was incubated at 37°C for 40 min, and the content in each well was washed with 200 μl of wash buffer. After that, HRP conjugate (1x, 100 μl) was added. The plate was again incubated at 37°C for 30 min. The plate was rewashed, and 90 μl of TMB substrate was added and further incubated at 37°C for 20 min. At last, 50 μl of stop solution was added, and the absorbance was measured at 450 nm in a Spectramax M5 microplate reader (Molecular Devices, California, USA).
Statistical analysis
Statistical analysis was performed using SPSS version 23 (SPSS Statistics 23.0, International Business Machines Corp., Armonk, NY, USA). Data are presented as mean ± SD values of three independent experiments (duplicates each). Comparison of the differences in more than two and two groups of quantitative data was performed using one-way ANOVA followed by Fisher's LSD test The statistical significance level was set at p = 0.05.
Results
Na/k-ATPase genes
The selected nonlethal (12.5 μM) and lethal (50 μM) concentrations of β-eudesmol suppressed the expression of atp1a1a (α1a) gene by 0.22- and 0.18-fold, respectively, compared to the control (p < 0.01 for both). The same concentrations also downregulated the expression of the atp1b1a (β1) gene by 0.24- and 0.18-fold, respectively (p < 0.001 for both). No significant effect on the expression of the atp1a2 (α2) gene was found (Figure 2a). Similarly, atractylodin at nonlethal (12.5 μM) and lethal (25 μM) concentrations downregulated the expression of the atp1a1a (α1a) gene by 0.63- and 0.71-fold, respectively, compared with control (p < 0.01 and p < 0.05, respectively). The selected concentrations suppressed atp1b1a (β1) gene expression by 0.64- and 0.73-fold, respectively (p < 0.01 and p < 0.05, respectively). Atractylodin also appeared to suppress the expression of the atp1a2 (α2) gene at nonlethal concentrations (Figure 2b).

Relative expressions of Na/K-ATPase genes in zebrafish embryos exposed to different concentrations (μM) of the test compounds (a) β-eudesmol and (b) atractylodin for 72 h. Expressions were compared to β-actin as the internal control gene.
Hepatotoxicity genes
β-eudesmol at nonlethal (12.5 μM) and lethal (50 μM) concentrations caused significant downregulation of the fabp10a gene by 0.41- and 0.28-fold, respectively, compared with control (p < 0.001 for both). The selected concentrations of β-eudesmol did not cause any significant change in cyp4501a1 gene expression when compared with control (Figure 3a). For atractylodin, only the lethal concentration (25 μM) resulted in significant downregulation of the fabp10a gene (0.47-fold, p < 0.05), while the nonlethal concentration did not show any significant change. Similarly, only the lethal concentration caused upregulation of the cyp4501a1 gene (1.24-fold, p < 0.05), while the nonlethal concentration did not alter the expression of the gene (Figure 3b).
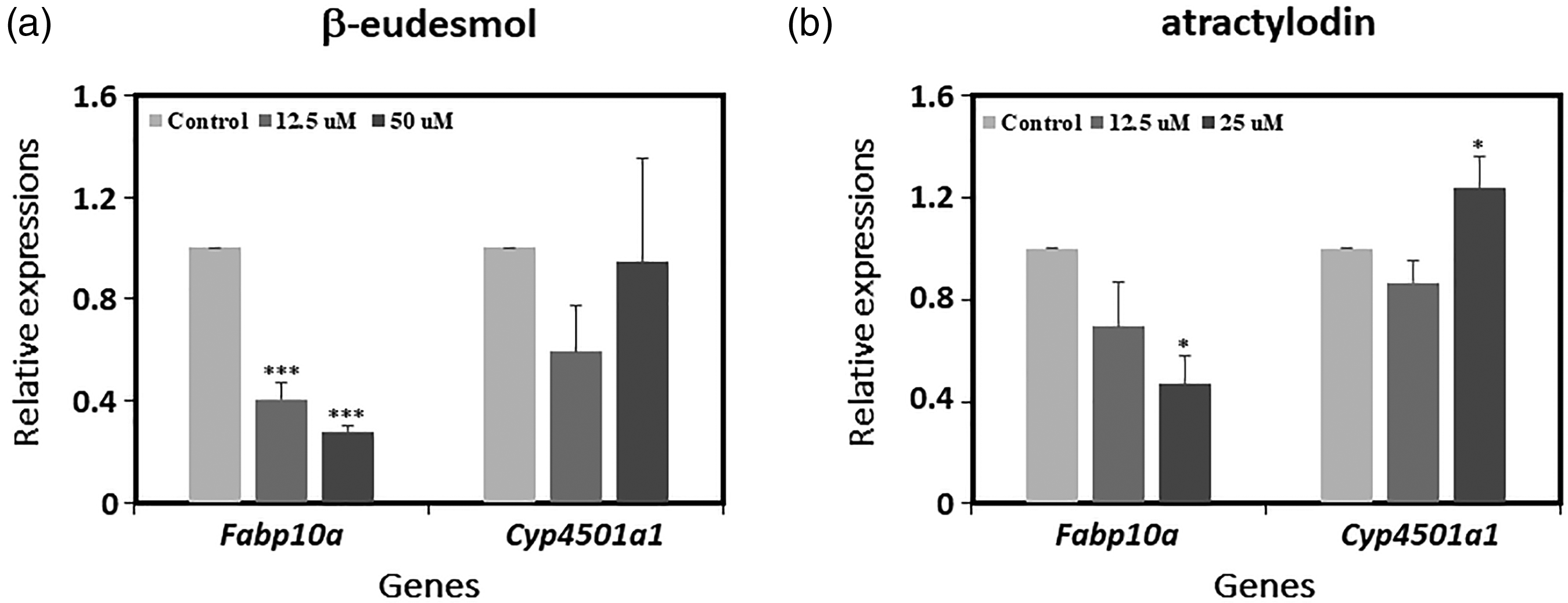
Relative expressions of genes involved in hepatotoxicity in zebrafish embryos exposed to different concentrations (μM) of the test compounds (a) β-eudesmol and (b) atractylodin for 72 h. Expressions were compared to GAPDH as the internal control gene.
Whole-body cortisol levels
Using selected concentrations of β-eudesmol (50 and 100 μM), it was found that only the highest concentration (100 μM) caused significant suppression of whole-body cortisol concentration (8.08 ng/ml) in embryos when compared with control (12.06 ng/ml) (p < 0.05, Figure 4a). Atractylodin at both selected concentrations (25 and 50 μM), however, did not produce any significant alteration in whole-body cortisol concentrations in zebrafish embryos (Figure 4b).

Cortisol levels (ng/ml) obtained from zebrafish embryos exposed to different concentrations of (a) β-eudesmol and (b) atractylodin compared to the control group.
Discussion
