
Abstract
Attention to soil biodiversity and its importance for sustainable food production has markedly increased in recent years. In particular, the loss of soil biodiversity as a consequence of intensive agriculture, land degradation and climate change has raised concerns due to the expected negative impacts on ecosystem services, food security and human health. The result is a strong demand for ‘nature-based’ practices that stimulate soil biodiversity or beneficial soil organisms and enhance soil health. Here, we examine the origin of popular ideas on the role of soil biology in sustainable soil management, as well as their potential to address key global challenges related to agriculture. Three examples of such ideas are discussed: 1) a higher fungal:bacterial (F:B) biomass ratio favours soil carbon storage and nutrient conservation; (2) intensive agricultural practices lead to a decline in soil biodiversity with detrimental consequences for sustainable food production; (3) inoculation with arbuscular mycorrhizal fungi reduces agriculture's dependency on synthetic fertilizers. Our analysis demonstrates how ecological theories, especially E.P. Odum's ( 1969) hypotheses on ecological succession, have inspired the promotion of agricultural practices and commercial products that are based on the mimicry of (soil biology in) natural ecosystems. Yet our reading of the scientific literature shows that popular claims on the importance of high F:B ratios, soil biodiversity and the inoculation with beneficial microbes for soil health and sustainable agricultural production cannot be generalized and require careful consideration of limitations and possible trade-offs. We argue that dichotomies and pitfalls associated with the normative use of nature as a metaphor for sustainability can be counterproductive given the urgency to achieve real solutions that sustain food production and natural resources. Finally, implications for soil ecology research and sustainable soil management in agriculture are discussed.
Keywords
Introduction
Persistent global issues, including the negative impacts of modern agriculture on soil and water resources, biodiversity, the global climate and the resilience of food production, call for urgent actions to transform the ways in which we manage land. In particular, widespread soil degradation due to unsustainable agricultural practices and intensification of agriculture has been associated with the decline of soil biodiversity, carbon and nutrient losses, disease outbreaks, increased production costs, low resilience to extreme climatic events, food insecurity and human health risks (e.g. Wall et al., 2015; Bender et al., 2016). These concerns have resulted in a strong demand among practitioners and decision makers for nature-based solutions 1 and agricultural practices that stimulate soil biodiversity or beneficial soil organisms and enhance soil health, thereby contributing to sustainable food production.
Soil health, seen as a powerful concept or metaphor by some (Lehmann et al., 2020; Janzen et al., 2021) but contested by others (Baveye, 2020; Powlson, 2021), has been defined as “the continued capacity of soil to function as a vital living ecosystem that sustains plants, animals, and humans” (Lehmann et al., 2020). Central to this concept is the multifunctionality of soils as living systems that host a tremendous diversity of soil biota that play a fundamental role in the form and functioning of natural ecosystems and agroecosystems alike. Through their activities and interactions, soil organisms mediate ecosystem processes that are critical for the continued biomass productivity of land and for the delivery of other ecosystem services such as the provision of clean water or greenhouse gas (GHG) mitigation (Bünemann et al., 2018). Soils are thus considered healthy when they support ecosystem functions such as nutrient cycling, carbon storage, water regulation, disease and pest suppression and contaminant regulation. ‘Functions’, are not limited to ‘services’, which tend to imply immediate human benefits, but also include processes that maintain stability of ecosystem properties and the biosphere (Hooper et al., 2000; Janzen et al., 2021). The concept of soil health can thus be applied to both natural ecosystems and agro-ecosystems, or other land uses, although the relative contribution of different types of ecosystems to different ecosystem functions and services varies. Soil health is therefore always context-dependent (Janzen et al., 2021). Unlike natural ecosystems, agricultural systems primarily, though not exclusively, focus on the production of nutritious and safe food or other agricultural goods (Bünemann et al., 2018). The emphasis on living soils highlights the importance of understanding the ecological processes and mechanisms that lead us to better ways of managing soils within their specific context (Janzen et al., 2021).
The importance of soil health and soil biodiversity is strongly emphasized in popular farming approaches such as agroecology, regenerative agriculture and conservation agriculture, in accordance with their aim to mimic biological processes in natural ecosystems to achieve sustainable food production (Mitchell et al., 2019; HLPE, 2019; Schreefel et al., 2020; Giller et al., 2021). Natural-ecosystem mimicry, or the use of natural processes or elements to improve ecosystem functions, is central to ecological intensification (Malézieux, 2012; Bommarco et al., 2013) and nature-based agriculture (Simelton et al., 2021). These concepts offer promise to reduce the dependency of modern agriculture on high amounts of external inputs, thereby diminishing environmental externalities and/or yield gaps, to achieve food and water security and climate goals (Bommarco et al., 2013; Dynarski et al., 2020; Miralles-Wilhelm, 2021). Currently there is a strong momentum among policy institutions, agri-food businesses and NGOs to promote practices that restore soil health, while contributing to key sustainability targets such as ‘carbon neutrality’, ‘net-zero emissions’, ‘nature-positive agriculture’ and ‘climate smartness’ (European Commission, 2015; HLPE, 2019; Miralles-Wilhelm, 2021; WBCSD 2 , 2018). At the same time, a growing awareness of the tremendous diversity of soil life has created a plethora of popular claims on the benefits of soil biodiversity for agricultural productivity and sustainability, as well as technologies, products, and services being offered to farmers. Questions arise, such as: “To what extent can knowledge on soil biology and associated processes in natural ecosystems inspire sustainable management of agricultural soils?”, “How can such knowledge be translated into practices or technologies that effectively improve soil functions in agriculture?”, and “How can such knowledge help to define indicators to monitor changes in soil health?”
