
Abstract
The African spiny mouse (Acomys spp.) is an emerging animal model with remarkable biological characteristics that make it a subject of interest for a broad range of research fields. Typically a desert species adapted to a low-calorie diet, spiny mice develop diabetes-related symptoms when switched to high-energy diets. Spiny mice undergo relatively long gestation periods and have small litters of highly developed pups, making them an adequate model for late organogenesis and perinatal biology. Recently, they have been shown to have remarkable healing and regeneration capabilities, which make them unique among mammals. In this work, we describe our experience in housing a colony of African spiny mice and cover all basic aspects of feeding, maintenance and breeding for research purposes.
Introduction
The genus Acomys, commonly known as the spiny mouse, is a small rodent distributed throughout Africa, the Middle East and south-west Asia. 1 It was first described in 1803 by E. Geoffrey 2 and derives its common name from the presence of thick hair spines on its back. The genus presents a number of interesting biological characteristics that have been the subject of research since the 1950s, when the first laboratory colonies were established. Early interest focused on Acomys as a model of type 2 diabetes, based on the observation that animals fed high-fat diets developed obesity and hyperglycaemia. 3 A second area of interest is based on the fact that this genus has a relatively long gestation period (approximately 39 days) after which pups are born in an advanced state of development. Therefore, the spiny mouse is used as a model for organogenesis and late pregnancy biology.4–6 More recently, Acomys has been demonstrated to undergo menstruation (a rare characteristic in mammals) and may become an important rodent model for the study of the menstrual cycle. 7 The genus has also emerged as a model of mammalian regeneration due to its remarkable healing and regenerative properties.8–10 In this paper, we present an overview of Acomys husbandry based on several years of experience in housing a colony. Animal husbandry and procedures were carried out as regulated by both Portuguese law (DL 113/2013) and European law (directive 2010/63/EU). Projects at our institute involving Acomys spp. are approved by the Animal Welfare Body of the University of Algarve and by the competent authority (Direcção Geral de Alimentação e Veterinária). The Center for Biomedical Research (CBMR-UALG) bioterium is a licensed animal facility for small rodents.
Nomenclature, taxonomy and geographical distribution
Acomys derives its scientific name from the Greek ‘acme’ (pointy, referring to the shape of the snout) and ‘mus’ (mouse). 11 Its common name, ‘spiny mouse’, describes the thick spine-like hairs along its dorsum, which are presumably an anti-predator adaptation. Currently, the International Union for the Conservation of Nature (IUCN) recognizes 18 distinct species of Acomys. However, the taxonomy within the genus Acomys remains somewhat controversial, with the number of species that compose it varying considerably as the genus has been studied over the years. 12 As its common name suggests, Acomys had traditionally been classified as a murid based on various morphological traits.13–16 However, analysis of biochemical and immunological data called this classification into question,17–19 and further DNA-based analysis, including DNA–DNA hybridization analysis 20 and direct sequencing of the anti-chemotrypsin or cytochrome b genes12,21,22, found strong evidence of a close phylogenetic relationship with gerbils, leading these authors to propose classifying Acomys within the subfamily Gerbilinae. However, more recent studies based on the sequences of multiple nuclear genes support the inclusion of Acomys in the subfamily Deomyinae.23,24
The spiny mouse is found throughout Africa, the Middle East and south-west Asia. Within the genus, some species receive their common names after their main geographical location and are therefore referred to as Egyptian spiny mouse (Acomys cahirinus), Cretan spiny mouse (A. minous), Cyprus spiny mouse (A. nesiotes), Arabian spiny mouse (A. dimidiatus) or Turkish Spiny Mouse (A. cilicicus). Other species names commonly refer to coat colouring (golden spiny mouse (A. russatus) and grey spiny mouse (A. cineraceus)), while several others go exclusively by their scientific names, such as A. kempi, A. percivali, A. wilsoni, etc. A thorough description of its habitats and geographical range can be found in The IUCN Red List of Threatened Species 2017-3.
25
Briefly, within Africa, the genus has a wide geographical range (Figure 1), with various species being found in Maghreb and Egypt, Ethiopia, Sudan, Kenya, Uganda, Tanzania and as far south as South Africa. In the Middle East, a number of species are found in Turkey, Jordan, Israel and the Arabian Peninsula. The A. cahirinus/A. dimidiatus group has branched out to the Mediterranean islands of Cyprus (A. nesiotes) and Crete (A. minous), but the genus has not been found in the wild in southern Europe. In south-west Asia, Acomys has been reported in Pakistan and Iran.1,26–29 It typically inhabits dry, semi-arid, rocky desert areas or short grasslands. In Kenya, it has been described as inhabiting rocky canyons, cliff bases and rock formations known as kopjes. The species can be commensal with man, inhabiting the crevices of human dwellings
11
or associated with fruit orchards, palm groves or gardens. Acomys do not dig burrows or build nests, tending to take refuge in whatever shelter the terrain offers.