Apart from the pharmacological properties, toxicological effects also significantly determine the successful development of a candidate compound for the targeted disease. 16 Even after the drug has been launched for clinical use, some serious safety issues may also lead to the withdrawal of the drug from the markets like in the case of thalidomide.7,8 Therefore, the ideal approach would be to investigate the possible toxic effects of the test compounds/candidate drugs on critical organs and organ systems during the preclinical studies to avoid the occurrence of serious drug reactions in humans.
Cardiotoxicity and hepatotoxicity are the most common causes of withdrawal of candidate drugs during preclinical and clinical development. 16 In this study, the cardiotoxic and hepatotoxic effects, and in addition, the effect on the endocrine system of β-eudesmol and atractylodin were investigated in the zebrafish model. The effects of both compounds on the expression of the Na/K-ATPase-related genes in the heart and some selected biomarker genes in the liver, including the production of whole-body cortisol were evaluated. Na/K-ATPase is a protein oligomer comprising α and β subunits and FXYD proteins. 17 Its function is to generate and maintain Na+ and K+ gradients across plasma membranes. 18 Genes related to Na/K-ATPase activity are expressed in various organs of the zebrafish, such as the heart, brain, spinal cord, and olfactory placode. 19 In the heart, the prominently expressed genes are atp1a1a (α1a), atp1a2 (α2), and atp1b1a (β1). 20 They function to maintain the ion gradient across the heart muscle and also help maintain the left-right symmetry in the heart as well as cardiac patterning. 21 Liu et al., studied the potential toxicity of Palytoxin (produced by marine dinoflagellates) in mice and found out that the compound selectively binds to Na + /K + -ATPase with a Kd of 20 pM, causing death within several minutes due to intense vasoconstriction, increased arterial pressure, and heart failure. 22 A study by Huang et al., investigating the toxicity of carboxin in zebrafish reported that the compound inhibited the activity of the Na + /K + -ATPase, thereby resulting in heart failure. 23 Similar inhibitory effects were observed with the test compounds. Both β-eudesmol and atractylodin were found to significantly downregulate the expressions of α1a and β1 genes. Shu et al. studied the importance of α1 and β1 genes in embryonic heart development in heart and mind (had) mutant zebrafish that lack Na/K-ATPase α1 and β1. The had mutant embryos showed abnormalities in primitive heart tube extension, cardiomyocyte differentiation, and also cardiac function. 24 This suggests that suppression of α1 and β1 genes by β-eudesmol and atractylodin may also lead to similar abnormalities. The extent to which such abnormalities are caused and subsequent effects on the normal functioning of the cardiovascular systems remains to be further investigated in other animal models. Atractylodin also seemed to suppress the expression of the α2 gene. Deficiency of α2Na + /K + -ATPase has been reported to reduce heart rate in zebrafish. 21 It is possible that the inhibitory effect of atractylodin on the heart rate of the embryos, as observed earlier, could also be due to suppression of the α2Na + /K + -ATPase activity. β-Eudesmol, on the other hand, did not have any significant effect on α2 expression. This observation supported the results of our previous study on the absence of the effect of the compound on the heart rates of the embryos. 25