In this paper we examine popular ideas and assumptions that have inspired nature-based soil management approaches, in relation to the scientific evidence for their potential to address key global challenges related to sustainable agriculture. First, we analyse the origins and ecological theories, including soil ecological theories, that underly the interest in natural ecosystems as a model for sustainable agriculture. Next, three examples of widely-used assumptions on soil health, for which the nature mimicry hypothesis is most explicitly used, are discussed: 1) a higher ratio of fungal-to-bacterial biomass favours soil carbon storage and nutrient conservation; (2) intensive agricultural practices lead to a decline in soil biodiversity with detrimental consequences for sustainable food production; (3) inoculation with arbuscular mycorrhizal fungi reduces agriculture's dependency on synthetic fertilizers. Throughout the paper, we quote a variety of sources to illustrate how nature-mimicry, and related ideas on soil ecology, have become part of a discourse on sustainable farming that extends from scientific publications to popular media and the promotion of commercial products to farmers. Lastly, we reflect on the normative use of nature as a metaphor for sustainability, and implications for future research in soil ecology in support of “nature-based solutions” in agriculture.
The origins of nature-based approaches in agriculture
The idea that agroecosystems should be managed more like natural ecosystems, referred to hereafter as ‘nature-mimicry’, dates back several decades for tropical agriculture (Ewel, 1986; Beckerman, 1983). More recently it gained momentum in temperate agriculture, finding support in concerns about the consequences of intensive farming, such as the dependence on large amounts of agrochemicals and fossil energy, with associated environmental problems, climate change impacts and human health risks (Malézieux, 2012; Caron et al., 2014; Struik and Kuyper, 2017).
Arguments for nature-mimicry were first made explicit by Ewel (1999) and Van Noordwijk and Ong (1999), who proposed that natural ecosystems of the region in which the agricultural system is embedded could provide appropriate models for sustainable agriculture (Sumberg, 2022, this special issue). This idea was based on the hypothesis that structure and function of natural ecosystems result from natural selection towards sustainability and adaptation to resource constraints (Ewel, 1999; Van Noordwijk and Ong, 1999). A second hypothesis stated that agroecosystem sustainability can benefit from resembling the diversity of natural ecosystems (Van Noordwijk and Ong, 1999). Van Noordwijk and Ong (1999) showed how these hypotheses, while rooted in old ideas of “Nature Knows Best” (Commoner, 1971), could be subjected to rigorous testing and thus be integrated into scientific agriculture.
The theoretical foundation for nature-mimicry can be traced back to E.P. Odum's classic paper ‘‘The strategy of ecosystem development’’ (Odum, 1969). Odum proposed a list of 24 structural and functional attributes, including soil attributes, that change during ecological succession. Among the list of ecosystem attributes that would characterise more mature systems, he mentioned a larger amount of organic matter, closed and less rapid cycling of nutrients, a shift from nutrients in mineral form to those held in organic matter, an increased species richness, greater stability in the face of external disturbance, a larger share of mutualistic interactions, more complex food webs and the presence of larger organisms with longer life cycles and slower growth (K-strategists) (Odum, 1969). According to his theory, the self-organization of ecosystems leads to higher stability (resistance to external perturbation) and more developed nutrient conservation, whereas earlier development stages are associated with higher biomass productivity and poorer stability. Odum (1969) further emphasized the analogy of early successional development stages with systems that are frequently disturbed by humans through intensive agriculture, burning, or flooding (e.g. for rice cultivation).
A clear link can be seen between the theories of Odum (1969) and key concepts for the design of agricultural systems since the 1970s (Caron et al., 2014). For example, Ewel (1986, 1999) described and tested how species-rich successional communities can inspire sustainable agroecosystems in the humid tropical lowlands because they offer traits of potential value to agriculture: efficient use of resources (nutrients, light, water), high productivity, high nutrient retention and high resistance and resilience to pests. Malézieux (2012) proposed that nature-mimicry can provide new ways for agroecosystem design that combine objectives of productivity and ecosystem services (referred to as ecological intensification; EI) and are applicable to temperate and tropical contexts. His 17 characteristics used to classify natural ecosystems vs. traditional and modern agriculture show significant overlap with the 24 attributes of Odum (1969), thus confirming the clear link between theories of ecosystem development and the concept of EI, more than 40 years later (Table 1).
Similarities and dissimilarities between theory on ecosystem development (Odum, 1969) and the classification of agricultural systems (natural ecosystems, traditional agriculture, modern agriculture) according to Malézieux (2012).

Odum (1969), Ewel (1986, 1999) and Van Noordwijk and Ong (1999) were well aware of the limitations of the analogy between natural and agricultural ecosystems. Odum (1969) referred to trade-offs between maximising productivity in early-stage ecosystems versus maximizing biomass in mature ecosystems and wrote that “recognition of the ecological basis for this conflict is a first step in establishing rational land use policies”. Ewel (1986) stated clearly that nature-mimicry cannot yet, and perhaps never, provide the yields that we expect from modern agriculture, because “the plant's photosynthetic energy may be allocated either to harvestable products or to the ecological functions that sustain complex ecosystems, but not to both simultaneously”. However, many subsequent proponents of nature-mimicry, especially in popular media (Box 1, 4) but also in some of the peer-reviewed literature (Box 2; Andrade et al., 2020; White 2020) are less cautious about these trade-offs.
Quote from Australian website Farmtable, illustrating how nature-mimicry, and related ideas on soil ecology, have become rooted in popular communication 9
“Overnight he stopped fertilising, and stopped resowing introduced pasture species.
Keeping the soil permanently covered with plant matter and using animals to naturally fertilise the paddocks allowed Colin to largely eliminate chemical use.
“By restoring the grasslands the whole soil ecosystem has changed,” Colin said.
Colin has also tripled the amount of carbon stored in his soil, which means he increased the soil's ability to sequester carbon dioxide in the air as well as its ability to absorb water. This increased his resilience to drought.
“It's worked for millions of years. All we need to do is mimic that a lot closer.” “If we mimic Mother Nature, it can’t not work,” Colin said.