Geographical distribution of Acomys species (this image was created using information available in the International Union for the Conservation of Nature Red List of Threatened Species, Version 2017-3;
25
www.iucnredlist.org).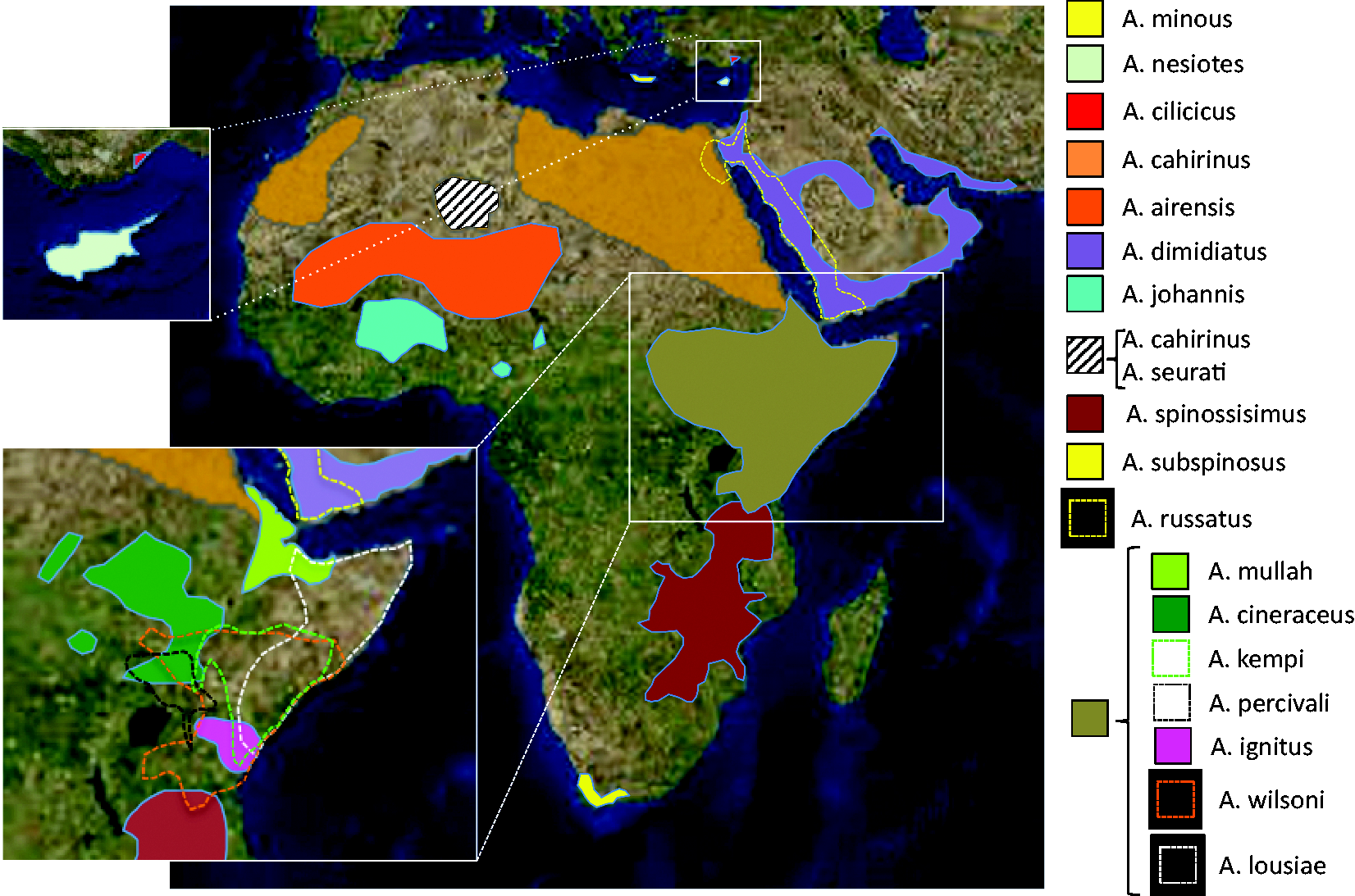
Basic biology
Acomys is somewhat larger that the average laboratory Mus musculus. 1 In our colony, at 1 year of age, their weight ranges between 39.05 and 63.41 g (N = 52). Average weight is 49.3 ± 6.1 g. Males weigh 49.9 ± 6.7 g (N = 24) and females 48.8 ± 5.4 g (N = 28), with no statistical differences between sexes (p = 0.87). Body length (snout to anus) ranges from 114 to 96 mm. Average body length is 106.8 ± 4 mm (N = 52). Males measure 107 ± 4.2 mm (N = 24), while females measure 106.6 ± 3.7 mm (N = 28), with no statistical differences between sexes (p = 0.67). Tail length ranges between 102 and 88 mm. Average tail length is 94.4 ± 3.6 mm (N = 46). Male tails measure 93.8 ± 3.3 mm (N = 22), while female tails measure 94.9 ± 3.8 mm (N = 24), with no statistical differences between the sexes (p = 0.3). In general, no sexual dimorphism is evident.
