
Abstract
The objective of this study was to elucidate the characteristics of visually perceived momentum of others’ motions. Twenty participants watched and compared two consecutive point-light running motions: one at a fixed velocity of 8.0 km/h and the other at one of seven velocities (5.6, 6.4, 7.2, 8.0, 8.8, 9.6, and 10.4 km/h). They then evaluated which had greater momentum, or if they were the same. The results indicated that as the velocity deviated from the standard velocity of 8.0 km/h, the correct rates increased; in particular, the correct rate at 5.6 km/h, which differed the most from the standard velocity, was the greatest (p < .001). Additionally, the mean response times at 5.6 and 6.4 km/h, which were relatively smaller than the standard velocity, were significantly lower (both p < .05). This study indicates that humans can accurately perceive the momentum of others’ motions, consistent with previous studies demonstrating an accurate perception of the physical and mechanical properties of human-like motion.
Introduction
In everyday life and sports, we recognize the motions of other people and objects and respond accordingly. We estimate the impact that other people and objects have on us because we often physically and mechanically interact with them. While there are ample studies on how we perceive the physical and mechanical properties of objects, limited attention has been given to how we perceive them in human motion.
Many studies for intuitive physics demonstrate how humans visually perceive the physical and mechanical properties of object and its motion (e.g., Deeb et al., 2021; Kubricht et al., 2017; Neupärtl et al., 2020, 2021; Schwettmann et al., 2019; Vicovaro, 2014, 2021, 2023; Vicovaro & Burigana, 2016; Vicovaro et al., 2019, 2023). Previous review articles (Vicovaro, 2021, 2023) overviewed some studies on intuitive physics while introducing different intuitive physics tasks. For example, when presenting to participants the simulated spheres made of wooden (heavy) or polystyrene (light) falling to the ground and asking them to rate the naturalness of the motion, they consider higher values of acceleration or velocity of motion as natural for the wooden sphere and lower values for the polystyrene sphere (Vicovaro et al., 2019). Humans tend to perceive heavier objects as falling faster than lighter objects, that is, they overestimate the influence of mass on the speed of objects falling downward (for reviews, see Vicovaro, 2021, 2023). While several studies have been conducted on intuitive physics based on objects and their motion, the theoretical mechanism has not been identified yet.
Compared to the plethora of studies on objects and their motion, the number of studies that have investigated how humans visually perceive the physical and mechanical properties of human and human-like motions is relatively limited (Huber et al., 2019; Smeeton et al., 2023; Ueda et al., 2018; Vicovaro et al., 2014; West Jr. et al., 2022). Several important studies have provided significant insights into the perceptual characteristics of physical and mechanical properties of human motion. For example, Huber et al. (2019) asked participants to view multi-joint arm motions and rate the arm stiffness and demonstrated humans’ adeptness in visually discerning the joint stiffness of multi-joint arm motions. Also, Ueda et al. (2018) showed greater sensitivity to speed discrimination of various biological motions, such as walking and kicking when the stimuli presented were more human-like figures. These studies suggest the significance of human/human-like figures to make humans perceive the physical and mechanical properties of human motion. Despite the scarcity of studies on the perception of physical and mechanical properties in human and human-like motions compared to object motion, understanding how humans perceive such properties holds significance for daily life interactions and sports engagements.
In this study, we aimed to elucidate the characteristics of visually perceived momentum of others’ motions by focusing on momentum, a key physical characteristic of human motion. Momentum, the product of velocity and mass, holds particular significance in sports, especially those involving contact with a ball or other individuals (Baker & Newton, 2008; Mann et al., 2022; Roane, 2011). To our knowledge, there are no previous studies examining the perception of momentum in human motion. Therefore, in this study, we investigated the characteristics of the visually perceived momentum of human motion for the first time by manipulating the running velocity component in running motion, one of the basic human movements.
Running motion in this study was presented as biological motion (point-light motion) as in Ueda et al. (2018), who systematically investigated the speed perception of biological motions in various actions, such as walking and kicking. In the sports domain, numerous studies have explored biological motion perception and anticipation based on it (e.g., Abernethy et al., 2008; Abernethy & Zawi, 2007; DeCouto et al., 2024; Higueras-Herbada et al., 2017; Huys et al., 2009; Smeeton et al., 2019). Previous studies have utilized various biological motions from different sports, such as badminton strokes and tennis shots, often comparing experts and novices regarding their ability to anticipate the direction of these motions. This study is the first to investigate the perceived momentum of running biological motion, a common and essential motion across various sports. Unlike Ueda et al. (2018), where participants were required to discriminate the speeds of stimuli, our study asked participants to evaluate the perceived momentums of running motions. Momentum is a physical quantity that represents the perception of the unstoppability of a moving object, which is different from the mere perception of velocity/speed, and the change rate of momentum over time indicates force. Therefore, the greater the momentum (i.e., the greater the mass and velocity), the greater the force required to change the momentum (Roane, 2011), which the force required for such a change in momentum could be involved in the evaluation of the momentum. In summary, the objective of this study was to elucidate the characteristics of visually perceived momentum in human motion, making it the first to explore this phenomenon.