…
“Soil biology is this perfect little symbiotic relationship, that we can embrace and work with, or we can come in and cultivate, we can come in and spray herbicides and disrupt that system and that cycle,” Derek said”
Quote illustrating how nature-mimicry and related ideas on soil ecology have rooted in scientific communication (Adreote and Pereira e Silva, 2017)
“Along their evolution, plants learned to interact to soil microbiota, extracting their utmost capacity to provide resources for plant development and successful colonization of terrestrial systems, where the great soil biodiversity is keen on properly exert this role”
—
“…if we better learn about the connection between plants and its associated microbiota in nature, we can lead agriculture to a better exploration of this omnipresent source of nutrients and protection, increasing yield and sustainability”
Nature-mimicry in soil biology
The recent literature on nature-based solutions and natural-ecosystem mimicry often emphasizes the importance of soil health, but rarely pays explicit attention to soil biology (e.g. Malézieux, 2012; Miralles-Wilhelm, 2021). This does not mean that soil biology has been neglected in the literature on nature-mimicry. Some of the foundational publications highlighted above describe changes in soil communities and soil biological processes during succession. Odum's list of 24 attributes that undergo change during succession includes many attributes that relate to below-ground components of the ecosystem, such as organic matter, nutrient cycling and retention, species richness and the share of mutualistic interactions (Odum, 1969; Table 1). Ewel (1986) indicated that restoration of soil quality after disturbance is biologically mediated, for example through the recovery of earthworm populations and deep rooting plants, recolonization of arbuscular mycorrhizal fungi and nitrogen-fixing symbionts, and increased recycling of organic matter during the fallow period. He also noted that, to achieve high yields many crops require high rates of fertilizer and this tends to switch off microbially-mediated nutrient acquisition such as rhizobia and mycorrhizal fungi (Ewel, 1986).
Important studies in the 1980s and early 1990s advanced theory on structure and function of soil food webs in natural and agricultural systems. This work included modelling of energy and nutrient flows based on trophic interactions under grassland (Hunt et al., 1987) versus forest soils (Ingham et al., 1989) and in arable cropping systems with different intensities of management (e.g. Hendrix et al., 1986; De Ruiter et al., 1994). Soil food webs in natural and agricultural systems across long-term field sites were shown to exhibit common responses to disturbances, due to soil tillage and changes in crop residue management (Moore, 1994). In line with the theories of ecosystem development, more disturbed (early successional) systems had poorer nutrient retention and were less stable (less resistant to external perturbation). These systems were also associated with a greater relative importance of the bacterial over the fungal-based energy channels (Hendrix et al., 1986; Ingham et al., 1989) and lower abundance or complete absence of higher trophic levels and soil fauna with larger body sizes (Hendrix et al., 1986; Wardle et al., 1995). Moore (1994) and Wardle et al. (1995) emphasized the evidence that different disturbance regimes inherent to agricultural practices (in particular mulching, tillage, and weed control) affect food web structure through cascading effects with important consequences for nutrient cycling and organic matter decomposition. In this context, Wardle et al. (1995) clearly linked food web theory to concepts of nature-mimicry by stating that it was increasingly evident that “those systems that bear the closest resemblance to natural ecosystems may require fewer inputs because greater reliance can be placed upon ecosystem self-regulation”.
In recent years, the study of soil communities has advanced into new directions, focusing increasingly on the importance of soil biodiversity for multiple ecosystem processes and functions, i.e., the concept of ‘soil multifunctionality’ or ‘soil health’ (Bender et al., 2016; Bünemann et al., 2018). Such studies generally encompass different types of interactions in soil communities beyond trophic interactions, including soil ecosystem engineering and rhizosphere interactions in support of sustainable production and ecological intensification (Pulleman et al., 2012; De Vries and Wallenstein, 2017; Bloor et al., 2021). This is also reflected in the idea that agriculture has interfered in rhizosphere biodiversity, including beneficial interactions between plants and soil microbes, and that we can learn from plant-soil interactions in nature to improve agricultural systems (Box 2; Andreote and Pereira e Silva, 2017).
Examples of nature-based soil health approaches and underlying assumptions
A clear relation exists between the ecological theories underlying nature-mimicry as discussed above, and commonly promoted practices to enhance soil health (Table 2). Many of the desired benefits of such practices and products thus seem to follow a change towards a more mature ecosystem state, in line with Odum (1969) and Malézieux (2012) (Table 1). Whereas the focus for sustainable soil management may have shifted from local challenges and specific soil functions in the past century, to global challenges and multifunctionality of soil communities at present, ideas on ‘nature-based’ soil management practices have not changed fundamentally over the past decades (Giller et al., 2021). What has changed is that soil biodiversity attracts increased interest and concern among diverse societal actors because of assumed links to soil health and consequences for sustainable food production. It has also created new momentum for the promotion of regenerative practices, products (e.g. inoculants) and services (e.g. microbial tests as potential metrics of soil health) that build on widely established assumptions on the role of soil biology in sustainable agriculture. Three examples of such assumptions, for which the nature-mimicry hypothesis is most explicit, are discussed in more detail below.
Examples of assumptions on the desired change in the structure or function of soil communities that can support sustainable agroecosystems; how they relate to commonly promoted soil-health practices or indicators; the expected benefits in terms of soil functions and associated societal benefits. The examples that are discussed in more detail in the current paper are marked with a #.