Animals have pointy snouts, relatively large protruding black eyes, large ears and a scaly tail with sparse short fine hairs. The dorsal coat colour is light golden brown peppered with reddish, black and white hairs (A. cahirinus), with some species presenting colourations ranging from grey (A. percivali and A. cineraceus) to an almost rusty orange (A. russatus). The ventral coat is significantly lighter, usually a creamy white. Spiny mice have a complex social organization including individual and kin recognition,30,31 communal breeding 32 and paternal care. 33
Pups are born in a relatively advanced stage of development. This precocity contrasts with the more common altricial strategies of other rodents (mice and rats) whose offspring are born at significantly earlier stages of development. Acomys pups are born covered with a soft, grey fur coat, unfurled ears, open eyes and are mobile within 1 or 2 days (Figure 2(a)); indeed, by one week of age they can be completely active and start feeding on solid food in addition to lactation (Figure 2b). At 4 weeks of age, the pups start becoming juveniles and start changing their coat commencing on their backs (Figure 2(c)), where dermal papillae already formed during embryogenesis undergo a second wave of proliferation (the first occurs during embryogenesis) to produce the thicker awl hairs that eventually develop into the genus’s characteristic spines;
34
colour changes gradually to the adult hue (Figure 2(d)). Males and females become sexually mature between 3 and 4 months, but may need to reach between 4 and 6 months to breed successfully, presumably depending on social conditions. A recent report has demonstrated that Acomys females have an 11-day oestral cycle
35
after which, remarkably, they undergo menstruation.
7
After mating, females do not form an evident external vaginal plug, a feature which renders the identification of exact mating time (E0) difficult. Spiny mice have a long gestation period compared to other rodents (39 ± 1 days), after which they give birth to small litters (one to four pups, usually two). Spiny mice are relatively long-lived compared to other murids; our oldest animals are now over 4 years of age, with some publications reporting longevities of up to 6 years in laboratory colonies.36–38
Postnatal development of A. Cahirinus. (a) 2-day-old pup, (b) 1-week-old pup, (c) 4-week-old juvenile and (d) 3-month-old adult.
Acomys have notoriously weak skin with a tensile strength significantly lower than that found in other murids, 10 presumably an anti-predatory adaptation. When handled vigorously, their skin tears easily and large patches of skin can come cleanly off the animal, exposing the underlying muscle fascia. Remarkably, bleeding is minimal, and animals walk off apparently unconcerned and quickly engage in normal behaviors like feeding or grooming. They go on to close their wounds through what looks like epimorphic regeneration, an exceptional trait in a mammal.8–10 Equally interesting is that they do not seem to develop infections in spite of what can be extensive wounds. Lizards are known to show autotomy and can shed their tails, which later regenerate. 39 Acomys show ‘false autotomy’, a tendency to lose the tail sheath due to mechanical stress. A spiny mouse that loses his tail sheath will quickly proceed to eat his own tail and is left with a short stump that does not regenerate. Therefore, Acomys should be handled with care. Transfer of animals to a new cage is best achieved by coaxing them gently into a plastic container tall enough to impede escape; if handled by hand, the handler should wear two pairs of gloves: soft leather gloves covered with disposable latex gloves, as Acomys (particularly adults) can occasionally bite if alarmed. Younger individuals can be picked up by the tail for quick transfers or sexing. Adults should in general not be picked up by the tail, particularly if the animal has been in isolation, as this results in significantly more agitated animals. The animal should be gently grasped with the animal’s back against the handler’s open palm with the fingers gently closing in on the animal’s whole body. Holding their head and front paws immobilized also seems to calm the animals while being handled.
Husbandry and colony maintenance
Housing
Our animals are kept in a standard animal facility. Temperature is maintained at 24–26℃. In our experience, spiny mice thrive well between 20 and 26℃, but may show discomfort (lethargy and stretching out) at temperatures >28℃ and increased huddling (presumably to retain heat) at temperatures <18℃. Pups in particular can suffer hypothermia if exposed to low temperatures for more than 10–15 minutes. Humidity is kept within 30 and 60% and lighting conditions on a 12-hr light/dark cycle. Light intensity is kept below 250 lux.