Methods
Participants
Twenty adult males participated in the experiment. All participants had normal vision, with a visual acuity of 0.8 or higher, as measured by the AS-4Fα vision tester (Kowa Company Ltd., Nagoya, Japan), and were right-handed, as assessed by the Edinburgh Handedness Inventory (Oldfield, 1971). In addition, all participants did not have orthopedic hand disorders that could hinder task performance and had no history of neurological disorders. Before commencing the experiment, all participants were informed about the experimental procedure, and all participants gave their written informed consent. This study was approved by the Ethics Committee of Nihon Fukushi University in accordance with the Declaration of Helsinki.
Stimuli
In the experimental task, all participants watched and compared two consecutive running motions displayed on a monitor (LCD-GC271UXB, IO DATA, Kanazawa, Japan). The stimuli were point-light running motions, generated from motion capture data, with the front of the body facing left from the participants’ perspective. Point-light motion as a biological motion is often used for investigating the perception of motion stimuli (e.g., Chang et al., 2021; Chang & Troje, 2009a, 2009b; Pavlidou et al., 2022; Ueda et al., 2018). The model for the stimuli was a male aged 25 years, with a height of 168.2 cm and body mass of 61.0 kg. Additionally, the model had been practicing athletics for 13 years. We captured running motion data at seven different velocities (5.6, 6.4, 7.2, 8.0, 8.8, 9.6, and 10.4 km/h) on the treadmill with a motion capture system (OptiTrack, NaturalPoint, Inc., Corvallis, USA) at a sampling frequency of 100 Hz. Infrared reflective markers (1.4 cm in diameter) were attached to the 20 points of the body as follows: right and left front head, right and left back head, middle-upper edge of sternum, cervical spine (C7), right and left acromion, right and left lateral epicondyle of humerus, right and left of the middle points between radial and ulnar styloid processes, right and left greater trochanter, the middle point between the right and left anterior superior iliac spine, the middle point between the right and left posterior superior iliac spine, right and left lateral points of the knee joint, and right and left lateral ankles.
Experimental Design
All participants sat on a chair in front of a monitor with the head fixed on a chin rest (SR-HDR, SR Research Ltd., Ottawa, Canada). In each trial, they watched the fixation cross for 1 s, followed by the instruction of subsequent display of the first stimulus for 1 s and a movie of running motion at 8.0 km/h as a standard stimulus for one cycle. Then, the instruction of subsequent display of a second stimulus for 1 s and a movie of running motion at one of seven different velocities as a comparative stimulus for one cycle were displayed, followed by a display of the instruction asking the participants to indicate whether motion momentum was greater between the standard and comparative stimuli or the same between them. According to the instruction, the participants promptly pressed one of three keys of the keyboard (PERIBOARD-409, Perixx Computer GmbH, Dusseldorf, Germany) based on the evaluation of momentum. If the participants evaluated the momentum of the first movie as greater than that of the second movie, they pressed the key ‘1’. In the opposite case, they pressed the key ‘2’. If the participants evaluated the same momentum between them, they pressed the ‘ = ’ key. After pressing each of the keys, the next trial started. All participants completed 13 blocks (including the first practice block). Each block consisted of 21 trials, with three trials per comparative stimulus velocity. A short break was set between blocks. The display of stimuli and acquisition of responses were all controlled in MATLAB (Mathworks, Inc., Natick, USA) using Psychtoolbox (Brainard, 1997; Kleiner et al., 2007; Pelli, 1997).