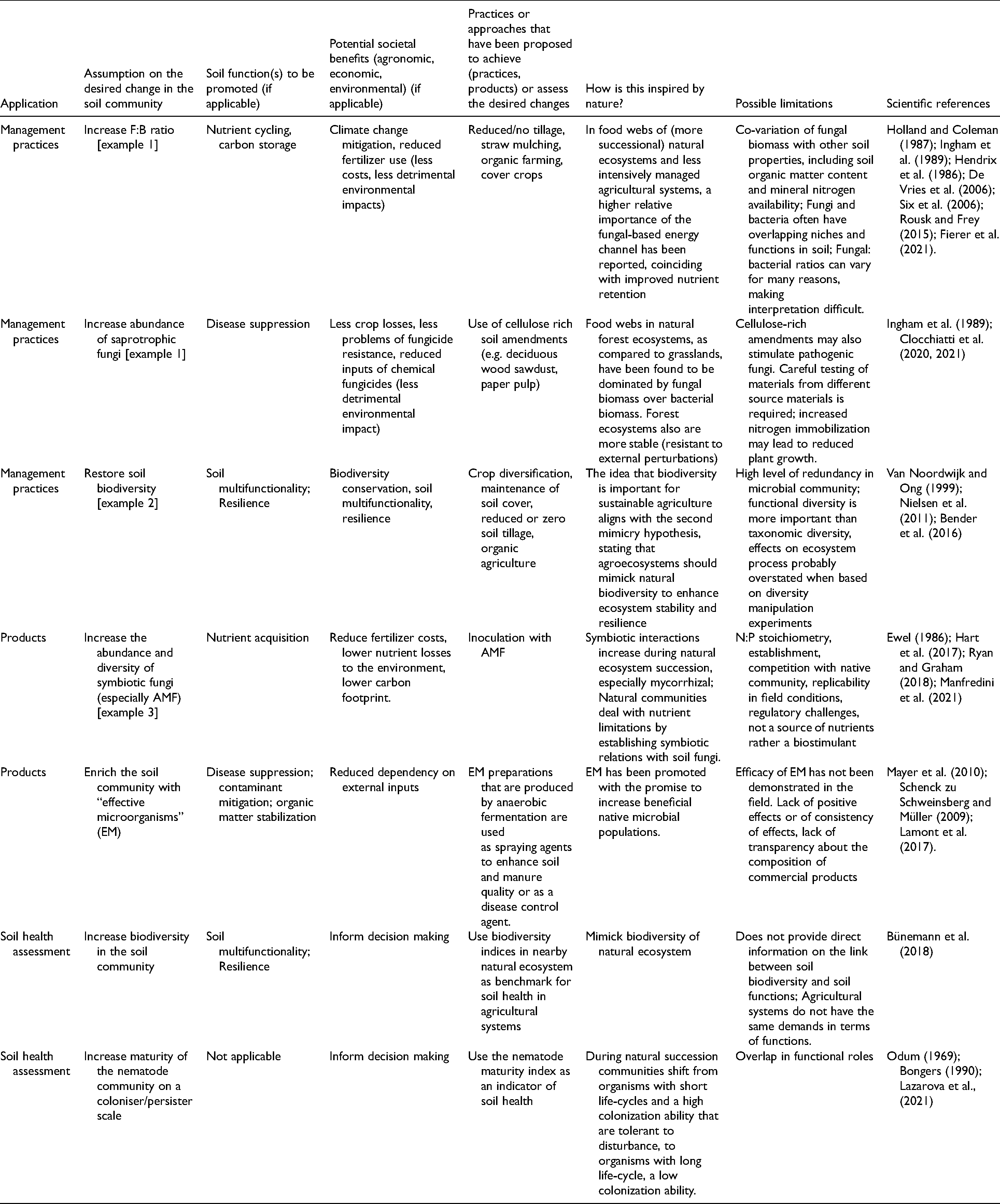
Example 1: A higher fungal-to-bacterial ratio favours soil carbon storage and nutrient conservation
The hypothesis that a greater biomass of fungi relative to bacteria (a higher F:B ratio) results in more stable soil organic matter and reduced risk of nitrogen emissions dates at least back to Waksman et al. (1928) and was attributed to a more complete humification of organic matter and greater contribution of ammonium than nitrate in soils that favour fungal communities. Subsequent soil food web studies showed that native ecosystems and less disturbed agroecosystems had higher F:B ratios, which also correlated with more conservative carbon and nitrogen cycling (Ingham et al., 1989, Hendrix et al., 1986; Holland and Coleman, 1987; see previous section). Similar correlations were found for grasslands with lower nitrogen input (De Vries et al., 2006) and for chronosequences of abandoned arable land (Van der Wal et al., 2006; Morriën et al., 2017). By contrast, De Vries et al. (2021) reported that the F:B ratio did not change during ecosystem succession and concluded that increased N retention in ecosystems with a higher F:B ratio was due to low soil N availability.
Although the fore-mentioned studies have not provided direct evidence for a causal relationship between fungal dominance and conservative C and nutrient cycling, they have nourished the theory that the bacterial decomposer channel (associated with high nitrogen sources, high soil disturbance and an r-selected life strategy) supports fast turnover of easily available organic substrates, while the fungal-dominated channel (associated with lignocellulosic organic compounds, less soil disturbance and a K-selected life strategy) supports slower decomposition of more complex organic matter and retention of nutrients. In line with this theory, the idea that a high F:B ratio is a desirable property of agroecosystems and composts and indicative of healthy soils has gained the status of received wisdom. This idea has inspired commercial soil health tests that are offered to farmers to inform agricultural management 3 , 4 (Fierer et al., 2021) and are at the basis of overoptimistic claims about the benefits of manipulations of soil communities and F:B ratios on crop yields and carbon sequestration that have not been scrutinized by peer review (Box 3).
Quote from Dr David Johnson
10
. This
“Applying agricultural management practices to enhance SMC population and F:B structure, in a 4.5 year agricultural field study, promoted annual average capture and storage of 10.27 metric tons soil C ha-1 year −1, 20–50 times the currently observed soil carbon increase in agricultural no-till soils. These soil C% and F:B increases also promote increasing soil macro-, meso- and micro-nutrient availability offering a robust, practical and cost-effective carbon sequestration mechanism within a more productive and long-term sustainable agriculture management approach”
Quote from a video on the website of the food web school of Elaine Ingham 11 claiming that a ‘balanced’ soil food web can replace fertilizer use in agricultural soils while hugely increasing yields.
“The good news is that we can restore the soil food web of the most soils within just a few months. This results in a number of benefits, both for the farmers and for the environment. With a balanced soil food web in place, farmers need not use fertilizers at all. They don't need to use pesticides either, as nature's operating systems protects plants from attacks. Herbicides, use to kill weeds, are not required either as weeds only thrive in conditions where the food web is out of balance”
—
“It also means that their yields increase dramatically, in some cases, farmers working with Dr Elaine Ingham, have seen yield increase by over 200%. This is because the soil food web provides plant access to a constant flow of nutrients from soil organic matter and from the soil particles themselves.”