Our animals are housed in transparent plastic cages of different sizes. Cages of 50 × 30 × 22 cm (or 60 × 45 × 28 cm) are fitted with a metallic grid (1 cm2) lid. The cage should not offer any non-metal surface that the animals can bite into, as they will readily chew holes large enough to exit the enclosure. The number of animals per cage varies between 2–3 or 3–5 adults in the smaller and larger cages, respectively. Alternatively, they can be kept in groups of up to 20 animals in large wire cages (18 × 24 × 16 inches). 11 Spiny mice are highly social and communal breeders, and seem to do well in what could be considered slightly crowded conditions for other rodents. Given their highly social nature, we do not house them individually except in case of disease, injury caused by attack by cage-mates or due to experimental reasons, and if so, strive to minimize the time spent by the animals in isolation. A thin (2-cm deep) layer of substrate is adequate, as spiny mice are notoriously devoid of digging and burrowing behaviors. We use non-scented commercial cat litter, but other substrates like corn bedding and pine shavings can be used. The desert adaptations of spiny mice and their water homeostasis result in relatively odour-free animal rooms (compared to Mus). As a rule, cages are cleaned out and animals moved to fresh substrate every 2 weeks. While increasing the substrate layer to a depth of 3 cm allows substrate changes every 3 weeks without any deleterious effect of any aspect of the animal’s health or reproductive behavior, we prefer a 2-week changing schedule. We provide one, or more, small wooden boxes and/or polyvinyl chloride pipe sections (8 cm diameter) as hiding places or burrows. This minimizes consequences of occasional aggressive behavior that can sometimes be observed in cages and seems to encourage breeding, as females usually have their litters inside the boxes. Dry wooden branches for gnawing and climbing, as well as a small running wheel, are provided for environmental enrichment and seem particularly appreciated by the younger individuals. As animals are usually housed in groups, an individual identification system is required. Ear punches cannot be used due to the fact that Acomys can regenerate extensive full-thickness wounds in the ear pinna (see below). Ear clips are equally unsuitable, as spiny mice ears are relatively delicate and can tear easily. Given their regenerative potential, we have not tested toe clipping as an identification strategy. We currently use microchips (IDVet, Portugal) that can be inserted subcutaneously between the shoulder blades, allowing unambiguous identification of each particular individual.
Diet and nutrition
Spiny mice are highly omnivorous. In the wild they feed on green vegetation, assorted seeds, dry or fleshy fruits, small arthropods, mushrooms and snails. 40 They do not hoard food. It is not known if Acomys consumes meat in their natural habitat,41,42 but cannibalism of pups and dead cage-mates is a relatively common occurrence in our colony. Pups can disappear without a trace, and a cannibalized adult can be reduced to a pile of bones and skin relatively quickly. Acomys is adapted to dry environments 43 and derives a significant part of its water intake from food. 44 This makes Acomys more sensitive to salt and fat-rich diets, which results in a reduction of fertility and an increase in the probability of appearance of type 2 diabetes.3,45,46 Food can be dispensed into a flat ceramic bowl or directly on to the substrate. While they will eat almost everything that is put in front of them, they are not without their preferences. After trying different feeding regimes, we finally settled for a mixture of seeds (commercial poultry seed mixes, rich in wheat, barley, corn, sunflower seeds, etc). Animals are fed approximately 15% of their weight two times a week. As a rule of thumb, no more than 10% of the previous meal should remain uneaten at the next feeding. Once a week, the animals diet is supplemented with a mixture of vegetables and fruit. A number of breeding groups from our colony were fed normal laboratory mouse pellets ad libitum: after three months, breeding dropped off precipitously but recommenced quickly when animals were switched back to their normal seed-based, vegetable/fruit-supplemented diet. Of note, the mouse chow used was standard maintenance type (Ultragene Maintenance Diet, 12% protein content); it is possible that use of a high-protein level mouse chow would allow reproduction to continue, although we have not yet tested this. Protein content can be reinforced with live mealworms, crickets or earthworms once a week. Pups less than 1 month old or animals recovering from sickness or injury benefit from softer food. Animals receive a vitamin supplement (Wevit, Portugal) once a month and mineral water is provided ad libitum. Unrestricted feeding can lead to overweight animals and possibly the development of diabetes. In particular, carbohydrate-rich and especially fat-rich diets should be avoided (see below).
Breeding
For reproduction, we have used breeding groups consisting of one male with one to three females without observing major differences in reproductive output; however, published reports suggest that maximum productivity is obtained with groups consisting of several related females paired to an unrelated male and their offspring, with litter size correlating to female weight and number of immature females in the group. 33 Litters are small, ranging from one to four pups, but most often two pups. Overall sex ratio has been reported as relatively stable at 1:1. 47 If needed, pups can be weaned by 3 weeks of age; alternatively, we occasionally leave offspring in the parental cage for up to 2 months without having observed effects on productivity of the breeding group; indeed, mature females can become receptive shortly after parturition and a new litter will be produced between 45 and 60 days after the previous litter. However, this can be quite variable. Acomys breed year-round, but we have observed variable productivity decreases during winter, despite the fact that bioterium environmental conditions remain unchanged.