Data Analysis
We computed the mean response rate of how the comparative stimulus was evaluated compared to standard stimulus (i.e., smaller, equal to, or larger), the mean correct rate and mean response time within 12 blocks at each comparative stimulus velocity for each participant. In the mean response time, we used the data on correct trials for the computation. We conducted a normality test and found non-normality in the data. However, following the procedures suggested by Blanca et al. (2024), a one-way repeated measures ANOVA was performed through SPSS software version 27.0 (IBM Corp., New York, USA) by setting these measures as the dependent variables and setting the motion velocity as an independent variable. If we found a significant main effect of the motion velocity factor, we conducted a post-hoc analysis applying the Bonferroni correction to compare stimulus velocities. If the assumption of the sphericity was violated, the Greenhouse-Geisser correction was applied. We set the significance level at .05.
Results
Participants’ Characteristics
Participants’ Characteristics.

Response Rate
Figure 1 shows the mean response rate at which the momentum of the comparative stimulus was evaluated as smaller, equal to, or larger than that of the standard stimulus at each velocity. ‘Small’ in Figure 1 represents the percentage of responses where participants evaluated the momentum of the comparative stimulus as smaller than the standard stimulus at each comparative stimulus velocity, increasing as velocity decreased below the standard stimulus. ‘Equal’ means that participants evaluated