Fabp10a is a member of the liver fatty acid-binding protein family that acts as an antioxidant during oxidative stress.26,27 It serves as a biomarker for liver toxicity and its impairment could influence liver disease pathogenesis.28,29 The deficiency of fabp10a in zebrafish embryos led to increased oxidative stress and inflammation-mediated hepatotoxicity. 30 A study by Liu et al., found that mitoxantrone significantly decreases the expression of the fabp10a in zebrafish livers inducing hepatotoxicity. 31 In this study, β-eudesmol at all concentrations significantly reduced the expression of fabp10a gene. For atractylodin, only the highest concentration caused significant inhibition of the fabp10a gene expression. Results suggest the toxic effect of β-eudesmol on the developing liver of the embryos through inhibition of fabp10a gene, while atractylodin has such effect only at the highest concentration. Whether these toxic effects would lead to observable defects and long-term effects on developing hearts should be further investigated.
Cyp1a1 is a member of the cyp450 heme monooxygenases enzyme family. 32 Cyp enzymes are responsible for the detoxification of xenobiotics as well as endogenous substrates.33,34 Zebrafish cyp1a1 has been reported to be prominently expressed in the liver, 35 while other members like cyp1b1, cyp1c1, cyp1c2, and cyp1d1 are found in small quantities in the liver, gill, and kidney.35,36 Exposure to external substances can have varying effects on the production of cyp enzymes. Some xenobiotics tend to increase the production of enzymes (enzyme inducers), while others might suppress enzyme production (enzyme inhibitors). Enzyme inducers can accelerate the metabolism of other compounds, while enzyme inhibitors can delay the metabolism. 37 β-Eudesmol showed a nonsignificant increase in the expression of the cyp1a1 gene with increasing concentrations. Atractylodin at the highest concentration significantly increased the expression of the gene. Hence, it might be concluded that β-eudesmol does not influence the cyp1a1 enzyme, while atractylodin at higher concentrations tends to act as a cyp1a1 enzyme inducer, which might affect the rate of metabolism of the substrates that are metabolized by this enzyme.
The stress hormone cortisol is the major corticosteroid in most mammals and fishes, which is produced in the hypothalamic–pituitary–interrenal (HPI) axis. Its production is mostly triggered by various stressful conditions such as confinement, handling, the sight of predators, and exposure to extremes of temperatures and toxicants. 38 Zebrafish's response to stress by the increase in the production of cortisol has been confirmed in several studies. Zebrafish exposed to a sublethal concentration of ammonia (equivalent to the amount of ammonia found in the environment) for 24 h led to more than a threefold increase in the whole-body cortisol levels when compared with the control group. The response to stress through an increase in cortisol production was maintained as long as the inducer was present, whereby the cortisol levels were about twofold higher than the control group even after 14 days of exposure. 39 Cortisol levels can also be reduced when the fish are exposed to certain compounds with stress-reducing properties. For instance, zebrafish treated with essential oil from Aloysia triphylla had significantly lower plasma cortisol levels when compared with the control group. 40 In another study, treatment of zebrafish with Hydrangeae Dulcis folium significantly prevented the increment in cortisol levels due to net handling stress. 41 Such a trend of reduction of cortisol production was observed in the present study with both test compounds, particularly β-eudesmol. With increasing concentrations of β-eudesmol, whole-body cortisol levels produced by the treated embryos were significantly decreased. This result suggests the stress-reducing activity of β-eudesmol. This finding was in agreement with the reported lower saliva cortisol levels in human subjects who received β-eudesmol prior to a Trier social stress test (TSST) in a previous study. 42 Atractylodin did not have any significant effects on the whole-body cortisol levels, although the amount appeared to decrease with increasing concentrations.