Sex can be readily determined at 3–4 weeks of age by inspecting the genitalia, with the distance between the anus and the sexual organs distinctly larger in males than in females (Figure 3). Sexing younger animals is more difficult. Establishing new breeding groups is best done when the animals are still relatively young and sociable (<4 months old), regardless of whether they have started to mature sexually (>1 month old). In this case, animals can be put together in a new cage without any special precautions. Older individuals become more territorial and may not accept new cage-mates readily. Females can be particularly aggressive to new males introduced into their territory, particularly if there is more than one female;
48
new males can be chased, attacked, severely injured or even killed. We have not tested whether this behavior is dependent on the time of year/season of male introduction. When attempting to establish a breeding group with older animals, we put the male in the cage first and allow him 2 or 3 days to establish his territory. We then put a cardboard divider with a window covered with metallic mesh, dividing the cage into two equal areas. A female is introduced into the second compartment, giving animals the opportunity to become habituated to each other’s presence and scent. After 2 days (if the animals have not already chewed through it) the divider can be taken out. Usually some chasing and scuffling occurs (particularly of females chasing males) and can result in scratches, bitten tails and ears, or light skin injuries. After a few hours, the animals calm down. The animals should be monitored to ensure mutual acceptance and separated if aggression results in severe injury. If joining is successful, the divider may be reinstalled and a second female introduced.
Sexing of A. cahirinus.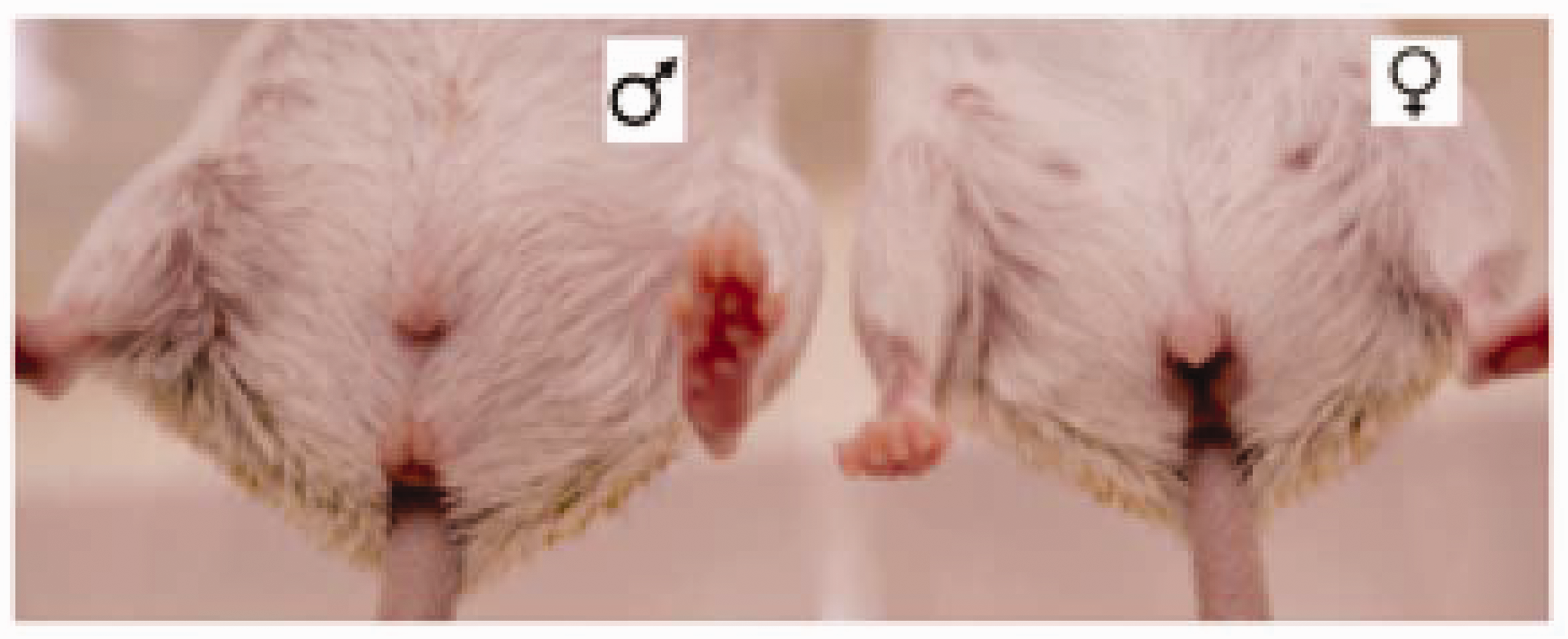
General and reproductive behavior
Once breeding groups are established, the group tends to live harmoniously. Acomys are generally nocturnal; one notable exception is A. russatus, which is nocturnal but adopts a diurnal lifestyle when it shares its habitat with A. Cahirinus. 49 During the day, animals sleep in huddled groups, becoming active for short periods, particularly if they are stimulated by human presence or fed. They are naturally curious and become readily accustomed to their handlers. Of note, cage lids should be well secured, as Acomys are inquisitive, lively, and something of ‘escape artists’. If they do escape, recapturing them can be a challenge, as they are quick runners capable of sizable jumps; recapture is best achieved by using a box-type mouse trap that allows access to food and snaps shut when the animal nibbles at the morsel. Acomys do not burrow, dig or show foot-hopping behavior. Hierarchical relationships are established by chasing and tail nipping, and it is not uncommon for tails to be scratched, bitten and injured. Lower-status animals can be recognized by a greater number of scabs and bite-marks on their tails, and tend to occur in breeding groups under situations of social stress. If a tail becomes severely injured, or if the animal has lost its tail sheath due to poor handling, the usual outcome is that the animal chews off the entire tail, leaving a short stump. Females tend to be dominant over males. A superovulation protocol for spiny mice has recently been reported and could constitute the first step towards the development of transgenic technologies. 