Several scientific studies have discussed the evidence for the presumed relation between F:B ratio and soil carbon or nutrient cycling (Six et al., 2006; De Vries et al., 2013; Rousk and Frey, 2015) or on its use as a general soil health metric for agricultural soils (Fierer et al., 2021). Mechanisms that could explain why a greater F:B ratio would favour soil carbon storage include (i) the widespread assumption that fungi have a higher metabolic growth efficiency than bacteria (Six et al., 2006; Kallenbach et al., 2016), (ii) greater stabilization of microbial-derived SOM in the presence of substrate additions that favour fungal abundances (Kallenbach et al., 2016) and (iii) a greater contribution of fungi than bacteria to physical protection of soil organic matter through soil aggregate formation and interactions with clay minerals (Witzgall et al., 2021). Yet, Strickland and Rousk (2010), based on review of literature, concluded that shifts in the F:B ratio along environmental or management gradients, as well as relationships with ecosystem functioning, frequently contradicted general theory. This observation was confirmed by Rousk and Frey (2015) who found no consistent relationship between soil carbon quality, F:B ratios and carbon use efficiency across study sites. A major reason for the lack of consistent scientific support for generalized theories on the role of fungi versus bacteria in carbon and nutrient retention is the much larger diversity of lifestyles within groups than between groups of fungi and bacteria, while the lack of attention to interactions between both groups in ecological networks further contributes to the maintenance of a simplistic view of separate energy channels (Ballhausen and de Boer, 2016; De Menezes et al., 2017; Fierer et al. (2021).
It thus remains unclear whether a higher F:B ratio is a driver for, or a response to, the larger amounts of soil carbon and nitrogen retained in less disturbed ecosystems or in systems with lower substrate quality (Six et al., 2006; De Vries et al., 2013; De Vries et al., 2021). Moreover, soil F:B ratios can vary for many reasons. This further complicates the interpretation of differences in F:B ratios between natural and disturbed ecosystems and among soils (Fierer et al., 2021), and the use of F:B ratios found in natural ecosystems as a benchmark to guide management decisions in agricultural soils. For example, the greater F:B ratio in the top 5 cm of forest soil as found by Ingham et al. (1989) may (partly) be driven by the abundance of ectomycorrhizal (EcM) fungi. However, EcM fungi are absent in agricultural soils. Differences in soil pH or clay mineralogy may also explain variation in F:B ratios (Six et al., 2006; Kallenbach et al., 2016; Fierer et al., 2021).
We conclude that the idea that a higher F:B ratio favours a more conservative carbon and nutrient turnover does not reflect current ecological understanding of complex, multitrophic soil food webs. The same limitation then applies to the use of F:B ratio as an easily interpretable and robust indicator of soil health (Fierer et al., 2021). Our general understanding on how the composition of soil communities in agricultural soils affects carbon and nitrogen cycling is currently insufficient to guide management and policy decisions.
Besides nutrient and carbon cycling, saprotrophic fungi provide other soil functions such as disease suppression and soil structure formation (Lehmann et al., 2017; Ghorbanpour et al., 2018), so there may be other reasons to stimulate fungal biomass in agroecosystems, for example, by making use of their ability to degrade certain substrates (e.g. with higher lignocellulose concentrations). Clocchiatti et al. (2020; 2021) showed that the addition of cellulose-rich soil amendments like wood sawdust or paper pulp can stimulate saprotrophic fungi and associated disease suppression (Table 2). It should be noted that adding soil amendments with high C:N ratios (including so-called fungal compost) can induce strong N immobilization during the first stages of decomposition and thus present a trade-off with crop productivity or fertilizer use (Clocchiatti et al., 2020).
Example 2: intensive agricultural practices lead to a decline in soil biodiversity with detrimental consequences for sustainable food production
Agricultural intensification and loss of soil biodiversity
Soils contain a bewildering and fascinating diversity of living organisms that attracts growing visibility and appreciation among researchers and the public at large (Orgiazzi et al., 2016). At the same time, intensive agriculture, through habitat destruction or disturbance, has been associated with an overall loss of soil biodiversity with potentially dramatic consequences for ecosystem service provision, global food security and human health (Wall et al., 2015; Bender et al., 2016). Such concerns about loss of soil biodiversity and function are also reflected in communications to the general public. Apocalyptic headlines appeared in Dutch media a few years ago that referred to agricultural soils being ‘zombie soils’ and ‘as dead as a dodo’, according to their English translation) 5 , 6 Yet, we should be aware that such statements are blatant exaggerations and that effects of agriculture on soil biodiversity are not easily generalized.
Review of the scientific literature shows that the effects of land use and agricultural management on the biomass and taxonomic richness of soil biota depends strongly on the soil fauna or microbial group considered (Rutgers et al., 2009; De Graaff et al., 2019) and can vary according to climatic region (Trivedi et al., 2016). Based on meta-analysis at continental scale, Trivedi et al. (2016) found significantly higher soil bacterial diversity in agricultural than natural systems of arid and temperate climatic regions, which may be attributed to higher nutrient availability in agricultural soils. De Graaff et al. (2019) used meta-analysis to quantify effects of agricultural intensification on soil biodiversity across studies. They concluded that intensive agricultural practices (e.g. synthetic N fertilization, tillage) mostly showed negative effects on AMF and faunal diversity, but synthetic N fertilization positively affected on fungal- and bacterial diversity.