the momentums of the comparative and standard stimuli as equal, with the highest percentage occurring when the velocity of the comparative stimulus was 8.0 km/h. ‘Large’ represents the percentage of responses where participants evaluated the momentum of the comparative stimulus as larger than the standard stimulus, increasing as velocity rose above the standard stimulus. The ANOVA for the mean correct response rates at each stimulus velocity revealed a significant main effect (F(2.995, 56.899) = 41.400, p < .001, η
p
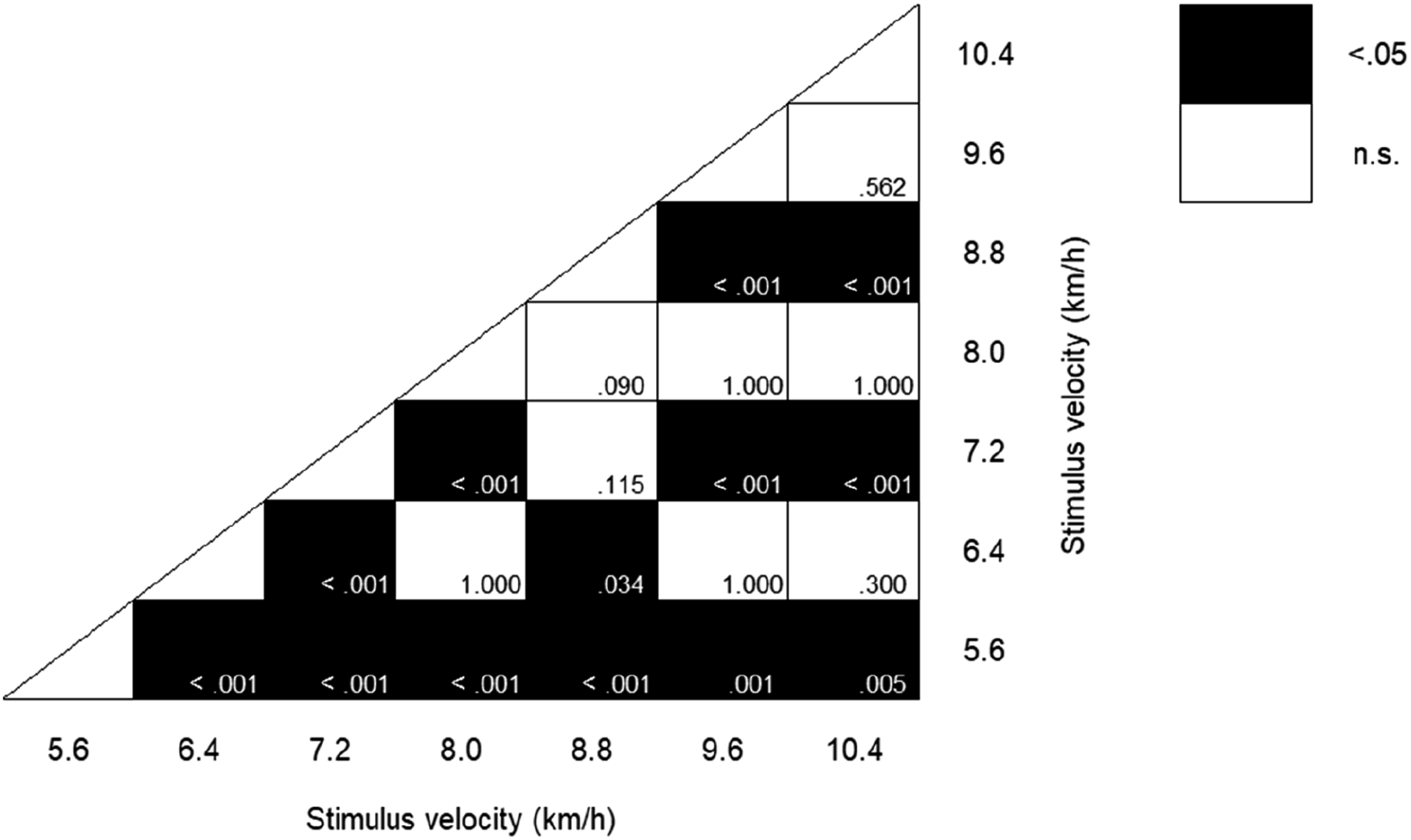
2 = .685), although the Greenhouse-Geisser correction was applied because the assumption of sphericity was violated. Figure 2 shows statistically significant differences with p-values for multiple comparisons between conditions. The correct rate at 5.6 km/h was significantly higher than all other velocity conditions (all p < .01). The correct rate at 6.4 km/h was higher than that at 7.2 and 8.8 km/h (both p < .05). The correct rate at 7.2 km/h was significantly lower than all other velocity conditions (all p < .001), except for 8.8 km/h. A trend was noted where the correct rate at 8.0 km/h was higher than that at 8.8 km/h (p < .1). The correct rate at 8.8 km/h was significantly lower than at all other velocity conditions (both at 9.6 and 10.4 km/h, p < .001). Response rate. Statistical significance with p-values in multiple comparisons for correct rate between stimulus velocities.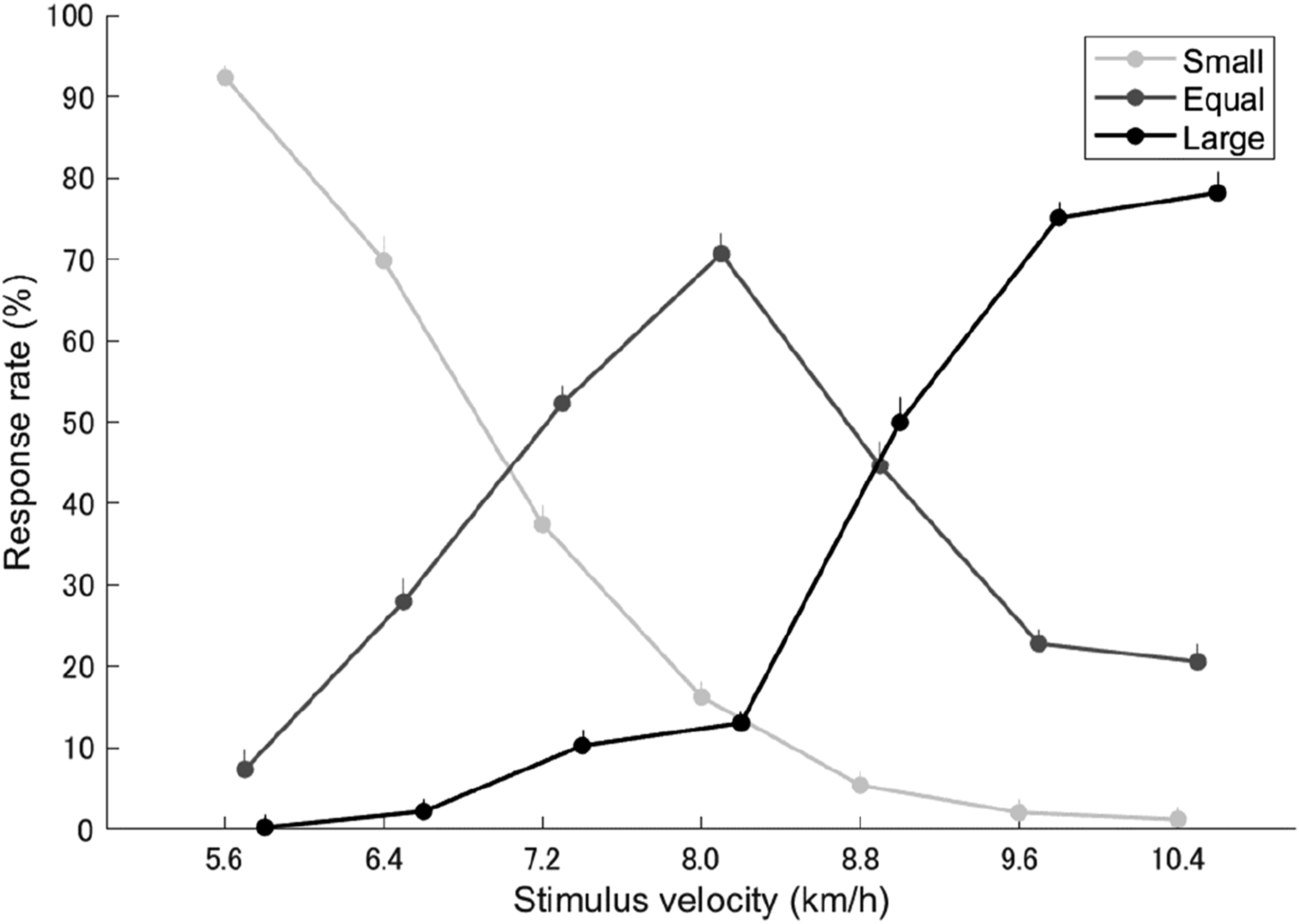