50 When the female becomes receptive, the male will chase the female until she is successfully mounted. The coitus itself involves rapid thrusting and lasts only for 1–2 seconds, after which both animals lick their genitalia clean. Somewhat inconveniently for researchers, pregnant females do not form a visible vaginal plug upon pregnancy making determination of E0 problematic. Females become conspicuously heavier around midpregnancy, which lasts approximately 39 days.
Health
Acomys are generally robust and relatively easy to maintain in good health when given good living conditions and a healthy diet. Any new animals arriving at the colony remain quarantined for 2 weeks and are monitored for signs of disease, external parasites, and nasal or ocular secretions. Healthy spiny mice are clear eyed, socially active and inquisitive. Discomfort or pain usually causes the animals to become lethargic and assume a hunched position. Other signs of sickness are semi-closed eyes and puffy eyebags. After the quarantine period is over, animals are integrated into the colony. When cases of sickness do occur, we typically isolate the animal in a separate room and monitor its development. Occasionally, animals may fight and wounds may be found; these are usually small, of little concern and heal quickly without requiring treatment. It will take a very severe skin wound to threaten the life of a spiny mouse; full-thickness wounds of up to 30% of the surface of the back heal without any particular intervention; indeed, after 60 days, little external sign of the wound remains (Figure 4). Histological analysis has shown substantial regeneration of such wounds, including the formation of new hair follicles in the bed wound. Information about disease and parasites of Acomys, either in the wild or in captivity, is limited. Wild-captured Acomys have been reported to carry ticks and fleas,
51
cestodes, nematodes
52
and coccidian protozoa.
53
In addition, a number of blood-borne parasites have been detected, including Babesia, Bartonella, Haemobartonella, Hepatozoon and Trypanosoma.
54
Diagnostic tests specific for Acomys are not commercially available but standard tests available for rodents (PCR assays, foecal floats, pelage and tape tests) can be used to determine the pathogen status of the colony.
11
In addition, we expose sentinels to Acomys soiled bedding and test them twice a year. While we have never detected parasites or infectious agents, we have relatively rarely had animals become sick and quickly die; on one occasion 11 animals died within a 10-day period, suggesting an infectious outbreak from an agent that we were unable to identify. The ‘outbreak’ resolved spontaneously, did not propagate throughout the colony and has not reoccured since.