Some general limitations to the interpretation of studies on the relations between agricultural intensification and soil biodiversity should be highlighted. First, studies often focus on indicators like taxonomic richness or Shannon index which can mask differences in community composition. Different species or taxonomic groups may respond to disturbances in a more predictable manner than overall richness (De Graaff et al., 2019). For example, Trivedi et al. (2016) found that different phyla of soil bacteria responded differently to land use resulting in consistent patterns at the continental scale, although these responses could not be related to growth rates and or substrate preferences. Another important limitation is that many scientists focus on specific groups of soil organisms according to their own discipline, while in reality soil communities consist of multiple trophic levels that interact in complex ways (El Mujtar et al., 2019). There is compelling evidence that food webs in forest soils are more diverse than in agricultural land in terms of the number of trophic links and food-web complexity (Rutgers et al., 2009) and that soil disturbances related to agriculture have stronger negative effects on higher trophic levels in the food web and on soil fauna with larger body sizes (Hendrix et al., 1986; Wardle et al., 1995; Postma-Blaauw et al., 2010; Tsiafouli et al., 2015; De Graaff et al., 2019). Bloor et al. (2021) reported that agricultural intensification reduced trophic group diversity and connectivity in soil networks, but not necessarily taxonomic richness. All these studies confirm that different forms of agricultural management result in different soil communities, but the responses are not as predictable as people might assume in terms of the “biodiversity of the system” (Wardle et al., 1995).
Loss of soil biodiversity and effects on soil functions
Regarding the general idea that loss of soil biodiversity jeopardizes the general sustainability and continued productivity of agricultural land, we can clearly link this to the second nature-mimicry hypothesis of Van Noordwijk and Ong (1999) and related concepts of productivity, efficiency, stability, and resilience of natural ecosystems that have inspired the concept of ecological intensification (Malézieux, 2012; Table 1). The specific importance of diversity of soil communities for ecological intensification was discussed by Bender et al. (2016) who concluded that to enable the proper functioning of ecosystems, soil biodiversity must be enhanced and maintained. Yet, our reading of the scientific literature suggests that assumptions about the consequences of soil biodiversity loss for ecosystem functioning and sustainable food production are particularly hard to ground.
A number of recent studies and meta-analyses suggest that variation in soil biodiversity does explain differences in ecosystem processes, including organic matter decomposition, carbon mineralization, nutrient losses and plant production (Wagg et al., 2014; Bender and Van der Heijden, 2015; De Graaff et al., 2015; Tardy et al., 2015; De Graaff et al., 2019). Yet we should be cautious of fundamental methodological issues associated with these studies when interpreting the observed effects of changes in taxonomic or functional group richness on ecosystem processes. Studies are of two main types. First, those that are based on correlations that do not allow for the separation of cause and effect; Second, those that apply experimental manipulation of diversity levels using dilutions or inoculations of sterilized soil that likely overestimate the effects on ecosystem processes relative to measured biodiversity losses from environmental perturbations (De Graaff et al., 2019).
Field studies in agricultural soil contaminated with heavy-metal can provide an interesting opportunity to study the importance of soil biodiversity for soil processes, ecosystem function and resilience. Such pollutants persist in soil over time and even small concentrations of heavy metals can cause drastic reductions in soil biodiversity, including microbial diversity (Sandaa et al., 2001; Giller et al., 2009). Yet, Giller et al. (1998) found surprisingly little disruption of ‘generalist’ functions such as decomposition. This observation fits with the understanding that soil microbial communities comprise such high diversity and functional redundancy that a loss of diversity does not result in loss of ‘generalist’ soil functions (Kuyper and Giller, 2011). Functional redundancy can thus provide a high level of resilience to disturbance (Giller et al., 1997; Nielsen et al., 2011). By contrast, it has been shown that changes in soil food web structure and loss of so-called “keystone species” that confer specific functions and belong to physiologically and phylogenetically ‘narrow’ organism groups can result in a drastic loss of function. Examples of such keystone species are Rhizobium strains that fix nitrogen with a specific legume (McGrath et al., 1988; Wardle et al., 1995; Schimel and Schaeffer, 2012) or ecosystem engineers such as earthworms and termites (Jongmans et al., 2003; Paul et al., 2015). In the context of resilience, a pertinent question is to what extent changes in soil biodiversity are reversible, or to what extent do observed changes in soil represent a ‘loss’ of biodiversity or simply a change in the relative abundance of different taxonomic groups. If a decrease in biodiversity is the result of a local disturbance (for example, soil tillage or a reduction in organic inputs), a change in management could restore below-ground diversity relatively quickly.
In summary, current insights imply that simplification of soil food webs and the loss of particular soil biota can affect soil functions and that community composition often has stronger effects than taxonomic richness per se. This highlights the difficulties in applying an umbrella concept like soil biodiversity (Hooper et al., 2000), because relations are likely to vary from function to function and depend on the way that biodiversity is defined (e.g. taxonomic richness, functional groups, community structure) and what levels of organization are considered (single trophic levels, food webs) (Lazarova et al., 2021). Translating knowledge on soil biodiversity into universally applicable soil management recommendations to enhance food production therefore remains challenging (El Mujtar et al., 2019).
Example 3: inoculation with arbuscular mycorrhizal fungi reduces agriculture’s dependency on synthetic fertilizers
Odum (1969) and Ewel (1986) noted that during ecosystem development symbiotic interactions increase and they specifically referred to the mycorrhizal symbiosis between trees and certain root-inhabiting fungi as a strategy to overcome increasing deficiencies in plant-available P with ecological succession. Agricultural practices, including soil disturbance through tillage, high doses of mineral fertilizer (especially phosphorus), long-term bare fallow and the use of fungicides, reduce species richness and abundance of mycorrhizal fungi (Verbruggen and Kiers, 2010). Based on the apparent incompatibility of mycorrhizal fungi with intensive agricultural practices and its overtone of mutual help, mycorrhizal symbiosis has become symbolic for relations between plants and soils, and between humans and nature (Sheldrake, 2012). Moreover, mycorrhizal symbiosis has strongly inspired nature-based solutions in agriculture.