In addition, we computed the incorrect response rates in which participants evaluated the comparative stimuli (5.6, 6.4, 7.2, and 8.0 km/h) as greater than the standard stimulus of 8.0 km/h (i.e., the proportions of overestimations). Similarly, we computed those in which participants evaluated the comparative stimuli (8.0, 8.8, 9.6, and 10.4 km/h) as smaller than the standard stimulus (i.e., the proportions of underestimations) to further clarify their incorrect response directions. A one-way repeated measures of ANOVA revealed a significant main effect of comparative stimulus velocity (F(3.301, 62.710) = 48.730, p < .001, η p 2 = .719). Post hoc tests showed that the lowest incorrect response rate occurred at 5.6 km/h, although it did not significantly differ from the rates at 8.0 km/h (see details in Supplemental Figures 1 and 2).
Response Time
As for the response time, we found a significant main effect (F(3.354, 63.726) = 10.036, p < .001, η
p
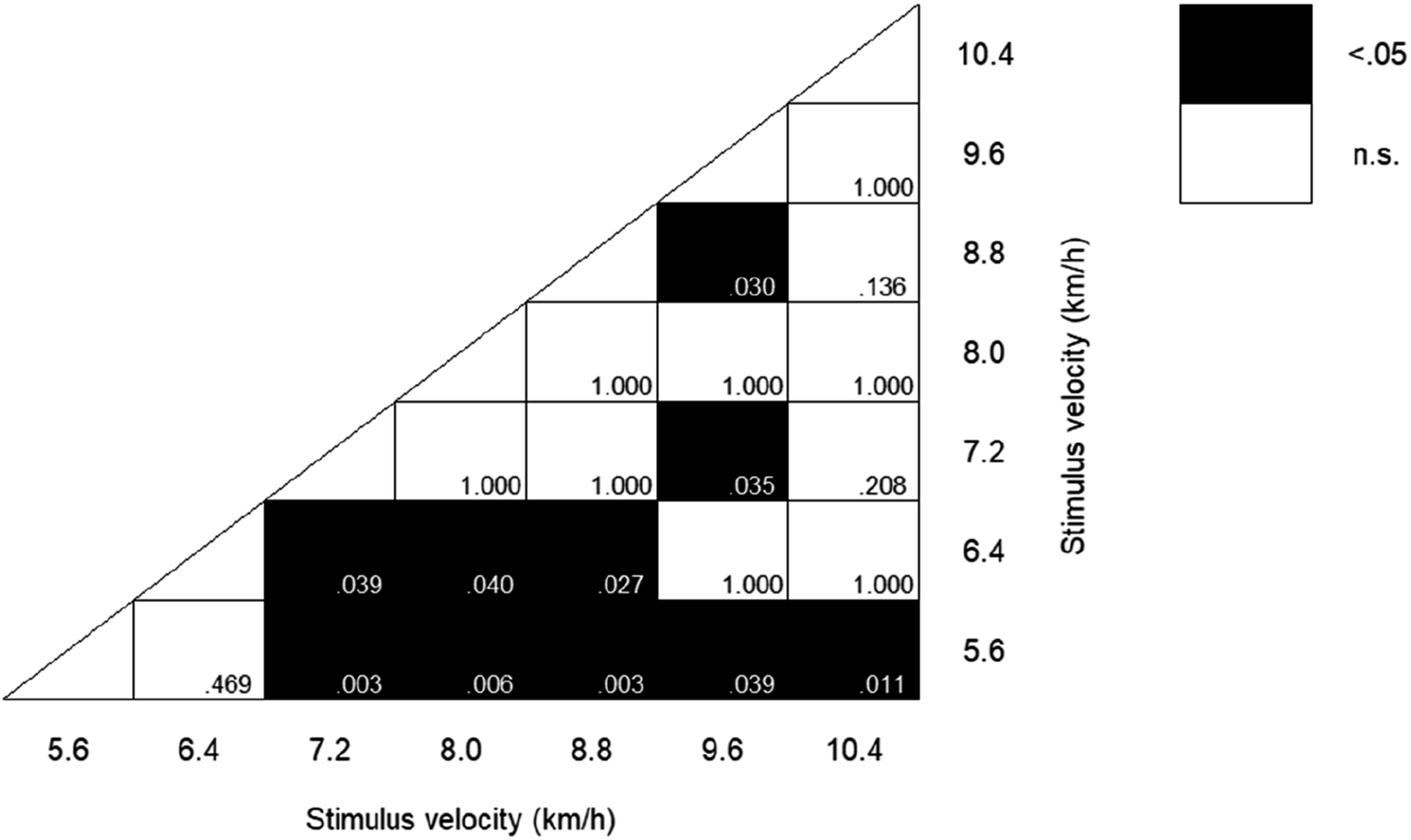
2 = .346, Figure 3), although the Greenhouse-Geisser correction was applied because the assumption of sphericity was violated. Therefore, we conducted multiple comparisons. Figure 4 illustrates statistically significant differences with p-values for multiple comparisons between conditions. The response time at 5.6 km/h was significantly shorter than for all other velocity conditions (all p < .05) except for 6.4 km/h. The response time at 6.4 km/h was shorter than that at 7.2, 8.0 and 8.8 km/h (all p < .05). The response time at 7.2 km/h was significantly longer than that at 9.6 km/h (p < .05). The response time at 8.8 km/h was significantly longer than that at 9.6 km/h (p < .05), similar to 5.6 and 6.4 km/h. Response time. Statistical significance with p-values in multiple comparisons for response time between stimulus velocities.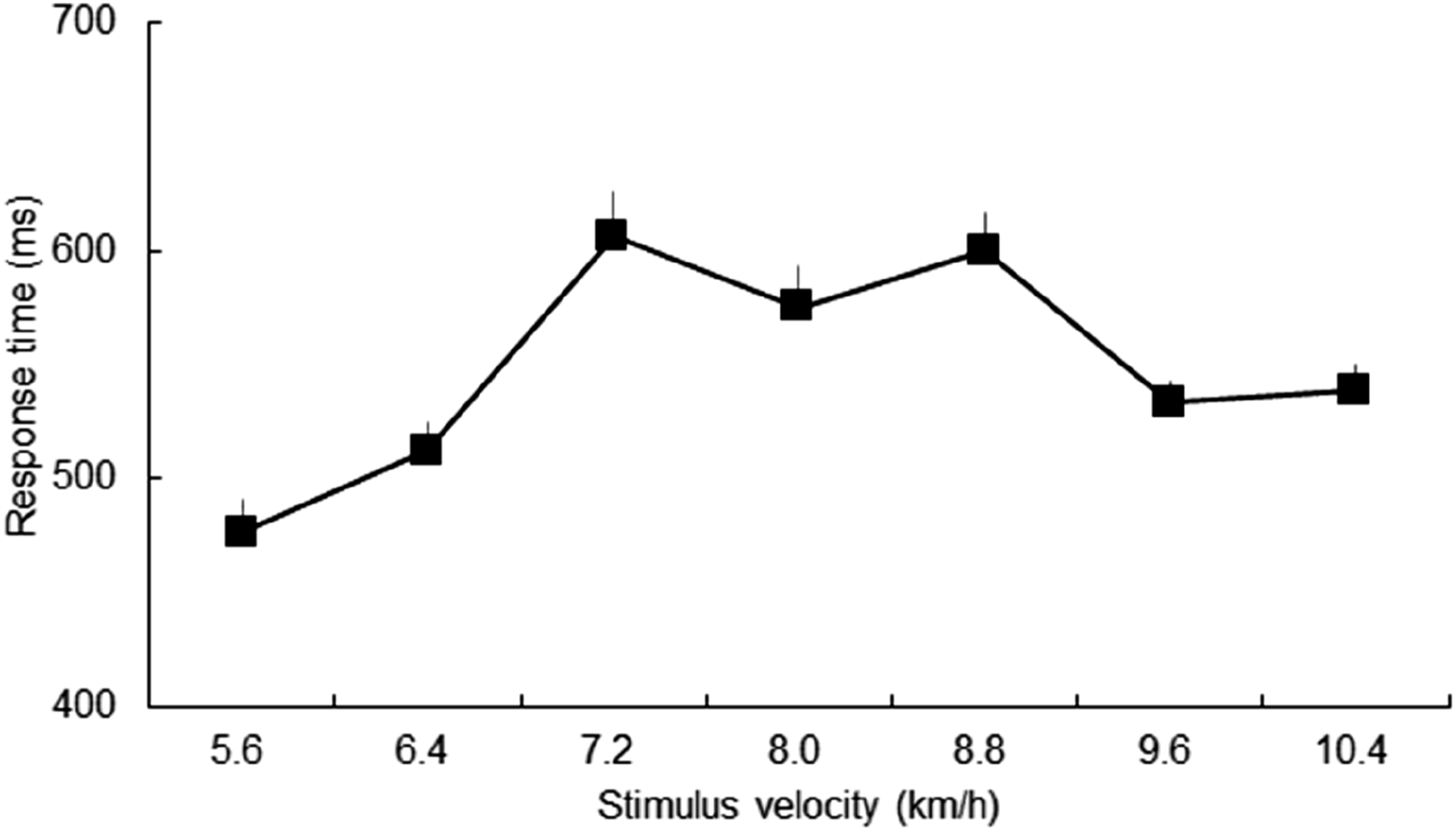