Remarkable healing and regeneration of skin wounds in adult A. cahirinus. (a) An adult animal caught in a dysfunctional running wheel tore himself free; the wound was trimmed clean but was otherwise not treated. (b) 2 days post-injury, (c) 12 days post-injury, (c) 21 days post-injury and (e) 50 days post-injury.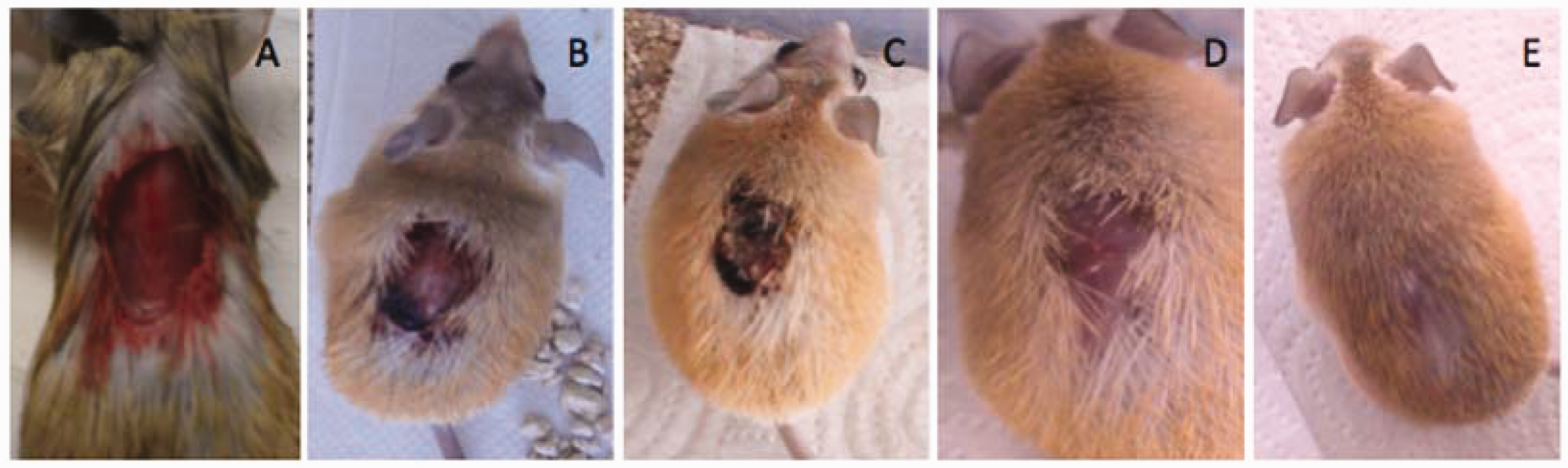
Uses as an experimental model
The first spiny mouse laboratory colonies were established in the 1950s. While Acomys has never been a mainstream model, it has been used to study a range of subjects, including physiology (in particular, adaptations to desert habitats), ecology, evolution, behavior and metabolism. Three major themes stand out: diabetes, pre-natal development and regeneration.
Diabetes research
Obesity and diabetes were reported in an A. cahirinus colony kept at the University of Geneva in the 1960s.3,55 Comparative dietary studies of spiny mice in two different colonies led to the conclusion that the diabetic phenotype was a metabolic response of a desert species to nutritional plenty rather than selection of a genetic variant.45,46 Up to 15% of 1-year-old animals developed nutritionally induced diabetes, with beta-cell hyperplasia, 3 increased pancreatic insulin, low insulin responses, hyperinsulinaemia, hyperglycaemia and glucosuria. 56 After some time, islets of Langerhans broke down, the pancreas atrophied and ketosis occurred, eventually leading to death. 57 During the 1960s and 1970s, the diabetic phenotype of Acomys under fat- or sucrose-enriched diets was characterized. While a fat-rich diet induced obesity and diabetes, a sucrose-rich diet induced hepatomegaly, hyperactive lipogenesis and elevated cholesterol blood levels. Pancreatic hyperplasia and insulinaemia also occurred, but ketosis and pancreatic atrophy were delayed in comparison to a fat-rich diet. Overall, mortality was significantly higher for the fat-rich diet.56,58 From the 1980s onward, diabetes research in Acomys decreased in favour of other more genetically tractable mouse models. From a practical point of view, it is not clear that Acomys individuals within outbred colonies would all become diabetic on high-energy diets, but we proceed under the assumption that they do have a potentially diabetic-prone metabolism.