There are earlier studies that reported the toxicity profile of the extract of A. lancea but not of β-eudesmol and atractylodin directly. Plengsuriyakarn et al. reported that rats and mice, when fed with up to 5000 mg/kg body weight of A. lancea extract, produced gastric stimulation and central nervous system inhibition symptoms (diminished response to touch and balance and decreased alertness and motor ability). 43 A study by Koonrungsesomboon et al., on acute and subacute toxicity of A. lancea extract, observed that rats and mice well-tolerated a wide range of dose levels (1000–5000 mg/kg body weight) of the extract. 2
Conclusions
This study demonstrated the effects of β-eudesmol and atractylodin on some of the critical organ systems in developing zebrafish embryos. Results revealed some adverse effects on the functions of the Na/K-ATPase-related genes, which could subsequently affect the functioning of the heart. Similar adverse effects were also observed in the liver, whereby the biomarkers for liver toxicity were triggered. Nevertheless, some constructive effects were also observed, particularly with β-eudesmol, with regard to the reduction of whole-body cortisol production, which would be useful as an approach for further development of the compound for stress relief.
Limitations of the study
This study focused only on a few target genes that were earlier reported to be markers of toxicity of the particular organ system. As such, it may not be representative of the overall toxicity study of the systems discussed here. Other targets at different levels might have to be further studied. However, our study indeed confirms the effects of the test compounds on the selected targets in various organ systems as per our experiments by comparing them with earlier studies.
Footnotes
Acknowledgements
The authors would like to thank Nano Safety and Risk Assessment Laboratory, National Nanotechnology Center, and National Science and Technology Development Agency (NSTDA), Pathum Thani, for their kind support of laboratory facilities.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research and/or authorship of this article: This study was supported by Chulabhorn International College of Medicine (CICM) of Thammasat University, Center of Excellence in Pharmacology and Molecular Biology of Malaria and Cholangiocarcinoma of Thammasat University, National Research Council of Thailand (NRCT), National Research University Project of Thailand (NRU), Office of Higher Education Commission of Thailand, and National Nanotechnology Center (NANOTEC). Kesara Na-Bangchang is supported by the National Research Council of Thailand under the Research Team Promotion grant (grant number NRCT 820/2563). Mr Gyem Tshering is supported by Thammasat University for PhD program and also received financial support from the research fund of Chulabhorn International College of Medicine contract No. T1/2562.
Ethical approval
Ethical approval to report this case was obtained from the Ethics Committee for Animal Research of the National Science and Technology Development Agency, Thailand (Number 005/2561).
Author biographies
Gyem Tshering is a PhD graduate in Bioclinical Sciences in the field of Pharmacology and Toxicology. He has a background in Pharm D. He has three international publications published as the first author.
Wittaya Pimtong is a senior researcher at the Nano Environmental and Health Safety Research, National Nanotechnology Center, National Science and Technology Development Agency of Thailand. He is an expert in Developmental Biology, Molecular Biology, Biochemistry and Toxicology. He has around 20 international publications.
Tullayakorn Plengsuriyakarn is an assistant professor at the Graduate Studies at Chulabhorn International College of Medicine, Thammasat University. He is an expert in Pharmacology (Animal Research). He has more than 30 international publications.
Kesara Na-Bangchang is a professor in pharmacology and an expert in pharmacology and toxicology, especially pharmacokinetics. She has more than 200 international publications. Recently, she won the 2021 National Outstanding Researcher in Chemical and Pharmaceutical Science of Thailand. Currently, she serves as the director of Graduate Studies at Chulabhorn International College of Medicine, Thammasat University.