Discussion
In this study, participants observed two consecutive running motions and evaluated the momentum via three choices: pressing key ‘1’ for greater momentum in the first motion (standard stimulus), key ‘2’ for greater momentum in the second motion (comparative stimulus), or key ‘ = ’ for equal momentum. The results indicated that the correct rates for velocities of 7.2 and 8.8 km/h were lower than the correct rate of standard velocity (8.0 km/h). As the velocity deviated further from the standard velocity, the correct rates increased; in particular, the correct rate of 5.6 km/h, which differed the most from the standard velocity, was the greatest. The mean response times at lower velocities (5.6 and 6.4 km/h) were shorter than the standard stimulus, whereas those at higher velocities (8.8, 9.6, and 10.4 km/h) showed no significant difference.
We found that the correct rates increased as the velocity difference compared to the standard velocity increased, with the highest correct rate at 5.6 km/h, which was the slowest in this study. These results suggest that the smaller the velocity difference from the standard velocity, the more difficult it is to evaluate the momentum. Moreover, even with equal velocity differences (e.g., 2.4 km/h at 5.6 km/h and 10.4 km/h), lower velocities (e.g., 5.6 km/h) were easier to evaluate than higher velocities (e.g., 10.4 km/h). This result could be attributed to Weber-Fechner law. According to the law, participants could sensitively perceive the differences in momentum between the standard and comparative stimuli at lower velocities than higher velocities. An alternative explanation is that the result might be attributed to the familiarity with the velocity of human motion. The velocity of 5.6 km/h in this study is close to that of walking (fast walking), which can be observed in daily lives; therefore, familiarization with the low velocity of human motion may improve the correct rate of perception of human motion momentum in this study. We also found that the correct rates at 7.2 and 8.8 km/h were relatively low. This suggests that a ±10% difference from the standard stimulus velocity served as the discrimination threshold in this study. Interestingly, error trials revealed that participants often mistook velocities of 7.2 and 8.8 km/h for 8.0 km/h, suggesting difficulty in discriminating between these velocities.
We observed significant differences in response times between 8.0 and 5.6 km/h and 8.0 and 6.4 km/h. Additionally, there were significant differences between 5.6 km/h and all other velocities except 6.4 km/h. These results suggest that lower velocities (5.6 and 6.4 km/h) decrease the response time. In particular, the velocity of 5.6 km/h exhibited the highest correct rate and quickest response time, suggesting that it was the most easily assessed by participants in evaluating running motion momentum. As mentioned above, these results could be attributed to Weber-Fechner law and/or the familiarity with the lower velocity of human motion.