Pre-natal and perinatal development research
Spiny mice are an interesting model for a range of developmental studies focusing on organogenesis, late pregnancy and perinatal biology. Pups are born in an advanced state of development with open eyes, unfurled ears and completely covered in fur. This precocity results in most of their organ systems being relatively well developed at birth and contrasts markedly with the state of development found in altricial rodent pups. Spiny mouse pups are born capable of basic locomotion and become completely mobile within 1 week. They are also capable of a degree of thermoregulation, although pups under 3 weeks are sensitive to hypothermia if isolated from contact with adults. Liver, lungs, certain brain regions and the kidney all show high levels of maturity at birth.59–64 The spiny mouse model has been used to examine the effect of maternal exposure to glucocorticoids. Exposure to elevated concentrations of dexamethasone levels during midgestation has deleterious effects on placental function and is affected by the sex of the foetus and time elapsed since exposure,65,66 an observation that parallels what is observed in human pregnancies. Another aspect of human pregnancy that has been modeled in Acomys is hypoxia during parturition, which can cause cerebral palsy, cognitive disabilities, impaired memory and retarded development. 67 A study on pups subjected to hypoxia at birth revealed central nervous system (CNS) inflammation and a range of effects in behavioral testing. 67 Follow-up studies suggested that treating pregnant females with melatonin before asphyxia, or supplementing the maternal diet with creatine during pregnancy, improves survival and ameliorates effects on both the structure and function of the CNS, kidneys and diaphragm.4,5,68–72 The model has also been used to study in utero CNS development.59,73,74
Regeneration research
A recent interesting development in spiny mouse research has been the discovery of the ability of this species to regenerate extensively after wounding. Regeneration occurs broadly but unevenly throughout the animal kingdom; however, it is relatively rare in mammals, which tend to respond to injury with fibrotic scarring rather than epimorphic regeneration. 75 In 2012, Seifert and colleagues reported that two species of spiny mice (A. kempi and A. percivali) have weak skin that tears easily in response to mechanical stress. 10 It is not uncommon for animals to suffer extensive, full-thickness skin wounds. Remarkably, the wounds close quickly; histological analysis has shown that bona fide epimorphic regeneration occurs, with animals reconstituting the original tissue architecture, including adipose tissue, dermis, epidermis, sebaceous glands and hair. Moreover, 4-mm circular full-thickness wounds in the ear pinna close fully within 2 months, regenerating hyaline cartilage, adipocytes, dermis, epidermis and hair follicles. 10 This observation has been confirmed in a third member of the species (A. cahirinus) by our own work. 9 In addition, we have observed extensive angiogenesis, nerve fibers and the presence of muscle fibers in the regenerated region. 9 The cellular and molecular mechanisms underlying this regenerative capacity are still poorly understood.
The nature of the cellular and molecular mechanisms underlying the regenerative response in Acomys are currently under investigation. Interestingly, in the ear-punch model, both Mus and Acomys are capable of mounting a proliferative response to wounding; however, this response is abrogated in Mus and only Acomys goes on to establish a proliferating blastema, which eventually leads to regeneration. 76 In the regeneration field, two important themes are the role of the extracellular matrix (ECM) and the immune system in the regulation of the fibrotic vs. regenerative response. During Acomys regeneration, the ECM has significantly lower levels of collagen type 1 deposition than in Mus, 77 with relatively higher levels of collagen III and V. 8 Transcriptional profiling has shown a relatively higher ratio of metalloproteinases to metalloproteinase inhibitors in Acomys as compared to Mus, suggesting higher levels of ECM turnover in Acomys, which could facilitate cell migration. 8 Initial analysis of the role of inflammatory cells in Acomys regeneration suggested that, compared to Mus, Acomys skin wounds have greater numbers of mast cells but lower numbers of mature macrophages. Interestingly, proinflamatory cytokines levels were significantly lower in Acomys, suggesting a blunted inflammatory response in spiny mice. 76 However, a more recent study contradicted this view. While neutrophils and macrophages infiltrated both Mus and Acomys wounds, the relative amounts of cells infiltrating the region had distinct temporal profiles in both systems, with CD11b+ cells being significantly more abundant in Mus at day 3 post-injury. 78 Furthermore, cells showed different distributions, with pro-inflammatory macrophages (CD86+) being excluded from the blastema in Acomys. Importantly, transient local depletion of macrophages by clodronate liposome injection into the ear abrogated blastema formation, which resumed once the depletion was resolved. In addition, the scarring response was characterized by higher myeloperoxidase activity while regeneration showed higher reactive oxygen species activity. 78 The exact role of the different subsets of inflammatory cells in the fibrotic vs. regenerative response remains to be elucidated. An important question is whether the remarkable regenerative capabilities of Acomys are limited to skin and ears or extend to other organs and systems.
Discussion
Spiny mice show a range of biological characteristics worthy of study. While their interest as a model for diabetes type 2 has waned, a number of groups continue to study diverse aspects of their physiology, ecology and behavior. In addition to offering a system in for the study of late organogenesis and perinatal events, they have recently been shown to possess remarkable regenerative abilities, the full extent of which remains to be determined. They are a robust genus that is relatively easy to maintain and breed. Their main limitation is the small litter size and the longer time to sexual maturity (compared to Mus). While a long gestation period offers the advantage of being able to study advanced developmental processes, it also limits their reproductive potential and building up sufficient numbers of animals for research can be challenging. They are also rather uncharacterized in terms of reproductive biology, genetics and genomics; some initial transcriptomic data is published8,77,79 and genome sequencing is underway, but other techniques and reagents, particularly antibodies, remain to be developed. We hope this work can ease the entry of new researchers into the field of Acomys biology.
Footnotes
Acknowledgement
The authors acknowledge funding from the Fundação para a Ciência e a Tecnologia of Portugal and from the Center for Biomedical Research, University of Algarve.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: the Fundação para a Ciência e a Tecnologia (grant PTDC/BIA-ANM/0697/2014 and UID/BIM/04773/2013) and by structural funds from the Center for Biomedical Research, University of Algarve, (UID/BIM/04773/2013 CBMR).