Almost all agricultural crops can form symbiotic relationships with arbuscular mycorrhizal fungi (AMF). In this context, AMF have often been referred to as biofertilizers (Berruti et al., 2016; Madawala, 2021), based on an incorrect analogy with nitrogen-fixing bacteria such as rhizobia and actinobacteria that form nodules with certain shrubs and trees and fix atmospheric nitrogen into plant-available nitrogen. However, AMF do not add nutrients to the soil and there is no evidence that they have a positive impact on the soil phosphorus pool over ecologically-relevant time scales. Instead, they extend the extraction zone of nutrients that are already present in the soil and make poorly mobile nutrients such as phosphorus, zinc or copper, available to the plant. Additional benefits of the AMF symbiosis include disease resistance, drought tolerance and soil structure formation, as described by Kuyper et al. (2021). These are considered beyond the scope of this example as they do not directly relate to nutrient acquisition.
Within the nature-based solutions discourse, AMF play a rather ambiguous role. It is sometimes claimed in the popular media that fields of modern “industrial” agriculture are mycorrhizal deserts 7 and that inoculation is required to restore mycorrhiza. Commercial biofertilizers are a rapidly growing global business and the use AMF inoculum is widely promoted based on bold claims of yield enhancement and input reductions (Berruti et al., 2016) 8 . In the scientific literature the importance of AMF for (sustainable) crop production is a topic of active debate. In their review, Ryan and Graham (2018) argued that there are currently no demonstrated benefits that warrant the use by farmers of specific AMF management practices, and showed that the mycorrhizal fungal community may be more resilient to many agricultural practices than often assumed. Other recent studies suggested the opposite and reported clear benefits of indigenous AMF stimulation on nutrient acquisition efficiency in maize (Wang et al., 2020) and of AMF inoculation on yields of several cereal crops (Pellegrino et al., 2015; Zhang et al., 2019). Such discrepancy may be explained by the type of management interventions included in the studies, as well as crop and context-specific responses of AMF to such treatments. For example, Ryan and Graham (2018) and Wang et al. (2020) focused on specific crop management practices that can stimulate indigenous AMF, while Zhang et al. (2019) included a large set of field studies using inoculations.
The general picture that arises is that scientific evidence on the benefits of inoculation with (commercial) AMF in cropping systems where indigenous AMF communities are well-established is inconsistent (Hart et al., 2017). Effects of AMF inoculation on crop yields and nutrient acquisition under field conditions are extremely variable and crop and context specific (Ryan and Graham, 2018; Kokkoris et al., 2019; Zhang et al., 2019). Such variation has been attributed to a range of factors or limitations that can affect inoculant survival and establishment. Those may include low quality of commercial inoculants in the absence of universally adopted regulation to guarantee product quality (Manfredini et al., 2021; Salomon et al., 2022), competition with the indigenous AMF community, or abiotic soil conditions (Pellegrino et al., 2015; Hart et al., 2017; Kokkoris et al., 2019; Thomsen et al., 2021). Unfortunately, in most field studies, inoculant establishment is not reported as it is difficult to track the introduced inoculum in indigenous communities (Hart et al., 2017; Manfredini et al., 2021). Many studies have shown that AMF inoculation has no consistent agronomic benefit where AMF communities are well established (Hart et al., 2017; Bender et al., 2019). Inoculation with AMF can also result in trade-offs in N-limited soil due to competition between AMF and plants for nitrogen (Johnson et al., 2010). By contrast, recent meta-analyses have demonstrated overall positive, though variable, yield effects of AMF inoculation in cereal crops (Pellegrino et al., 2015; Zhang et al., 2019). It remains unclear whether these yield benefits of AMF resulted from AMF-mediated improvements in plant nutrient acquisition or from non-nutritional AMF benefits (Zhang et al., 2019) and to what extent fertilizer reductions may be possible without compromising yields.
We conclude that many questions remain regarding the soil and management conditions under which AMF inoculation is likely to be successful and economically profitable. Moreover, it is not clear to what extent, under what conditions and by which mechanisms AMF inoculations may help to reduce the use of synthetic fertilizers. Such questions need to be answered before large-scale recommendations on AMF management in sustainable cropping systems can be provided. Interesting, from a nature-mimicry perspective, is the hidden assumption that plants in agricultural systems make insufficient use of natural biological processes such as AMF-mediated nutrient acquisition, unless these AMF are added through a (commercial) inoculum. Pleas for AMF inoculation without addressing the causes of the inoculum insufficiency in fact reflect a commodification of nature rather than a nature-based solution (Oviatt, 2020).
Discussion
Dichotomies and pitfalls associated with the normative use of nature as metaphor
We have shown how ecological arguments, and especially those derived from theories on ecosystem succession, have been used to argue for the superiority of alternative agricultural approaches that mimic the structure and function of natural ecosystems, including natural soil communities. These alternative approaches, which have been promoted under the terms of “ecological intensification” and “agroecology” define themselves, or are defined by others, as a radical alternative to current agriculture (HLPE, 2019; Sumberg, 2022, this special issue). At the same time, nature-based approaches are being adopted by NGOs and multi-national companies as part of a more mainstream discourse on the need for climate-friendly, regenerative agricultural practices (Giller et al., 2021; Simelton et al., 2021) and sustainable intensification (Maes and Jacobs, 2015). This shows that the adjective nature-based can fulfil different roles in discourses on the future of agriculture. Analogous to the debate on ecological versus sustainable intensification (Kuyper and Struik, 2014) we can discern two broadly opposing discourses: the “Nature Knows Best” strategy which is ultimately rooted in the claim that ecosystems have been tested over time and selected for their sustainability, and the “Science-and-Technology Knows Best” strategy that is inspired by nature, but not necessarily mimics nature. One side may adhere to the Einsteinian dictum that “we cannot solve our problems with the same thinking we used when we created them” and that sustainable transformation should be achieved using holistic approaches that embrace complexity and nature (Box 5) (Altieri, 1999; Tittonell, 2014; HLPE, 2019). The other end of the spectrum emphasizes the role of empirical science in creating conditions where ecological and evolutionary processes and constraints can be smartly engineered to “improve nature” (Box 6) (Brussaard et al., 2010; Denison and McGuire, 2015).