Previous studies have demonstrated that humans can accurately estimate mechanical properties such as joint stiffness from observed kinematics (Huber et al., 2019; West Jr. et al., 2022). These findings suggest that mechanical and physical properties of human motion influence the observer’s perception and cognition. This study focused on the perceived momentum of biological running motion, expressed by the product of mass and velocity. Ueda et al. (2018) systematically investigated the speed perception of biological motions of various actions, such as walking and kicking. However, the speed/velocity is just the rate of change of the position of a body part over time, while the momentum is a physical quantity multiplied by the mass in addition to the velocity. Furthermore, the momentum represents the perception of the unstoppability of a moving object. Therefore, asking participants to determine the momentum does not correspond to asking them to evaluate which stimulus is faster, although it would be worthwhile to quantitatively examine the differences between velocity perception and momentum perception in future research. The results of our study align with previous findings revealing the influence of mechanical and physical properties such as joint stiffness and speed on the humans’ perception and cognition, providing evidence of the characteristics of the perceived momentum of human motion.
While there are limited studies on the perception of physical and mechanical characteristics of human motion, numerous studies, often discussed in the context of intuitive physics, have been conducted on the perception of these characteristics for object motion. This study on the perception of physical and mechanical characteristics of human motion would contribute to the development of limited previous facts and suggests a new perspective that might align human motion with object motions in terms of intuitive physics. Indeed, from the perspective of the explainability of our results from Weber-Fechner law and the familiarization, this study would have a small-step contribution to existing findings. However, this study focused on one of the significant physical quantity, momentum, would advance the knowledge on the visual perception of human motion in terms of physical and mechanical properties and may contribute to the future development of intuitive physics.
As for the neural mechanisms of the visual perception of the momentum of human motion, to our knowledge, no study has investigated it so far. However, many studies explored the brain regions activated when participants observed human motions, revealing the important role of the superior temporal sulcus (STS) (Beauchamp et al., 2002; Blake & Shiffrar, 2007; Bracci & Op de Beeck, 2023). Previous studies on the speed perception of non-human motion stimuli have indicated the relevance of V5/MT+ (Chawla et al., 1998; McKeefry et al., 2008), a mechanism that might be involved in the perception of momentum as well. In terms of the neural mechanisms of intuitive physics, Schwettmann et al. (2019) investigated the neural representation on object mass and revealed that it is represented in the premotor and parietal brain regions. Karakose-Akbiyik et al. (2024) studied the neural activity associated with observing animated actions by agents in comparison to similar movements of inanimate objects and discovered that the left dorsal premotor cortex and superior parietal lobules were more sensitive to information about inanimate object events that were solely shaped by the physics of the scene. Based on this evidence, one may speculate that STS, V5/MT+, and the premotor and parietal regions might be activated when humans observe the running biological motion and evaluate its momentum. Future studies on the neural mechanisms during the evaluation of momentum are needed to confirm it.
This study investigated the characteristics of visually perceived momentum by manipulating the velocity component of momentum, which is the product of velocity and mass in human motion. However, one of limitations was that the velocities set in this study ranged from 5.6 to 10.4 km/h, which were considerably slower than those for sprinting. Future studies are needed to determine whether the findings of this study can be replicated in additional velocity ranges. In addition, another limitation was that the mass component was not manipulated in this study. Mass is an important component that defines momentum, and its influence on the perception of momentum by manipulating human figures needs to be investigated. Furthermore, it would be worthwhile to quantitatively examine the differences between velocity perception and momentum perception in future research. In clinical and sports applications, the findings of this study could contribute to the development of interventions aimed at improving the abilities of clinicians and sports coaches to accurately evaluate the motions of patients and athletes, potentially benefiting both rehabilitation and athletic performance enhancement.
Conclusions
In this study, we investigated human perception of momentum by manipulating the velocity component. The results revealed a heightened accuracy as the velocity deviated further from the standard, particularly notable with the slowest velocity, which exhibited the most significant difference from the standard velocity. However, future studies are needed to determine whether the findings of this study can be replicated across different velocity ranges, to investigate the influence of mass on the perception of momentum, and to explore the neural mechanisms underlying the perception of momentum.
Supplemental Material
Supplemental Material - Visually-Perceived Momentum in Biological Motion: The Influence of Changes in Running Velocity
Supplemental Material for Visually-Perceived Momentum in Biological Motion: The Influence of Changes in Running Velocity by Shinji Yamamoto and Masanobu Araki in Perceptual and Motor Skills
Footnotes
Author Contributions
Shinji Yamamoto: Conceptualization, Methodology, Software, Formal analysis, Investigation, Writing, Visualization, Supervision, Project administration, Funding acquisition.
Masanobu Araki: Conceptualization, Methodology, Formal analysis, Writing, Visualization.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by JSPS KAKENHI Grant Number JP20K19525.
Ethical Statement
Data Availability Statement
The data will be made available upon request.
Supplemental Material
Supplemental material for this article is available online.
Author Biographies
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.