Quote from Altieri (1999) in peer-reviewed literature
“Intercropping, agroforestry, shifting cultivation and other traditional farming methods mimic natural ecological processes, and their sustainability lies in the ecological models they follow. This use of natural analogies suggests principles for the design of agricultural systems that make effective use of sunlight, soil nutrients, rainfall, and biological resources. Many scientists have now recognized how traditional farming systems can be models of efficiency as these systems incorporate careful management of soil, water, nutrients, and biological resources”
Quote from blog by Andrew McGuire, Washington State University (2014) 12
“This strategy [mimicking nature] arises from a long history of thinking that there exists a “balance of nature.” This idea has greatly influenced how we look at nature and agriculture. In the latter case, it drives much of what is done in organic farming and agroecology, but also finds its way into no-till farming. Nonetheless, it is false, and because it is false we can abandon the restrictive “nature knows best” argument in designing agricultural systems. Instead, we can improve on nature”
Next to that ambiguity, the adjective nature-based is both part of an analytical-scientific and of a metaphorical-rhetorical discourse. The use of carbon-rich substrates to specifically stimulate a guild of cellulolytic fungi that then outcompete other fungal guilds and that result, under certain specified conditions, in pathogen suppression (Clocchiatti et al., 2020, 2021) is an example of a nature-based practice rooted in empirical science. By contrast, claims that restoring food webs in agricultural soils makes the use of synthetic fertilizers superfluous is a nature-based solution rooted in rhetoric (Box 4). However, the boundary between scientific and rhetorical arguments for nature-based solutions is not always evident as our reflections on the use of the F:B ratio as an indicator of sustainable soil management show.
The use of the concepts “nature-based” and “nature-mimicry” as a trump card in debates that belong to different agendas on the sustainability of agriculture, shows that nature is sufficiently ambiguous to serve opposing perspectives and approaches. In this special issue, Lenné and Wood (2022) argue that monodominant vegetation is an appropriate model for ecologically sound cereal production systems. This contradicts the widely-held paradigm that mimicking high biodiversity levels of natural ecosystems, both aboveground or belowground, is essential to the sustainability and resilience of food production, whereas monocultures are ecologically dysfunctional, vulnerable and require large amounts of external inputs to be maintained. As shown by Lenné and Wood (2022), natural monodominant vegetation is common in nature and our ancestors could have argued that they mimicked natural disturbances such as fire or flood in their fields to favour annual crops. An example from soil ecology is the cultivation of certain mushrooms by termite colonies, a successful and evolutionary old relation between a crop-cultivating insect and a crop that is based on monocultures of a fungus without apparent problems with infectious diseases (Otani et al., 2019).
We argue that nature-based solutions are prone to what we refer to as the ecologistic fallacy (in analogy of the naturalistic fallacy, the derivation of values from facts). The ecologistic fallacy is based on the normative idea that there is an inherent optimality principle in natural selection - so when succession goes from open to closed nutrient cycles, from bacteria-dominated to fungi-dominated systems, or from species-poor towards species-rich communities, there must be an inherent benefit of such closed, fungi-dominated, species-rich soil systems. The assumption that our agriculture must then mimic those processes would only be valid if natural ecosystems (including soil communities) had been selected for optimising sustainable yield, for which there is no evidence (Denison and McGuire, 2015). It is incumbent upon us to maintain an open mind in observing and understanding the roles of soil communities in soil health and ecosystems functioning and to translate that knowledge into solutions that support sustainable agriculture (Fierer et al., 2021).
Implications for soil ecology research and sustainable soil management in agriculture
We have also shown that scientific evidence on the assumed benefits of popular nature-based soil management approaches (increase F:B ratios, restore soil biodiversity and AMF inoculation) for sustainable agricultural production is weak and cannot be generalized. Current insights imply that intensive agricultural practices lead to changes in community composition, including simplification of soil food webs and loss of keystone species, and that such changes in community composition are more important for soil functions than taxonomic richness per se. Applying a unifying concept like soil biodiversity to predict impacts on soil functions and sustainable agricultural production therefore remains a huge challenge. Building on Kuyper and Giller (2011) we conclude that in the absence of a clear understanding of the linkages between soil biodiversity and ecosystem functions, soil biodiversity has become a powerful metaphor to signify a ‘living soil’. In the past decade, with the support of many soil scientists, the metaphorical use of soil biodiversity, similar to soil health (Janzen et al., 2021), has been instrumental in focusing attention on soils in policy and public circles. Yet, we question if such a metaphorical use of the term soil biodiversity is helpful if we are to advance our understanding of soil ecology to inform context-relevant approaches for sustainable soil management, beyond “holistic approaches” that enhance soil life and multifunctionality (Bender et al., 2016; El Mujtar et al., 2019) and magical ‘‘win–win’’ solutions (Box 3,4; Amundson, 2022).
So, what is the way forward for "nature-based" soil management in agroecosystems? First, based on our review we propose that we need studies that focus on community composition and abundances rather than taxonomic richness and that consider the interactions between organism groups at different trophic levels. The rapid advancements in low-cost and high throughput sequencing methods will provide new insights on how soil communities respond to different disturbances (De Graaff et al., 2019). Second, we should be aware of, and where possible overcome, methodological issues that complicate the establishment of causal relationships between soil communities, soil functions and the subsequent extrapolation to real ecosystems. Most studies are correlative and it is very hard, if not impossible, to manipulate soil biodiversity in a realistic way without changing other properties of the system (Hooper et al., 2000; De Graaff et al., 2015). Third, we identified a strong need to place research on the role of soil biodiversity and soil communities in agroecosystems in the broader context of crop agronomy, breeding and agroecosystem sustainability. This includes careful consideration of limitations and possible trade-offs between soil biodiversity and agronomic performance indicators including crop yields (Vazquez et al., 2020). Finally, we propose that targeted stimulation of beneficial soil biota that are naturally present in soils deserves more attention as a basis for the development of cost-effective sustainable management strategies.
Footnotes
Acknowledgements
We are grateful to two anonymous reviewers for their insightful comments on a previous version of the manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship and/or publication of this article.